
区域性农田景观格局对棉蚜种群数量的生态学效应
2018-03-22 11:33:04张晴晴卢增斌李丽莉关秀敏欧阳芳张永生门兴元
生态学报 2018年4期
张晴晴,卢增斌,李丽莉,关秀敏,欧阳芳,张永生,门兴元,*
1 山东省农业科学院植物保护研究所,济南 250100 2 山东省农业厅植物保护总站,济南 250100 3 中国科学院动物研究所,农业虫害鼠害综合治理研究国家重点实验室,北京 100101 4 湖南农业大学植物保护学院,长沙 410128
农田景观是多种作物生境及其周边环境组成的斑块镶嵌体,在景观尺度上表现为物种生存的各类缀化栖地的空间网络结构[1]。一般来说,农田景观基本处于一种相对稳定的格局,但是近年来,随着人类对农田土地利用强度的增加,农田景观格局产生了剧烈的改变,而这种变化势必会影响到景观中害虫的发生、转移、扩散及自然天敌的生物控害功能,从而影响害虫种群数量和对作物的危害。因此,研究农田景观格局变化对害虫种群的生态学效应,不仅可以揭示人类活动对生物多样性结构与功能的影响,阐明农田景观中生物多样性整合和维持机理,而且实践上可为制定害虫区域性治理策略提供基础信息[2]。
农田景观格局的主要特征可以采用“质、量、形、度”来概括,其中“质”表示农田景观中不同的景观组成,“量”表示不同类型斑块的大小和面积比例等,“形”表示不同斑块类型的形状和排列方式,“度”则反映农田景观格局变化的时间和空间尺度[3]。目前,国内外的研究更多集中在农田尺度条件下非作物生境组成对农作物害虫的影响,例如,非作物栖息地维持了昆虫的多样性,多种木本和草地生境可为寄主性和捕食性天敌昆虫提供备选食物[4- 9]。同时,非作物栖息地也为一些害虫提供安全的越冬场所,从而有利于它们越冬[10]。而对于更大空间尺度条件下害虫对景观格局的响应研究相对较少,通过大尺度下系统研究多景观因子(质、量、形、度)对害虫种群发生的生态学效应,更能明确景观格局因子对农田害虫种群的驱动机制。已经有研究表明,不同类型的害虫对于农田景观格局的响应不同,例如草地与林地生境对麦蚜种群增长率促进最大,塑料大棚抑制麦蚜种群增长;但草地、林地与塑料大棚对蚜茧蜂的种群增长均有促进作用[11],麦田生态系统中麦长管蚜和禾谷缢管蚜种群数量在复杂农田景观中超过简单农田景观,而麦二叉蚜却恰恰相反[12-14];然而在同一作物生态系统中相似的物种对于农田景观格局的响应也可能有很大差异,例如麦田麦岩螨的发生量与森林斑块结构呈显著正相关,而麦圆叶爪螨种群数量与景观内水体斑块结构密切相关,两种害螨对不同景观格局的响应是其生物学特性差异的体现[15]。那么,农田生态系统中害虫的不同发生时期对农田景观格局响应是否一致?目前尚无相关报道。
