
Hap1型棉蚜在5种春季杂草上的生长发育情况
2023-02-03 10:22:08杨心怡苏宏华周福才景田兴杨益众
植物保护 2023年1期
鲍 安, 杨心怡, 苏宏华, 周福才, 景田兴, 杨益众, 张 帅
(扬州大学植物保护学院, 扬州 225009)
棉蚜Aphisgossypii属于半翅目Hemiptera蚜科Aphididae,是一种广泛分布的多食性害虫[1]。棉蚜的寄主植物可达116科912种之多,主要包括锦葵科、葫芦科、茄科、芸香科和菊科的农林作物及观赏花卉[2]。研究证明棉蚜的越冬寄主为木槿Hibiscussyriacus、花椒Zanthoxylumbungeanum和石榴Punicagranatum等木本植物,而夏寄主几乎覆盖了所有种类的蔬菜。生产上,由于对瓜类为害较为严重,又叫做瓜蚜。通过寄主转接试验,采自棉花Gossypiumhirsutum和黄瓜Cucumissativus上的棉蚜不能在对方寄主上建立种群[3-4]。随着研究的深入,分子鉴定技术逐步应用于棉蚜的研究,在棉蚜线粒体基因组序列的基础上,Zhang等[5-6]分析了华北棉蚜的单倍型,将华北地区棉蚜分为57个单倍型,其中Hap1型棉蚜为棉花上的主要类型,占华北棉蚜总数的54%(共计采集到1 046个棉蚜)。生物测定结果表明Hap1型棉蚜属于典型的棉花型棉蚜[7]。
棉蚜的生活史可分为全周期型和不全周期型。研究表明,大多数采自棉花上的棉蚜经诱导都可产生性蚜,而Hap1型棉蚜为棉花上的主要类型,推测Hap1型棉蚜在我国主要营全周期型生活史[8]。即其在秋季转移到冬寄主上产卵越冬,早春时孵化成干雌,经数代孤雌生殖之后,迁移到夏寄主上繁殖为害。经过观察,越冬寄主上棉蚜在3月底和4月初就已孵化并大量繁殖。而其主要为害作物——棉花,最早在4月底播种。在棉花出苗之前,已有大量棉蚜迁离越冬寄主,所以推测在棉蚜迁移到棉田之前存在中间寄主供其繁殖。之前研究也发现,棉蚜除了在木本植物上越冬,也可在刺儿菜Cirsiumarvense、夏至草Lagopsissupina等杂草上产卵越冬、繁殖,到棉花出苗时迁移到棉田为害棉苗[9-10]。
早春一般指一年中的2月到3月。在这期间,温度和湿度普遍较低。以扬州地区为例,温度一般在5~15℃,而湿度仅有50%。在此环境下大部分作物尚未播种或是刚出苗,但适宜荠菜Capsellabursa-pastoris、飞廉Carduusnutans等杂草生长。因此大部分地块边缘和部分地块中央呈现出杂草丛生的状态,在早春棉田中更是如此。此外,棉田杂草还作为中间寄主传播病虫害,加重了棉田病虫害的发生,给棉田病虫害防治带来诸多困难。