
鲁中南山地黑松人工林土壤和林下植被的动态格局
2018-03-23 09:51:02郭建曜马风云马雪松王文波董玉峰
浙江农林大学学报 2018年2期
郭建曜,王 渌,杨 静,马风云,3,4,马雪松,王文波,董玉峰
(1.山东农业大学 山东省高校森林培育重点实验室,山东 泰安 271018;2.山东省日照市林业局,山东日照276800;3.山东泰山森林生态系统国家定位观测研究站,山东 泰安271018;4.山东农业大学 山东省土壤侵蚀与生态修复重点实验室,山东 泰安 271018;5.山东省林业科学研究院,山东 济南250000)
中国是世界上人工林面积最大的国家。然而,由于中国人工林树种结构单一、管理粗放以及不合理经营采伐等原因,人工林生态系统稳定性较差,林下植被多样性较低等问题随人工林生长逐渐暴露,甚至随人工林生长出现地力衰退、人工林退化等严重情况。因此,本研究旨在揭示人工林生态系统的动态格局,为科学经营、管理人工林提供理论依据,促进人工林健康稳定生长,发挥其应有的经济。生态效益,是人工林经营领域重要的科学问题[1]。林下植被和土壤是森林生态系统的重要组成部分,对森林生态系统结构与功能有重大影响[2-3]。林下植被多样性对于维持森林生态系统物种的多样性、功能的稳定性以及促进养分循环、揭示植被演替特征等方面起着重要的作用[4-5]。林下植物多样性及优势种的变化反映植被演替动态和特征[6]。森林土壤作为森林生长发育的基础,为森林提供其生存所必需的水分、养分等重要物质,土壤水分状况、养分状况、微生物等理化性质及生物性状影响着森林的生长发育[7]。林龄变化影响林下植被和土壤的动态格局,林龄阶段不同,林下植被多样性[8]、土壤物理性状、养分[9]及微生物状况[10]也会有所变化。土壤含水量、容重、毛管孔隙度等是重要的土壤物理性状指标,土壤氮、磷、钾质量分数,土壤酶活性,土壤微生物丰度以及多样性直接或间接影响土壤肥力和养分供应能力[11]。由于人工林树种结构单一,加上长期不合理的高强度经营,导致生物多样性明显低于同龄天然林[12]。有研究表明:幼龄人工林林下植物多样性较造林前提高[13],但随着林龄增加和森林环境的变化,林下植物多样性逐渐降低。林地土壤生态环境也会随林龄增加发生变化[14],如50年生马尾松Pinus massoniana林中土壤纤维素酶、脲酶活性较6年生低[14],巨桉Eucalyptus grandis人工林土壤含水量随林龄增加而增加,土壤容重随林龄增加而逐渐降低,土壤状况改善[15]。黑松Pinus thunbergii是中国暖温带地区干旱瘠薄荒山造林的重要树种。研究黑松人工林生长发育过程中林分群落结构、林下植物多样性、林地土壤理化性质和微生物群落特征的动态格局及其内在关系,对于科学指导林分经营,调控林分群落演替动态,提升森林多种服务功能,实现黑松人工林长期经营具有重要的理论意义。
1 研究区域与研究方法
1.1 样地概况
调查区域位于鲁中南山地的山东省莱芜市雪野镇(36°24′N,117°34′E),海拔为290~320 m,土壤为棕壤,土层厚度为30~40 cm。该区域属温带亚湿润大陆性季风气候,多年平均气温为13.2℃,最低气温-20.0℃,最高气温36.2℃。区域内多为人工植被,林木以耐瘠薄干旱针叶树种为主,自然植被率低,偶见次生落叶、阔叶杂林。多年平均降水量为695.1 mm[16]。
1.2 调查方法