棉花是世界性的经济作物,也是关系民生的重要经济作物。我国是世界上最大的棉花生产国,同时也是最大的棉花进口国。棉蚜(AphisgossypiiGlover)是危害棉花的主要害虫,广泛分布于60°N至40°S地区[16]。棉蚜在黄河流域棉区危害十分严重,根据其不同发生时期,通常将棉花苗期的棉蚜称为苗蚜,成株期的棉蚜称为伏蚜,其中苗蚜在棉叶背面以及嫩尖上刺吸汁液,能造成叶片卷曲植株矮缩,破坏棉株的正常代谢,苗蚜分泌的蜜露还会影响棉花植株的光合作用[17];伏蚜密布于整个棉株,在叶背及嫩茎刺吸为害,可造成叶片发黄、变黑和蕾铃脱落,甚至导致棉花落叶而严重减产[18]。
山东省位于黄河流域,一直是我国棉花种植大省。近年来,随着产业结构调整、耕作制度变革及城镇化建设导致包括棉田在内的农田景观格局变迁, 使山东省区域性农田景观格局特征呈现多样化。因此,本研究以山东省棉花种植区为研究区域,通过卫星遥感影像和土地覆盖分类数据的分析,获得山东省14个样本县区的农田景观因子指数,田间系统调查获取2008—2010年各个样本县区的苗蚜和伏蚜的种群数量,分析了3类景观格局因子, 如景观组成类型、构成比例和形状结构与棉花苗蚜和伏蚜种群数量的相关性,从而明确农田景观格局对苗蚜和伏蚜的生态学效应,解析棉蚜不同发生时期对农田景观格局响应的差异,为棉蚜的区域性治理提供理论和技术支持。
1 材料和方法
1.1 研究区域
山东省属温带季风气候,年平均温度12—16℃,降雨量600—1200mm,非常适合棉花生长,2008—2010年山东省棉花种植面积均在73万hm2以上。选取山东省14个植棉县区,分别是:博兴、东昌府区、定陶、陵县、阳谷、德城区、莘县、茌平、临邑、商河、平度、邹平、沾化和滨城区。
1.2 遥感数据获取
1.2.1 遥感影像和土地覆盖分类
研究中使用的遥感数据源为2010年Landsat TM/ETM的中分辨率卫星影像(10—30m)。使用ENVI 5.0遥感图像处理软件对图像进行等预处理,结合野外调查数据,建立解译图谱库,获得2010年山东省土地覆被分类的栅格数据,将研究区域内的土地覆盖类型归为8类:森林、草地、湿地、水体、耕地、园地、居住工业道路交通、其他用地,方法参考欧阳芳等[3]。由于山东省内几乎没有草地和湿地,临近棉田的园地较少,因此文中只涉及森林、水体、耕地和居住工业道路交通4类。
1.2.2 景观格局分析
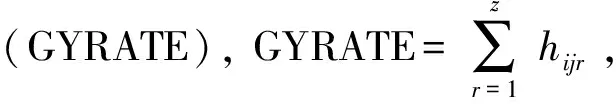
1.3 棉蚜种群数量
根据中华人民共和国国家标准-棉蚜测报调查规范(GB/T15799—1995)系统调查2008年至2010年期间山东省14个县区棉蚜发生的种群数量,其中苗蚜的调查时间是5月中下旬,此时正是苗蚜的危害高峰期,每个县区中尽量选择临近不同土地覆盖类型的棉田10块,每块田5点取样,定苗前每点查40株,定苗后每点查20株,统计百株蚜量;伏蚜是在6月25日前后,在上述10块棉田中,每块田采取5点取样法,每点调查10株,每株查主茎上的上(第一展开叶)、中、下(第一果枝主茎叶)三片叶子统计蚜量,计算百株(3叶)蚜量。