比如,早春小地老虎Agrotisipsilon会先在野豌豆Viciasepium上产卵孵化,生长至 2~3 龄期时再转移到小麦和棉花上为害[10]。棉田杂草种类多,不仅各季节群落差异巨大,其在不同地区也有不同的群落组成。以我国3大棉区为例,长江流域棉区雨水充沛,气候温润,根据王荣龙等[11]的调查结果,赣北棉区棉田共有35种杂草分属14科。黄河流域棉区因地区气温低于长江流域,降水量也较少,故杂草多为喜凉耐旱型。根据房锋等[12]的调查结果,山东主要棉区的棉田杂草有24科81种。而西北内陆棉区降水量稀少,昼夜温差大,因此棉田杂草多为耐盐、耐旱型。南疆棉田的杂草有16科,42种,其常见杂草有马唐Digitariasanguinalis、芦苇Phragmitesaustralis、田旋花Convolvulusarvensis、马齿苋Portulacaoleracea、苘麻Abutilontheophrasti、刺儿菜、龙葵Solanumnigrum等。其中常见阔叶杂草占比达棉田杂草的80%左右。北疆地区棉田主要杂草有17科,44种,龙葵、灰绿藜Oxybasisglauca和田旋花为优势杂草[13]。在田间害虫防治过程中,消灭虫源是重要的技术手段。推测春季杂草可以作为棉蚜种群扩张的寄主,成为棉田棉蚜的重要虫源地,而对于春季杂草在棉蚜种群繁殖过程的作用以及棉蚜可以取食的杂草种类还未见报道。本文选择扬州早春田间棉蚜发生较重的荠菜和阿拉伯婆婆纳Veronicapersica,及其他3种优势杂草,通过生命表的方式研究了棉花型棉蚜中具有代表性的Hap1型棉蚜对它们的利用情况,试图揭示早春杂草在棉蚜为害棉花过程中的中间寄主地位,为日后棉田棉蚜发生的预测、预报以及防治提供依据。
1 材料与方法
1.1 试验材料
1.1.1供试昆虫
供试棉蚜2020年采自扬州市扬州大学文汇路校区试验田的棉花植株,将其带回实验室,单头饲养建立种群。按照Zhang等的[5]方法使用单倍型特异序列进行生物型鉴定。确定其为Hap1型棉蚜后,在人工智能光照培养箱[(26±1)℃,L∥D=14 h∥10 h,RH=70%~80%,下同]内的健康棉花植株上饲养数代,用于后续试验。
1.1.2供试植物
试验所用棉花品种为‘SGK321’,由中国农业科学院植物保护研究所提供;杂草选用阿拉伯婆婆纳,荠菜,猪殃殃Galiumspurium,飞廉,飞蓬Erigeroncanadensis。所有植株均种植于人工气候室内,种植条件与供试昆虫培养条件一致。
1.2 试验方法
1.2.1棉蚜生命表的建立