1.2.1 样方设置 采用空间序列代替时间序列的研究方法[17]。在全面踏查基础上,选取处于6,30和50年生,共3个林龄阶段且立地条件相近、林分管理水平一致的黑松林人工纯林。各林龄段分别在选定的林分内设置3块20 m×30 m标准地(经水平校正),实测标准地内乔木的树高、胸径和冠幅。林分立地条件和生长状况如表1。

表1 林分的立地条件与生长状况Table 1 Site conditions and stand growth
1.2.2 林下植被 在设定的标准地内沿S形设置5个5 m×5 m的样方调查其中的下层乔灌木,在每个大样方内设置1 m×1 m的样格调查其中的草本植物。实测林下灌木和草本植物的种类、数量、高度、盖度等。计算草本植物和灌木的重要值、Simpson多样性指数、Shannon-Wiener多样性指数、Pielou均匀度指数。
1.2.3 土壤理化性质调查测定方法 2014年7月调查取样后,土壤物理状况2 d内测量完毕,土壤养分、土壤酶活性6 d内完成测量,土壤微生物样品于取样后2 d内送北京诺禾致源科技股份有限公司检测。①调查方法。在标准地内调查草本和灌木时选取的5个样方中心处作为土壤剖面调查点,清除上层凋落物和腐殖质层,挖掘土壤剖面,用50 cm2的环刀取20 cm深处土壤样本,置于塑封袋中包好,保证土体完整,带回实验室用于分析土壤空隙结构及水分状况;另从各剖面处挖取20 cm深土样,各取约100 g装入塑封袋中,带回实验室,风干后过80目筛,用于土壤养分分析。另将各剖面挖取土壤分别过60目筛,取100 g装入塑封袋,置于冰盒内带回实验室,用于土壤功能酶活性的测定分析。土壤微生物样本的采集采用多点混合取样法,由上述5个剖面挖取20 cm深土并过筛,等量混匀后称取15 g装入离心管,立即置于液氮罐内,用于土壤微生物群落特征分析。②土壤空隙结构。取样前称取干燥环刀质量(G0),称取环刀+鲜土质量(G2),将环刀样品放于盛有2~3 mm水层的瓷盘中8 h,取出拭去沥水后称量(G3),然后置于105℃烘箱内烘干至恒量并称量(G1)。计算土壤容重、毛管孔隙度[19]。③土壤养分。将土样风干压碎过80目筛,采用消煮法全自动凯氏定氮仪(K9860)测定土壤全氮,采用流动注射分析仪(AA3)测定土壤有效氮,采用双光束紫外分光光度计(TU-9500)测定土壤有效磷,采用火焰光度计测定土壤有效钾[18]。④土壤酶活性。脲酶以苯酚-次氯酸钠比色法测定,以培养24 h后1 g土壤中NH3-N的毫克数表示其活性;蔗糖酶以DNS试剂比色法测定,以培养24 h后1 g土壤中生成葡萄糖的毫克数表示其活性;纤维素酶活性以DNS试剂比色法测定,以培养72 h后1 g土壤中生成葡萄糖的毫克数表示其活性[19]。均使用双光束紫外分光光度计(TU-9500)比色。⑤土壤微生物群落特征。测序公司根据所扩增的16S区域特点,基于Illumina MiSeq测序平台,利用双末端测序(Paired-End)的方法,构建小片段文库进行双末端测序。从公司提供的测序分析报告中得到土壤微生物Chao1指数及Shannon指数。
1.3 数据处理
采用Excel处理实验测定的土壤理化性质数据并制作图表,使用SAS 9.0对数据进行统计分析,使用Duncan新复极差法(P<0.05)做多重比较,使用Pearson分析法进行偏相关分析。
2 结果与分析
2.1 不同林龄阶段黑松人工林的林下植被特征