1.4 数据分析
运用统计软件SPSS 16.0分别分析苗蚜和棉蚜的种群数量的频次分布,并进行正态拟合曲线。利用R 3.3.1计算苗蚜、伏蚜种群数量与景观因子指数之间的相关系数r并进行显著性分析。对所有分析,设置显著性水平为P=0.05。
2 结果分析
2.1 棉蚜种群数量的数据特征与分布
棉蚜种群在山东省不同棉花种植区呈现明显的区域性分布特征。棉花苗蚜的种群数量分布在300—7000头/百株的区间范围内,14个取样县区的平均发生量为1611.42头/百株,最大值为6968.33头/百株,最小值为328.10头/百株;伏蚜的种群数量在0—1500的区间范围内,平均发生量为187.06头/百株,最大值为1416.67头/百株,最小值为0(图1)。
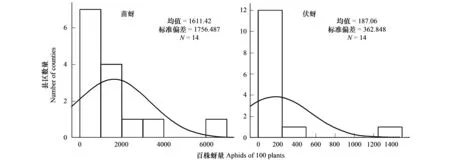
图1 以山东省为典型代表的苗蚜和伏蚜的种群数量特征Fig.1 The characteristic of populations of cotton seedling aphids and summer aphids in Shandong Province
14个县区中棉田苗蚜数量与伏蚜数量没有显著相关性(r=0.4511,df=13,P=0.1092),对各县区的苗蚜和伏蚜发生数量分别进行排序,各县区苗蚜与伏蚜的数量排序差别很大。商河县苗蚜的发生量最大,而其伏蚜的发生量为265.33/百株,排第2;平度苗蚜的发生量最少,但其伏蚜发生量为173.33头/百株,排第4;陵县的伏蚜发生量最大,而其苗蚜发生量仅为907.00头/百株,排名第9;邹平的伏蚜发生为0,而苗蚜的发生量为1105.83头/百株,排名第7(图2)。
2.2 景观格局对棉蚜种群数量的影响
2.2.1 景观组成对棉蚜种群数量的影响
棉田苗蚜和伏蚜的种群数量与景观组成因子的相关性不一致。其中,苗蚜的种群数量与景观总面积(TA)呈显著正相关(r=0.9582,P<0.0010),而伏蚜的种群数量与斑块丰富密度(PRD)呈显著正相关(r=0.6911,P=0.0062)(表1)。
2.2.2 景观构成对棉蚜种群的影响
景观构成指数与棉蚜苗蚜和伏蚜的关系也不一致。对苗蚜的种群数量影响最大的景观构成因子是Simpson多样性指标(SIDI),呈显著负相关(r=-0.6094,P=0.0461),而伏蚜与景观构成中的因子相关性均不显著(表2)。
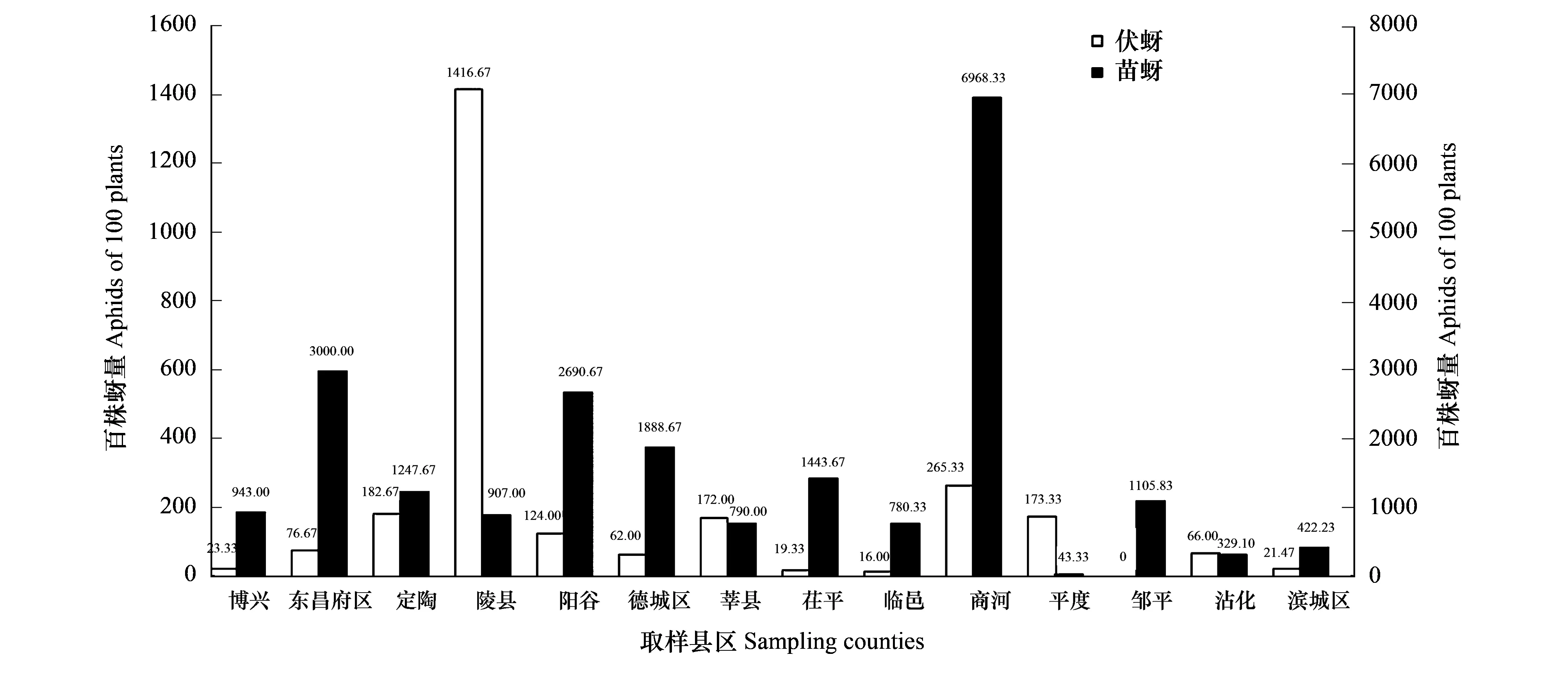
图2 山东省14县区棉田苗蚜和伏蚜种群数量Fig.2 Population density of cotton seedling aphids and summer aphids in 14 counties of Shandong Province