试验在人工智能光照培养箱内进行。将植物叶片的叶柄处用吸水脱脂棉保湿,正面贴于含有1.8%(质量分数)固体琼脂的培养皿(直径9 cm)中,适时换叶以保持叶片新鲜。将棉花上的无翅Hap1型棉蚜成虫接到目标寄主植物叶片上,寄主植物包括5种早春杂草,以棉花叶片作为对照,每种寄主上接20头棉蚜成虫。当24 h内棉蚜产仔超过35头时,移去成蚜,保留35头仔蚜,以后每天调查并记录存活虫量。待其长到3龄左右时将其单独移至对应的寄主叶片上,每天观察产仔量,并及时扫除所产仔蚜,直至起始蚜虫全部死亡。选Hap1型棉蚜在目标寄主植物上的子一代成虫20头,使用Motic SMZ-171图像测量系统测量成蚜的体长、体宽。
1.2.2数据分析
试验记录的原始数据按照生命表软件处理数据所要求格式进行统计,计算棉蚜各阶段发育历期、雌成虫寿命、总发育历期和产卵历期等。根据CHI等[14]的计算方法,基于年龄阶段、两性生命表理论,采用计算机程序TWOSEX-MSChart对所有个体的原始生命表数据进行分析。利用bootstrap技术估计种群参数的方差和标准误差,采用TWOSEX-MSChart程序中Paired bootstrap test对数据进行差异显著性检验,并采用Graphpad Prism 8.0以及Origin软件进行作图。
2 结果与分析
2.1 Hap1型棉蚜取食5种杂草及棉花的若虫发育历期
棉蚜若虫共有4个龄期,本研究共测定了其在5种杂草及棉花叶片上的若虫发育历期。结果(表1)显示,供试棉蚜在不同寄主上的若虫发育历期存在明显差异。取食荠菜叶片的棉蚜成虫前期最短,为4.16 d。取食阿拉伯婆婆纳、飞廉和飞蓬叶片的棉蚜成虫前期相差不到0.1 d。取食猪殃殃叶片的棉蚜成虫前期大于5 d,显著长于取食棉花、阿拉伯婆婆纳、飞廉、荠菜和飞蓬叶片的棉蚜成虫前期。比较各龄若虫的发育历期发现,取食猪殃殃叶片成虫前期较长主要是由于1龄若虫发育历期显著长于取食其他寄主。取食各种寄主的棉蚜4龄若虫的发育历期无显著差异(表1)。
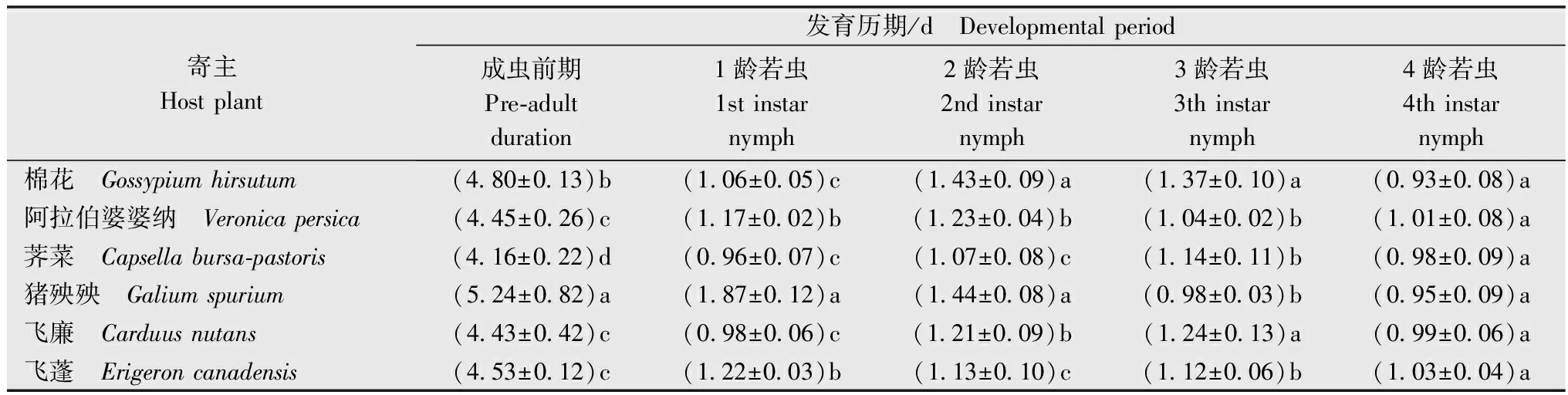
表1 6种寄主植物对Hap1型棉蚜生长发育的影响1)
2.2 Hap1型棉蚜取食5种杂草及棉花的繁殖力
相较于取食棉花叶片的Hap1型棉蚜,取食杂草的棉蚜成虫生殖前期缩短,产仔数减少。在5种杂草叶片上饲养的蚜虫的总生殖前期显著短于在棉花叶片上饲养的蚜虫。取食5种杂草的Hap1型棉蚜间总生殖前期无显著差异,其中取食猪殃殃叶片的Hap1型棉蚜总生殖前期最短,为5.31 d。取食阿拉伯婆婆纳和飞蓬的Hap1型棉蚜成虫生殖前期显著短于取食其他寄主的成虫生殖前期。取食荠菜、猪殃殃和飞廉叶片的Hap1型棉蚜的成虫生殖前期无明显差异,但显著短于取食棉花叶片的成虫生殖前期(表2)。Hap1型棉蚜在棉花叶片上平均每雌产仔数为64.30头,显著高于其他5种寄主;产仔最少的是飞蓬上饲养的Hap1型棉蚜,平均每雌产仔数为29.22头,产仔数显著低于其他5种寄主。