在所有样地中,黑松人工林林下植被共出现32种植物,分属17科28属,其中灌木植物6科6属7种,草本植物11科22属25种。黑松林下灌木种类偏少,草本种类相对较多(表2)。从灌木类树种来看,6年生阶段黑松林仅有2种,30年生阶段有5种,50年生有4种。其中6年生阶段林下优势灌木树种为截叶铁扫帚Lespedeza cuneata,30年生林下优势灌木树种为荆条Vitex negundo,50年生林下优势灌木树种为荆条和截叶铁扫帚。
不同林龄阶段黑松林下草本植物群落组成各不相同,每个林龄阶段均有少数几种优势植物。其中6年生阶段黑松林下草本植物优势种类为苦荬菜Ixeris denticulata,隐子草属1种Cleistogenes sp.,结缕草Zoysia japonica,狗尾草Setaria viridis。苦荬菜对土壤条件要求不高;狗尾草喜温暖湿润,但也具有一定抗旱、抗寒能力;隐子草和结缕草都具有良好的抗干旱能力。30年生草本优势种为苋草Alternanthera payonychiodes,卷柏Selaginella tamariscina和苦苣菜Sonchus oleraceus。

表2 不同林龄阶段黑松人工林林下植被概况Table 2 Undergrowth vegetation general situation of Pinus thunbergii plantation in different age stages
从图1可以看出:6年生和30年生黑松林的Simpson指数为0.86~0.88,差异不大;但是50年生松林只有0.76,比前2个年龄阶段的黑松林明显减小。
6年生和30年生 Shannon-Wiener指数相差不大,分别为2.293和2.314。50年生的Shannon-Wiener指数只有1.676,比最高的30年生降低27.6%。Shannon-Wiener为变化度指数,是一种能较好地反映个体密度、生境差异、群落类型、演替阶段的指数,它是物种丰富度和均匀度的函数。物种数量越多,分布越均匀,Shannon-Wiener值也越大。从不同林龄Shannon-Wiener指数可以看出,6年生与30年生黑松林的物种丰富度以及物种分布的均匀程度相差不大,50年生黑松林物种丰富度以及物种分布的均匀程度低于前2个林龄阶段。
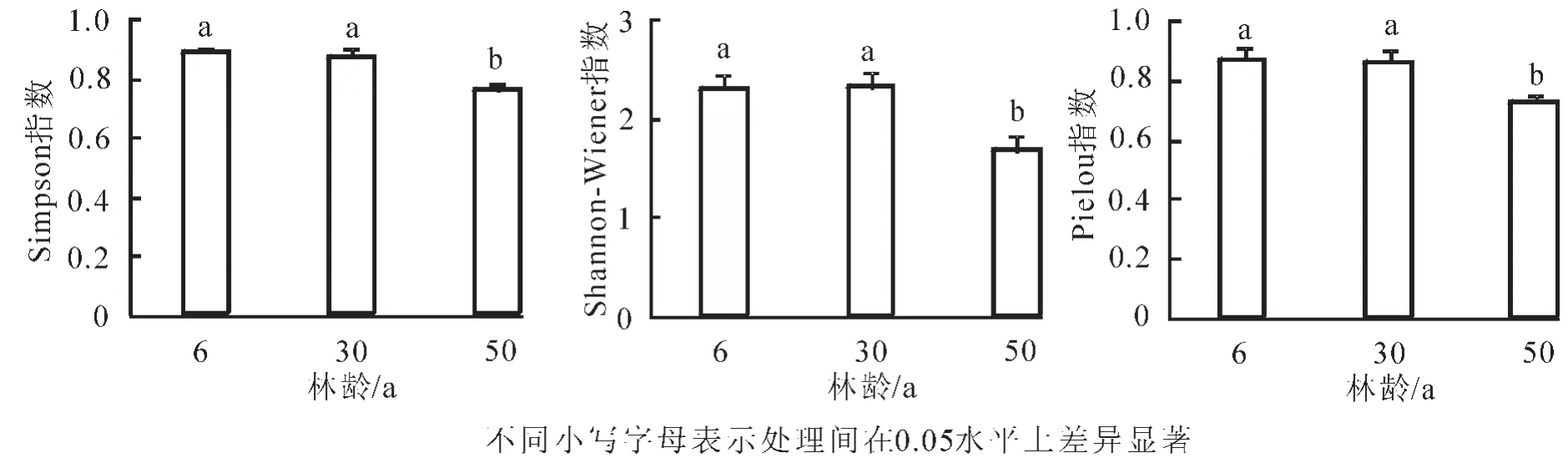
图1 不同林龄阶段黑松人工林多样性指数Figure 1 Diversity index of P.thunbergii plantation in different age stages
群落的Pielou均匀度是指群落中各个种的重要值的均匀程度,它所表征的是群落实测多样性与最高多样性之间的比率。6年生与30年生黑松林的Pielou指数相差不大,分别为0.869和0.862,而到50年生降低较大,只有0.728,比6年生降低了16.2%。
2.2 不同林龄阶段黑松人工林的土壤理化性质
2.2.1 土壤容重及毛管孔隙度 从图2看出:土壤容重随林龄的增大而降低,50年生比6年生黑松林降低了15.6%,差异显著(P<0.05)。土壤毛管孔隙度随林龄的增大递增,50年生比6年生增加了32.3%,差异显著(P<0.05)。