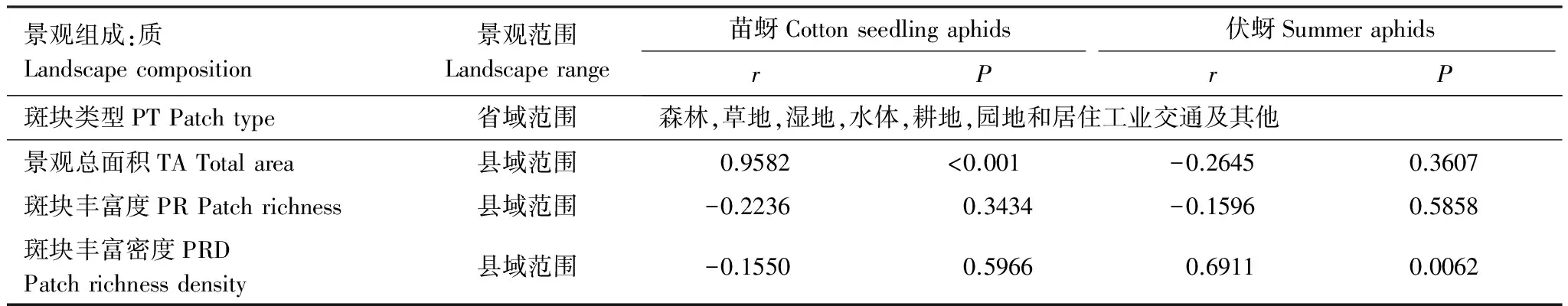
景观组成:质Landscapecomposition景观范围Landscaperange苗蚜Cottonseedlingaphids伏蚜SummeraphidsrPrP斑块类型PTPatchtype省域范围森林,草地,湿地,水体,耕地,园地和居住工业交通及其他景观总面积TATotalarea县域范围0.9582<0.001-0.26450.3607斑块丰富度PRPatchrichness县域范围-0.22360.3434-0.15960.5858斑块丰富密度PRDPatchrichnessdensity县域范围-0.15500.59660.69110.0062

表2 景观构成与棉蚜的相关性
2.2.3 景观结构对棉蚜种群的影响
景观结构指数与棉蚜苗蚜和伏蚜的关系也不一致。苗蚜的种群数量与耕地的分形指数(r=0.6103,P=0.0323)、县域范围的蔓延度(r=0.5435,P=0.0446)和县域范围的回旋半径(r=0.6327,P=0.0152)呈显著正相关;伏蚜的种群数量与居住工业交通的蔓延度(r=0.5677,P=0.0317)呈显著正相关。(表 3)
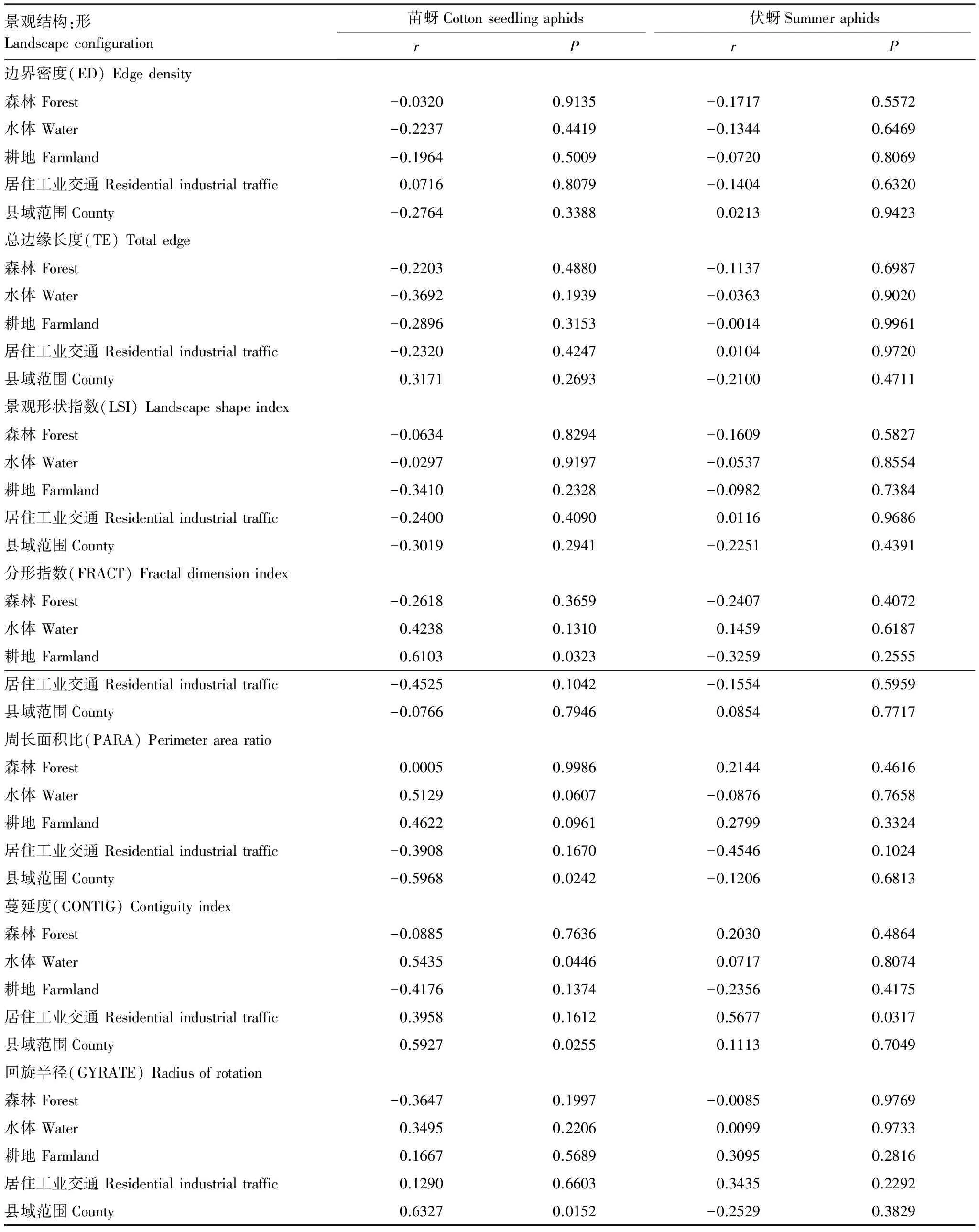
表3 景观结构与棉蚜的相关性
3 讨论
本研究发现,棉蚜的种群数量与景观组成、景观构成和景观结构有密切的关系,且棉花苗期的苗蚜和成株期的伏蚜对景观因子的响应并不一致。已经有研究表明不同种类的害虫对于农田景观格局响应不同[13-15],研究结果表明,同一害虫的不同发生时期,可能由于食物资源数量、质量和温湿度等环境条件等不同,对农田景观的响应产生了很大差异。
3.1 景观组成类型和构成比对棉蚜种群的影响
苗蚜的发生与景观组成(质)中的景观总面积有显著相关性,可能与景观中苗蚜的取食、越冬等资源数量有关,其机理还需要进一步研究。Simpson多样性指标(SIDI)与苗蚜的发生呈显著的负相关,Simpson多样性指标(SIDI)越高,苗蚜的种群数量越小,SIDI指数是反映农田景观多样性的指标,农田景观斑块的多样性越高越不利于苗蚜的发生,这可能是由于多样化的农田景观可以为自然天敌提供更加丰富的替代食物和栖境,从而有利于自然天敌的种群维持和控害功能的发挥[10],自然天敌在不同斑块间迁移能够有效的控制苗蚜[8]。前人研究也表明山东省农田景观中瓢虫等自然天敌对苗蚜有极强的自然控制作用,自然控害作用高达90%以上[20]。