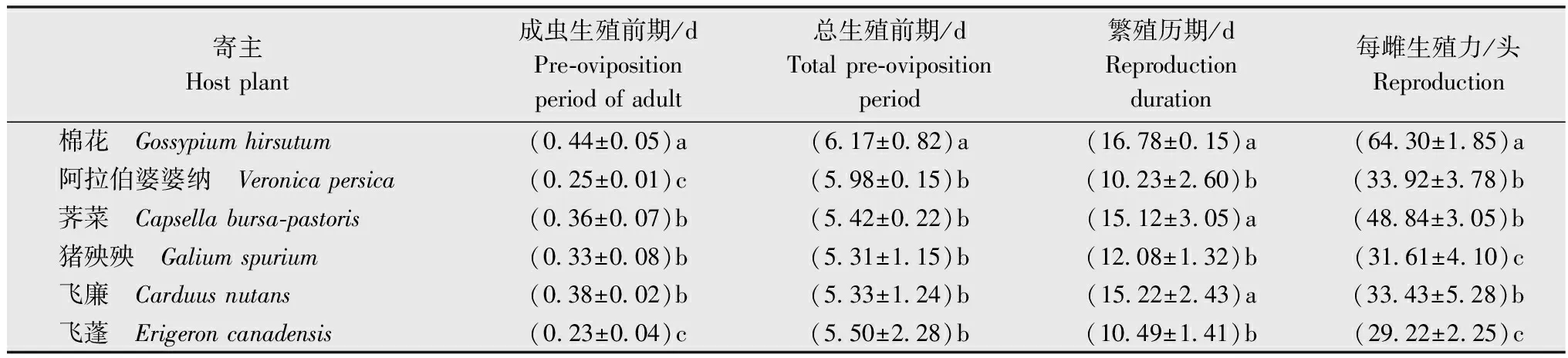
表2 寄主植物对Hap1型棉蚜繁殖的影响1)
2.3 Hap1型棉蚜取食5种杂草及棉花的存活率
Hap1型棉蚜在5种杂草上普遍可以存活25 d左右,但明显低于在棉花上的存活时间,在棉花叶片上最长可以存活44 d。取食杂草叶片的Hap1型棉蚜在第10 天左右开始出现死亡,取食5种杂草的棉蚜死亡率变化趋势相似。而取食棉花叶片的Hap1型棉蚜在第30 天时才开始出现死亡。
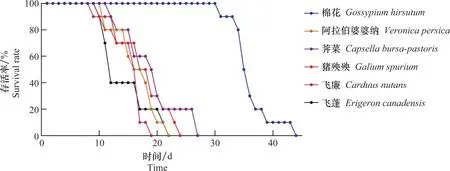
图1 Hap1型棉蚜在6种寄主植物上的存活率
2.4 Hap1型棉蚜在5种杂草及棉花上的生命表参数
Hap1型棉蚜取食5种杂草及棉花的生命表参数结果显示(表3),Hap1棉蚜最适寄主为棉花,取食棉花时寿命(41.50 d)和净增殖率(64.30)都最高,显著高于取食其他寄主;但其在杂草上也能建立种群。取食荠菜和飞廉的净增殖率均超过40,显著高于取食其他3种杂草的Hap1型棉蚜;取食飞蓬叶片的Hap1型棉蚜净增殖率最低,为29.43。在世代平均周期方面,按寄主依次为:荠菜(11.93 d)>猪殃殃(11.59 d)>棉花(11.38 d)>飞蓬(11.17 d)>飞廉(10.22 d)>阿拉伯婆婆纳(8.88 d)。此外,棉蚜在6种寄主植物上生活的内禀增长率也各不相同,其中取食棉花和阿拉伯婆婆纳的内禀增长率分别为0.37和0.39,显著高于取食其他寄主。

表3 Hap1型棉蚜在6种寄主植物上的生命表参数1)
2.5 Hap1型棉蚜在5种杂草及棉花上的形态参数
取食荠菜的Hap1型棉蚜体型最大,其体长平均达1 100 μm以上,取食飞蓬、阿拉伯婆婆纳、猪殃殃的棉蚜体长在1 000~1 100 μm之间。取食棉花叶片的Hap1型棉蚜平均体长为985 μm。取食飞廉叶片的棉蚜平均体长为916 μm,明显小于取食其他5种寄主植物的棉蚜。在体宽方面,取食飞蓬叶片的Hap1型棉蚜平均体宽最小,为500 μm左右,远低于取食其他5种寄主植物的棉蚜(图2)。
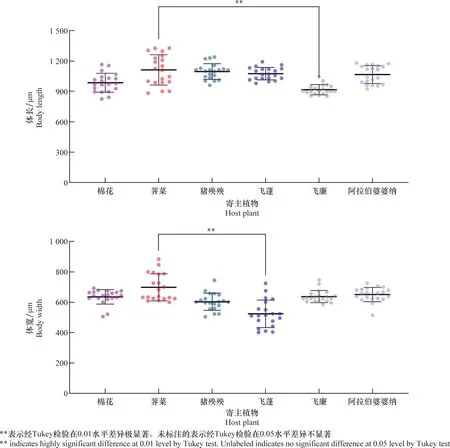
图2 Hap1型棉蚜取食6种不同植物后子一代的体长和体宽
3 讨论