2.2.2 土壤养分状况 黑松林分的有效磷质量分数随着林龄的升高呈下降趋势(图3),在30年生时下降尤为明显,与6年生相比下降了36.6%,差异达显著水平(P<0.05);但50年生开始有所回升,与30年生相比提高了16.1%,差异显著(P<0.05)。土壤有效钾质量分数呈先升高后降低的趋势,在30年生时达最高值,且不同林龄之间差异显著(P<0.05)。不同林龄黑松土壤有效氮质量分数在数值上差异不大,30年生最高,为0.107 g·kg-1,比最低的50年生高8.2%(0.008 g·kg-1),且30年生与另2个林龄间有显著差异。但土壤全氮质量分数与有效氮质量分数趋势一致,为30年生黑松林>6年生黑松林>50年生黑松林,50年生黑松林比6年生黑松林、30年生黑松林分别降低了27.2%和28.5%,达显著差异水平(P<0.05)。
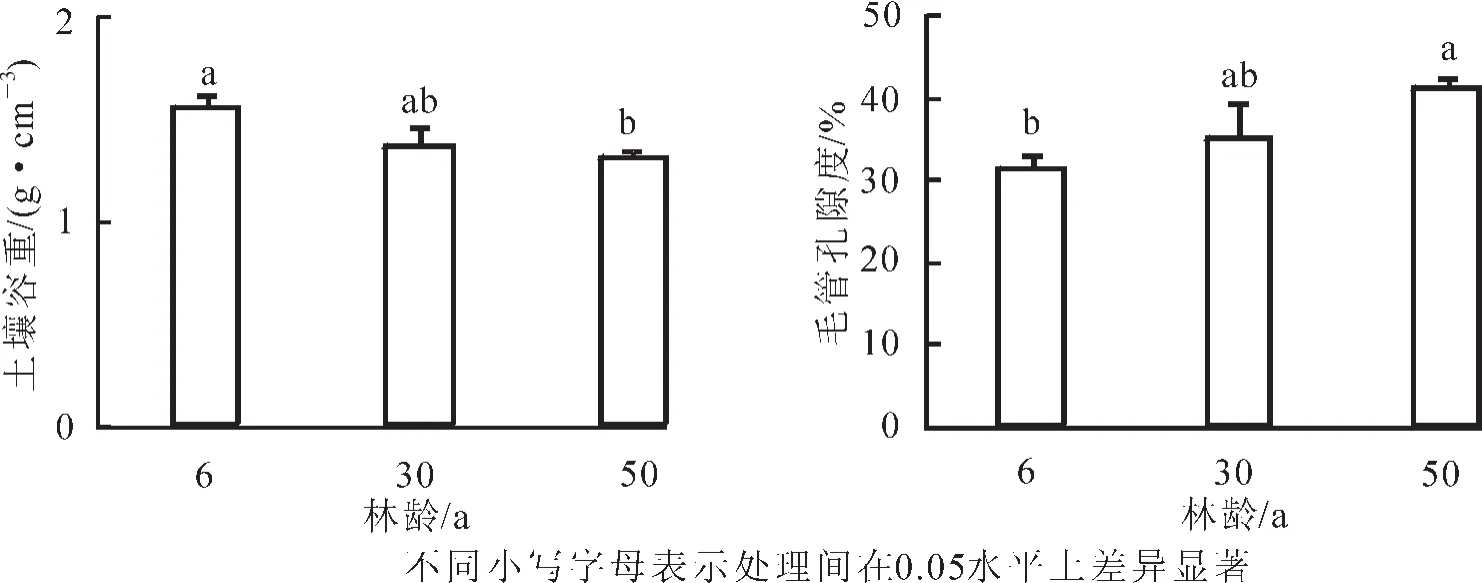
图2 不同林龄黑松土壤容重与毛管孔隙度Figure 2 Soil bulk density and capillary porosity of Pinus thunbergii plantation in different age stages

图3 不同林龄黑松土壤养分状况Figure 3 Soil nutrient status of P.thunbergii plantation in different age stages
2.2.3 土壤功能酶活性 不同林龄黑松林中,蔗糖酶活性先升高后降低,黑松林在30年生时达最高(图4)。50年生蔗糖酶活性最低,比6年生和30年生分别降低了32.2%和44.3%,且不同林龄之间差异显著(P<0.05)。土壤纤维素酶也是呈先升高后降低的趋势,黑松林在30年生时最高,50年生最低,比6年生和30年生时分别降低了33.6%和59.7%,且不同林龄之间差异显著(P<0.05)。脲酶活性为6年生黑松林>30年生黑松林>50年生黑松林。脲酶活性最低的50年生黑松林比6年生和30年生黑松林分别降低了43.0%和40.2%,差异显著(P<0.05)。由表3可以看出:黑松人工林土壤酶与土壤养分之间存在较多相关性。蔗糖酶活性与全氮呈极显著相关,与有效氮呈显著相关;脲酶活性与全氮呈极显著相关;纤维素酶活性与有效氮、有效钾呈极显著相关,与全氮呈显著相关。
2.3 不同林龄黑松人工林的土壤细菌群落特征
2.3.1 微生物多样性 α多样性分析是指对群落中物种的丰富度和个体在物种中的分布均匀程度指标进行分析。本研究使用Chao1指数和Shannon指数对土壤中细菌菌群多样性进行分析。Chao1指数为丰富度指数,其值越高则表明细菌群落物种的丰富度越高。不同林龄黑松人工林土壤种细菌Chao1指数为6年生>50年生>30年生,呈现出先降后增的趋势(图5)。最低的30年生黑松林与最高的6年生黑松林相比降低了24.3%,差异显著(P<0.05);50年生黑松林比30年生黑松林升高了10.1%,无显著差异。Shannon指数反映细菌群落的多样性程度,其值越高表示细菌群落结构的多样性越高。由图5可以看出:土壤细菌Shannon指数为6年生黑松林>50年生黑松林>30年生黑松林,同样为先降后增的趋势,且不同林龄间差异显著(P<0.05)。最低的30年生黑松林比最高的6年生黑松林降低了5.6%,50年生比30年生升高了2.8%。
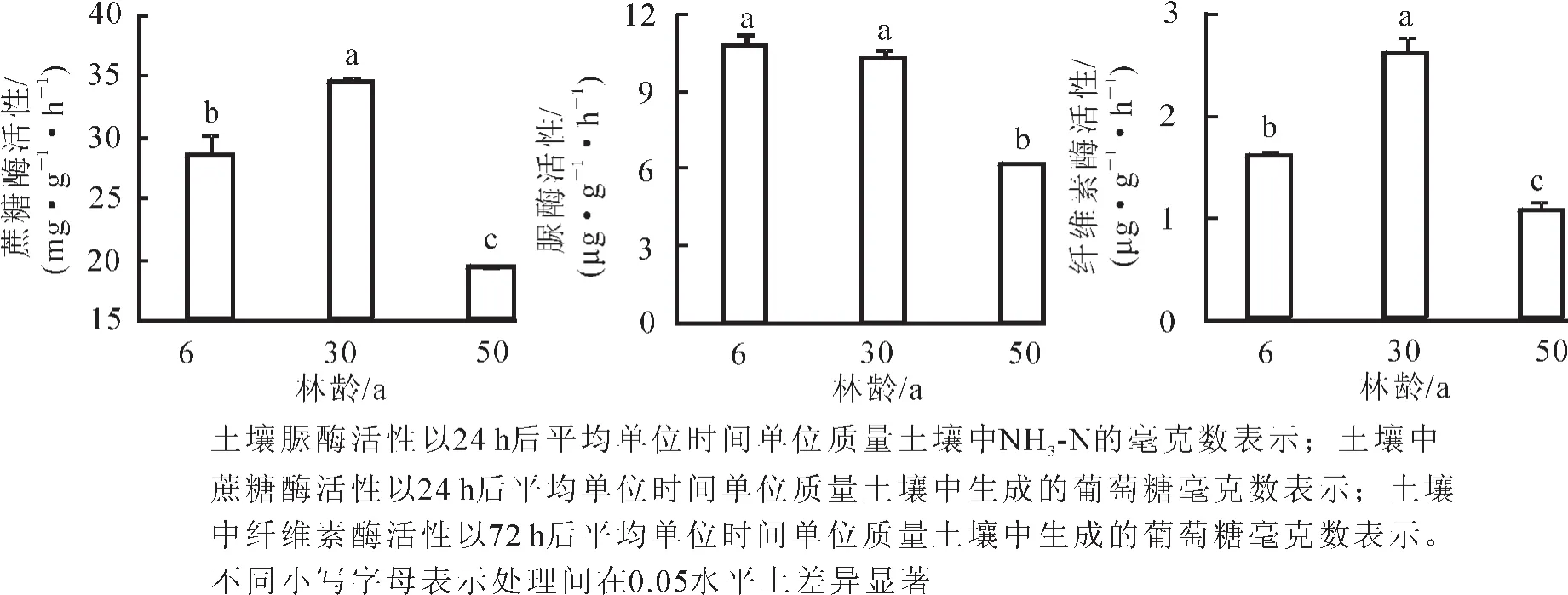
图4 不同林龄黑松土壤酶状况Figure 4 Soil enzyme status of Pinus thunbergii plantation in different age stages