伏蚜与斑块丰富度密度(PRD)有显著的正相关性,PRD的值越大,伏蚜的种群数量越多。PRD为景观中斑块类型数与景观总面积的比值,表示单位景观面积中斑块类型数,也是农田景观多样性的重要指标。有研究显示,斑块丰富度密度(PRD)越大,越利于麦田中麦蚜种群数量的增加,可能是由于小麦成熟收割后麦蚜迁飞转移到临近的非作物生境内,而栖息在非作物斑块内的麦蚜在下一季小麦生长期间,再回到小麦田,从而增加了麦田蚜虫的数量[3]。山东省农田景观中自然天敌对伏蚜的自然控制作用较小,自然控害作用在20%以下[20],因此多样性的农田景观虽然有利于自然天敌控害功能,但是其控制伏蚜种群的作用有限,而多样化的斑块类型,可能通过为伏蚜提供多样化的寄主,从而增加景观中伏蚜的种群数量,已有研究表明,农田系统斑块的多样性可以增加棉盲蝽、棉铃虫等的种群数量[21-22]。总之,苗蚜与伏蚜对农田景观格局的响应不同,这种不一致可能是通过食物资源与自然天敌控害对棉蚜不同发生时期的影响不同而产生的。
3.2 景观结构对棉蚜种群的影响
影响棉蚜发生的另一类重要景观格局因子是景观结构(形)。蔓延度(CONTIG)是指景观中不同斑块类型的非随机性或聚集程度,一般来说,高蔓延度值说明景观中的某种优势斑块类型形成了良好的连接性,相对的破碎化程度较低[19]。本文结果显示县域范围内斑块的蔓延度越高,越有利于苗蚜的发生,已有研究表明农田景观区域内田块面积越大越集中,而其他类型的景观要素类型越少以及其面积越小,越不利于天敌的持续稳定维持种群数量[3],因此不稳定的天敌种群可能是导致苗蚜大量发生的重要因素之一。另外,处于不同营养级的物种,对于景观破碎化具有不同的敏感性[23],苗蚜对景观破碎化的敏感性可能大于天敌,因此在连续性较好的斑块中属于r-对策者的棉蚜大爆发时,天敌不能在短时间内快速繁殖对苗蚜进行控制,具体的机制需要进一步验证。
伏蚜的种群数量与居住工业交通的蔓延度指数(RIT)呈显著正相关,RIT越高意味着人类的活动范围越集中,相对于农田生态系统的破碎化程度小,已有研究表明,线性景观要素(如高速公路等) 对两侧动物种群的遗传结构和基因流表现了明显的隔离作用[24-25]。因此农田生态系统的破碎化程度越小,越有利于伏蚜的繁殖。
由此可见,棉蚜的两个发生时期(棉蚜和苗蚜)的发生与景观格局有密不可分的关系,且苗蚜和伏蚜对景观的响应不完全一致。苗蚜和伏蚜对景观蔓延度(形)响应是一致的,景观的破碎化程度越小,伏蚜和苗蚜发生越重。苗蚜和伏蚜对景观多样性(质)的响应不一致,景观多样性高的农田景观不利于苗蚜的发生,对伏蚜没有显著影响;丰富度有助于伏蚜的发生,却对苗蚜没有显著影响。这一结果显示了农业害虫的不同发生时期对农田景观格局响应的复杂性。在棉蚜发生较重的区域,根据其苗蚜和伏蚜的发生情况,可通过减少越冬场所、减少斑块的蔓延度等,结合其他生物防治及化学防治的方法形成控制棉蚜的种群数量的区域性综合治理策略。相同物种在不同的生育期对景观结构的不同响应机制有待深入研究。