表3 土壤酶与土壤养分相关性Table 3 Correlation between soil enzyme and soil nutrient
2.3.2 优势菌群 由图6可以看出:在门水平上,黑松林地中的变形菌门Proteobacteria,放线菌门Actinobacteria和酸杆菌门Acidobacteria占绝对优势,各林龄这3类细菌数量之和均占到细菌总量的72%以上。随着黑松林龄的增长,变形菌门丰度先升后降,在30年生时显著高于6年生和50年生(P<0.05),占这种细菌总量的36.5%;放线菌门的丰度随林龄的增长递减,由6年生时的32.2%降低到50年生时的24.0%,下降了25.5%,差异显著(P<0.05);酸杆菌门丰度则是随着林龄的增长而递增,由6年生时的14.8%上升到50年生时的22.4%,升高了51.4%,差异显著(P<0.05)。

图6 门水平土壤细菌相对丰度前10位Figure 6 Relative abundance of top 10 soil bacteria in Phylum level
3 讨论
3.1 黑松人工林林下植被多样性及物种组成随林龄增加而变化
黑松人工林林下光照及土壤养分状况等环境因子随林龄而变化,进而影响林下植物的群落组成和多样性。黑松幼龄人工林(6年生)林分郁闭度较低,林内光照条件好,一些喜光、抗逆性强的先锋植物首先入侵并定居,如荆条、隐子草、结缕草等,其重要值较高,占据优势。随着林龄增加,森林凋落物增加,土壤主要矿质元素如有效氮、有效钾质量分数升高,土壤容重降低,30年生时一些对土壤环境要求相对较高的植物,如喜肥水的苋草和苦苣菜等,开始占据优势。此时虽然林下植被种间竞争逐渐加剧,物种组成发生变化,但整体多样性仍得以维持,30年生林下植被多样性指数与6年生时相比略有变化,但均无显著差异。进入50年生时,林分郁闭度进一步增加,林下光照条件变差,林地土壤养分周转速率降低,土壤有效钾、有效氮、全氮下降,林下植被对光照和养分等生活因子的种间竞争愈发激烈,抗逆、耐荫的植物,如隐子草和截叶铁扫帚等逐渐占据优势,一些喜光趋肥植物,如苋草和苦苣菜等则被逐渐淘汰,林下植被多样性指数显著降低,植被群落逐渐发生演替。
3.2 土壤主要矿质养分及功能酶活性随林龄的增加而变化
土壤矿质元素是影响土壤功能酶活性的重要因素,同时土壤功能酶又是土壤养分循环中重要的生物催化剂[20]。6年生至30年生黑松人工林林分郁闭度逐渐增加,随林分及林下植被生长,凋落物增加。此时期林分自身养分利用率较低,养分周转较快,有利于维持林地矿质养分质量分数[21],至30年生时土壤有效氮、有效钾质量分数升高,全氮质量分数无显著变化。凋落物量以及有效氮、有效钾质量分数升高又直接或间接影响到土壤纤维素酶、蔗糖酶活性[22],引起30年生黑松林地土壤纤维素酶、蔗糖酶活性的显著升高。由于脲酶参与氮素循环,其活性与全氮质量分数显著相关,而30年生黑松林地全氮质量分数与6年生时相比并无显著差异,因此其脲酶活性同样无显著差异。同时30年生黑松林地脲酶活性得以维持,有利于氮素的转化,这也是30年生时有效氮质量分数升高的原因之一。30~50年生时随着林分继续生长,养分周转速率降低[21],伴随着林分及林下植被生长消耗,至50年生时土壤有效钾、有效氮、全氮质量分数均显著降低。受此影响[22],土壤纤维素酶、蔗糖酶、脲酶活性也显著降低。
本研究中土壤有效磷质量分数随林龄增加的变化趋势与其他矿质养分变化趋势相反,推测是由于有效磷被植物吸收消耗、淋溶损耗,而磷素生物有效性过程却比较缓慢[23]导致的。6~30年生黑松林地有效磷随植被生长的吸收利用以及淋溶等逐渐消耗,而通过凋落物分解等过程有效归还缓慢,导致30年生时土壤有效磷显著降低。30~50年生时随着之前积累的凋落物等逐步分解,土壤有效磷得到的补充增加,同时土壤容重逐渐降低,有利于减少磷素淋溶损耗[24],所以至50年生时有效磷质量分数开始回升,但仍显著低于6年生黑松林地。
3.3 黑松人工林土壤微生物群落特征随林龄增加而变化
土壤微生物群落丰度和多样性在一定程度上反映土壤生态系统的演变特征,并随着林龄变化而变化。土壤中细菌多样性6年生黑松林>50年生黑松林>30年生黑松林,表现为随林龄的增加呈先降低后升高的变化趋势。随林龄增加,由于林下植被对光照、水分、养分的竞争逐渐加剧,至30年生时虽然林下植被多样性指数与6年生林相比无显著差异,但林下植被物种组成发生较大变化,喜水肥的植物出现并占据优势。同时,林分逐渐郁闭导致光照条件变差,土壤热量输入减小,土壤有机物降解速率下降[25]。这些环境的扰动都会对微生物多样性产生负面影响[25-26],所以30年生黑松林地土壤微生物多样性显著降低。30~50年生时,虽然林下植被多样性指数显著降低,但经过长时间的竞争演替其物种组成已趋于稳定,如林下草本中隐子草已占据绝对优势,灌木则是以荆条为主,因此由种间竞争、物种组成变化所引起的环境扰动减少。同时,林下灌草植物多样性下降,盖度降低,土壤热量输入增加,土壤有机质降解速率加快。这些条件均有利于微生物多样性增加,因此至50年生时林地土壤微生物多样性与30年生时相比显著升高。这一规律与MENYAILO[27]的研究结果一致。
土壤功能微生物类群决定土壤养分的循环。有研究发现,变形菌门Proteobacteria的一个亚门β-Proteobacteria,包含几乎所有种类氨氧化细菌[28],具有固氮作用。黑松人工林土壤中变形菌门的丰度随林龄的增加先升高后降低,与土壤中全氮以及速效氮质量分数的变化趋势一致。放线菌门作用于土壤肥力的形成和提高[29-30]。黑松人工林土壤中放线菌门的丰度随林龄的增长而递减,而酸杆菌门的丰度随林龄的增加而升高。这2个门下的细菌种属是否参与土壤养分循环尚有待进一步研究。
[1] 姜俊,陆元昌,谢阳生,等.基于发展类型理念的多功能人工林经营模式研究[J].世界林业研究,2015,28(3): 1-5.JIANG Jun,LU Yuanchang,XIE Yangsheng,et al.A study of multi-functional forest management modes based on forest development type [J].World For Res,2015,28(3):1-5.
[2] 尤业明,徐佳玉,蔡道雄,等.广西凭祥不同年龄红椎林林下植物物种多样性及其环境解释[J].生态学报,2016, 36(1): 164-172.YOU Yeming,XU Jiayu,CAI Daoxiong,et al.Environmental factors affecting plant species diversity of understory plant communities in a Castanopsis hystrix plantation chronosequence in Pingxiang,Guangxi,China [J].Acta Ecol Sin,2016,36(1):164-172.