[1] Turner M G. Spatial and temporal analysis of landscape patterns. Landscape Ecology, 1990, 4(1): 21- 30.
[2] 欧阳芳, 戈峰. 农田景观格局变化对昆虫的生态学效应. 应用昆虫学报, 2011, 48(5): 1177- 1183.
[3] 欧阳芳, 门兴元, 关秀敏, 肖云丽, 戈峰. 区域性农田景观格局对麦蚜及其天敌种群的生态学效应. 中国科学: 生命科学, 2016, 46(1): 139- 150.
[4] Denys C, Tscharntke T. Plant-insect communities and predator-prey ratios in field margin strips, adjacent crop fields, and fallows. Oecologia, 2002, 130(2): 315- 324.
[5] Rieux R, Simon S, Defrance H. Role of hedgerows and ground cover management on arthropod populations in pear orchards. Agriculture, Ecosystems & Environment, 1999, 73(2): 119- 127.
[6] Nicholls C I, Parrella M, Altieri M A. The effects of a vegetational corridor on the abundance and dispersal of insect biodiversity within a northern California organic vineyard. Landscape Ecology, 2001, 16(2): 133- 146.
[7] Hickman J M, Wratten S D. Use of Phelia tanacetifolia strips to enhance biological control of aphids by overfly larvae in cereal fields. Journal of Economic Entomology, 1996, 89(4): 832- 840.
[8] Men X, Ge F, Yardim E N, Parajulee M N. Evaluation of winter wheat as a potential relay crop for enhancing biological control of cotton aphids. BioControl, 2004, 49(6): 701- 714.