[3] 尤龙辉,叶功富,陈增鸿,等.公路建设对木麻黄生长及林下植被物种多样性的影响[J].浙江农林大学学报,2013, 30(1): 38-47.YOU Longhui,YE Gongfu,CHEN Zenghong,et al.Species diversity in a Casuarina equisetifolia understory along a highway [J].J Zhejiang A&F Univ,2013,30(1):38-47.
[4] 康冰,刘世荣,蔡道雄,等.马尾松人工林林分密度对林下植被及土壤性质的影响[J].应用生态学报,2009, 20(10): 2323-2331.KANG Bing,LIU Shirong,CAI Daoxiong,et al.Effects of Pinus massoniana plantation stand density on understory vegetation and soil properties [J].Chin J Appl Ecol,2009,20(10):2323-2331.
[5] 冯耀宗.物种多样性与人工生态系统稳定性探讨[J].应用生态学报,2003,14(6):853-857.FENG Yaozong.Species diversity and managed ecosystem stability [J].Chin J Appl Ecol,2003,14(6):853-857.
[6] 杨宁,邹冬生,杨满元,等.衡阳紫色土丘陵坡地不同恢复阶段植被特征与土壤性质的关系[J].应用生态学报, 2013, 24(1): 90-96.YANG Ning,ZOU Dongsheng,YANG Manyuan,et al.Relationships between vegetation characteristics and soil properties at different restoration stages on slope land with purple soils in Hengyang of Hunan Province,South-central China [J].Chin J Appl Ecol,2013,24(1):90-96.
[7] 周莉,代力民,谷会岩,等.长白山阔叶红松林采伐迹地土壤养分含量动态研究[J].应用生态学报,2004,15(10): 1771-1775.ZHOU Li,DAI Limin,GU Huiyan,et al.Dynamics of soil nutrient contents in cutting forestlands of broad-leaved Korean pine forest on Changbai Mountains [J].Chin J Appl Ecol,2004,15(10):1771-1775.
[8] 崔宁洁,陈小红,刘洋,等.不同林龄马尾松人工林林下灌木和草本多样性[J].生态学报,2014,34(15):4313-4323.CUI Ningjie,CHEN Xiaohong,LIU Yang,et al.Shrub and herb diversity at different ages of Pinus massoniana plantation [J].Acta Ecol Sin,2014,34(15):4313-4323.
[9] 孙鹏跃,徐福利,王渭玲,等.华北落叶松人工林地土壤养分与土壤酶的季节变化及关系[J].浙江农林大学学报, 2016, 33(6): 944-952.SUN Pengyue,XU Fuli,WANG Weiling,et al.Seasonal dynamics of soil nutrients and soil enzyme activities in Larix principis-rupprechtii plantations [J].J Zhejiang A&F Univ,2016,33(6):944-952.
[10] 牛小云,孙晓梅,陈东升,等.辽东山区不同林龄日本落叶松人工林土壤微生物、养分及酶活性[J].应用生态学报, 2015, 26(9): 2663-2672.NIU Xiaoyun,SUN Xiaomei,CHEN Dongsheng,et al.Soil microorganisms,nutrients and enzyme activity of Larix kaempferi plantation under different ages in mountainous region of eastern Liaoning Province,China [J].Chin J Appl Ecol,2015,26(9):2663-2672.
[11] 黄玉梅,杨万勤,张健,等.川西亚高山针叶林土壤微生物及酶对林下植物去除的响应[J].生态学报,2014, 34(15): 4183-4192.HUANG Yumei,YANG Wanqin,ZHANG Jian,et al.Response of soil microorganism and soil enzyme activity to understory plant removal in the subalpine coniferous plantation of western Sichuan [J].Acta Ecol Sin,2014,34(15):4183-4192.
[12] RAMOVS B V,ROBERTS M R.Understory vegetation and environment responses to tillage,forest harvesting and conifer plantation development[J].Ecol Appl,2003,13(6):1682-1700.
[13] 李伟,张翠萍,魏润鹏.广东中西部桉树人工林植物多样性与林龄和土壤因子的关系[J].生态学报,2014, 34(17): 4958-4965.LI Wei,ZHANG Cuiping,WEI Runpeng.Relationship of under-story vegetation diversity with stand age and soil factor of eucalypt plantations in central-western Guangdong [J].Acta Ecol Sin,2014,34(17):4958-4965.
[14] 葛晓改,肖文发,曾立雄,等.三峡库区不同林龄马尾松土壤养分与酶活性的关系[J].应用生态学报,2012, 23(02): 445-451.GE Xiaogai,XIAO Wenfa,ZENG Lixiong,et al.Relationships between soil nutrient contents and soil enzyme activities in Pinus massoniana stands with different ages in Three Gorges Reservoir Area [J].Chin J Appl Ecol,2012,23(02):445-451.
[15] ZHANG Danju,ZHANG Jian,YANG Wanqin,et al.The effect of afforestation with Eucalyptus grandis on soil physicochemical and microbiological properties [J].Soil Res,2012,50(2):167-176.
[16] 王承军,王琪珍,卜庆雷.莱芜近52年气候变化特征分析[J].江西农业学报,2011,23(2):167-170.WANG Chengjun,WANG Qizhen,BU Qinglei.Analysis on climatic change of Laiwu City in recent 52 years [J].Acta Agric Jiangxi,2011,23(2):167-170.
[17] 杨宁,邹冬生,杨满元,等.衡阳紫色土丘陵坡地植被恢复阶段土壤特性的演变[J].生态学报,2014,34(10): 2693-2701.YANG Ning,ZOU Dongsheng,YANG Manyuan,et al.Changes of soil properties in re-vegetation stages on slopingland with purple soils in hengyang of Hunan Province,South-central China [J].Acta Ecol Sin,2014,34(10):2693-2701.
[18] 章家恩.生态学常用实验研究方法与技术[M].北京:化学工业出版社,2007.
[19] 关松荫.土壤酶及其研究方法[M].北京:农业出版社,1986.
[20] 姜小凤,王淑英,丁宁平,等.施肥方式对旱地土壤酶活性和养分含量的影响[J].核农学报,2010,24(1):136-141.JIANG Xiaofeng,WANG Shuying,DING Ningping,et al.Effect of different fertilization on soil enzyme activity and nutrients [J].J Nucl Agric Sci,2010,24(1):136-141.
[21] 葛晓改,周本智,肖文发.马尾松人工林凋落物产量、养分含量及养分归还量特性[J].长江流域资源与环境, 2014, 23(7): 954-961.GE Xiaogai,ZHOU Benzhi,XIAO Wenfa.Study on litterfall,nutrient content and nutrient return of Pinus massoniana plantation [J].Resour Environ Yangtze Basin,2014,23(7):954-961.
[22] 万忠梅,吴景贵.土壤酶活性影响因子研究进展[J].西北农林科技大学学报(自然科学版),2005,33(6):87-92.WAN Zhongmei,WU Jinggui.Study progress on factors affecting soil enzyme activity [J].J Northwest Sci-Tech Univ Agric For Nat Sci Ed,2005,33(6):87-92.
[23] RICHER D D,ALLEN H L,LI Jianwei,et al.Bioavailability of slowly cycling soil phosphorus:major restructuring of soil P fraction over four decades in an aggrading forest[J].Oecologia,2006,150(2): 259-271.
[24] 吕家珑.农田土壤磷素淋溶及其预测[J].生态学报,2003, 23(12): 2689-2701.LÜ Jialong.Phosphorus leaching from agricultural soils and its prediction [J].Acta Ecol Sin,2003,23(12):2689-2701.
[25] STEENWERTH K L,JACKSON L E,CALDERÓN F J,et al.Soil microbial community composition and land use history in cultivated and grassland ecosystems of coastal California [J].Soil Biol Biochem,2002,35(3):489-500.
[26] 张逸飞,刘娟娟,孟磊,等.农业利用对海南省天然次生林土壤微生物的影响[J].生态学报,2015,35(21): 6983-6992.ZHANG Yifei,LIU Juanjuan,MENG Lei,et al.Agricultural use of natural secondary forests affects soil microorganisms in Hainan Province,China [J].Acta Ecol Sin,2015,35(21):6983-6992.
[27] MENYAILO O V.The influence of tree species on the biomass of denitrifying bacteria in gray forest soils [J].Eurasian Soil Sci,2007,40(3):302-307
[28] STEPHEN J R,MCCAIG A E,SMITH Z,et al.Molecular diversity of soil and marine 16S rRNA gene sequences related to b-subgroup ammonia-oxidizing bacteria [J].Appl Environ Microbiol,1996,62(11): 4147-4154.
[29] 杨斌,段泉宏,陈占全,等.青海沙珠玉人工植被系统土壤放线菌生态分布及拮抗性[J].应用生态学报,18(8): 1694-1701.YANG Bin,DUAN Qaunhong,CHEN Zhanquan,et al.Ecological distribution and antimicrobial effects of soil actinomycetes in artificial vegetation systems in Shazhuyu of Qinghai,China [J].Chin J Appl Ecol,2008,18(8):1694-1701.
[30] QUAISER A,OCHSENREITER T,LANZ C,et al.Acidobacteria form a coherent but highly diverse group within the bacterial domain [J].Mol Microbiol,2003,50(2):563-575.
猜你喜欢
美食(2022年12期)2022-12-09 02:08:48
安徽农业科学(2021年9期)2021-06-22 15:02:49
防护林科技(2020年10期)2020-12-22 11:46:42
美食(2019年7期)2019-09-10 07:22:44
科学大众·小诺贝尔(2018年12期)2018-02-22 12:05:38
防护林科技(2016年9期)2016-09-27 01:34:21
防护林科技(2016年5期)2016-09-05 01:19:43
防护林科技(2016年1期)2016-02-27 06:09:54
现代企业文化(2015年1期)2015-02-28 18:58:20
亚热带资源与环境学报(2015年1期)2015-01-22 07:04:58