[9] Ouyang F, Men X Y, Yang B, Su J W, Zhang Y S, Zhao Z H, Ge F. Maize benefits the predatory beetle,Propyleajaponica(Thunberg), to provide potential to enhance biological control for aphids in cotton. PLoS One, 2012, 7(9): e44379.
[10] Bianchi F J J A, Booij C J H, Tscharntke T. Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proceedings of the Royal Society B-Biological Sciences, 2006, 273(1595): 1715- 1727.
[11] 赵紫华, 王颖, 贺达汉, 关晓庆, 辛明. 麦蚜和寄生蜂对农业景观格局的响应及其关键景观因子分析. 生态学报, 2012, 32(2): 472- 482.
[12] Letourneau D K, Goldstein B. Pest damage and arthropod community structure in organic vs. conventional tomato production in California. Journal of Applied Ecology, 2001, 38(3): 557- 570.
[13] Jonsen I D, Fahrig L. Response of generalist and specialist insect herbivores to landscape spatial structure. Landscape Ecology, 1997, 12(3): 185- 197.
[14] 赵紫华, 石云, 贺达汉, 杭佳, 赵映书, 王颖. 不同农业景观结构对麦蚜种群动态的影响. 生态学报, 2012, 30(23): 6380- 6388.
[15] 卢增斌, 欧阳芳, 张永生, 关秀敏, 门兴元. 华北平原地区景观格局对麦田害螨种群数量的影响. 生态学报, 2016, 36(14): 4447- 4455.
[16] 高桂珍, 吕昭智, 夏德萍, 孙平. 高温胁迫及其持续时间对棉蚜死亡和繁殖的影响. 生态学报, 2012, 32(23): 7568- 7575.
[17] 张永孝, 赵之刚, 曹赤阳. 棉蚜为害损失与防治指标的研究. 植物保护学报, 1982, 9(4): 229- 236.
[18] Castle S J, Perring T M, Farrar C A, Kishaba A N. Field and laboratory transmission of watermelon mosaic virus 2 and zucchini yellow mosaic virus by various aphid species. Phytopathology, 1992, 82(2): 235- 240.
[19] 郑新奇, 付梅臣. 景观格局空间分析技术及其应用. 北京: 科学出版社, 2010.
[20] 于汉龙, 门兴元, 叶保华, 于毅, 张安盛, 李丽莉, 周仙红, 张思聪, 庄乾营. 自然天敌对苗蚜和伏蚜控制作用的定量分析. 应用昆虫学报, 2014, 51(1): 99- 106.
[21] Allen K C, Luttrell R G. Spatial and temporal distribution of heliothines and tarnished plant bugs across the landscape of an Arkansas farm. Crop Protection, 2009, 28(9): 722- 727.
[22] 吕昭智, 潘卫林, 张鑫, 李贤超, 张娟. 新疆北部棉区作物景观多样性对棉铃虫种群的影响. 生态学报, 2012, 32(24): 7925- 7931.
[23] Anton C, Zeisset I, Musche M, Durka W, Boomsma J J, Settele J. Population structure of a large blue butterfly and its specialist parasitoid in a fragmented landscape. Molecular Ecology, 2007, 16(18): 3828- 3838.
[24] Keller I, Nentwig W, Largiadèr C R. Recent habitat fragmentation due to roads can lead to significant genetic differentiation in an abundant flightless ground beetle. Molecular Ecology, 2004, 13(10): 2983- 2994.
[25] Vos C C, Chardon J P. Effects of habitat fragmentation and road density on the distribution pattern of the moor frogRanaarvalis. Journal of Applied Ecology, 1998, 35(1): 44- 56.
猜你喜欢
植物保护(2023年1期)2023-02-03 10:22:08
农药学学报(2022年6期)2022-12-27 12:00:28
基层中医药(2021年4期)2021-07-22 07:15:32
新疆农业科学(2020年11期)2020-10-27 09:05:32
——管氏肿腿蜂的应用技术
江苏林业科技(2020年3期)2020-08-31 01:00:08
今日农业(2019年13期)2019-08-12 07:59:02
人民调解(2019年5期)2019-03-17 06:55:16
现代园艺(2017年21期)2018-01-03 06:42:15
现代农业(2016年4期)2016-02-28 18:42:12
小说月刊(2015年12期)2015-04-23 08:51:10