棉蚜是棉花上的主要害虫之一,具有体型小、世代短、繁殖快和适应性强的特点。在冬季比较寒冷的区域,棉蚜产生性蚜,以卵越冬。在其他季节,棉蚜以孤雌生殖方式进行繁衍,在条件适宜的情况下,棉蚜可迅速扩大种群规模,对农业生产造成严重为害。在棉花苗期和蕾铃期,棉蚜为害较严重,生产上称为苗蚜和伏蚜[15]。一般情况下,苗蚜的发生频率和为害程度要高于伏蚜[16]。在以往研究中,棉花苗蚜的虫源被认为是来自于木槿、石榴和花椒等越冬寄主。笔者通过多年的观察发现,越冬寄主上棉蚜的种群数量在棉花种植之前已达到很高的水平,而且有翅蚜的产生也要早于棉花播种时间,推测在棉蚜从越冬寄主迁飞到棉田之前,种群已经扩散。早春时节,主要农作物小麦和油菜不是棉蚜的适宜寄主,而早春杂草生长茂盛,可能会作为棉蚜的迁移对象。从而导致早春杂草上繁殖的棉蚜和越冬寄主上的棉蚜共同构成苗蚜的虫源。
杂草是生态系统中种类较为丰富的一类生物,杂草作为棉蚜寄主已有较多的报道。李昌学[17]在山西进行了棉蚜接种试验,表明夏至草(后续研究表明文献中的夏枯草Prunellavulgaris应为夏至草,夏枯草不能作为棉蚜的寄主[18])、鼠曲草Pseudognaphaliumaffine、紫花地丁Violaphilippica、刺芋Lasiaspinosa、芥菜等可作为棉蚜的寄主。朱弘复等[19]根据接种试验,确定车前Plantagoasiatica是棉蚜的寄主。近期,景玉玺等[20]通过生物测定试验表明棉蚜在龙葵、裂叶牵牛Ipomoeanil、刺儿菜和棉花等4种寄主植物上均可完成世代发育,棉蚜在龙葵上的适合度、嗜食性最好,其次是裂叶牵牛,在刺儿菜上适应性最差。本研究选择5种早春优势杂草,通过生物测定试验表明,Hap1型棉蚜可以在阿拉伯婆婆纳、荠菜、猪殃殃、飞廉和飞蓬上建立种群。尽管Hap1型棉蚜在5种杂草上的生长和繁殖能力低于取食棉花的种群,但杂草是春季优势植物,分布广,种类丰富,是不可忽视的苗蚜虫源地。
Hap1型棉蚜在早春杂草上能够建立种群,完成生活史,可能与早春季节杂草的繁盛和棉蚜最适寄主的缺失相关。取食经历影响昆虫对寄主的适合度[21],由于本试验所用虫源为使用棉花饲养多代的种群,棉蚜可能对棉花产生寄主偏好性,从而导致Hap1型棉蚜对5种杂草上的适合度低于对棉花的适合度。后续通过多代饲养,结合生物测定可进一步明确早春杂草在棉蚜种群周年繁殖中的作用。
4 结论
通过生物测定的方法,本研究证实Hap1型棉蚜可以利用5种杂草完成世代发育。结合田间观察,推测田间早春杂草可能是棉花苗蚜的虫源之一,生产实践中可通过铲除田间杂草切断棉蚜传播途径,降低棉田苗蚜的虫口基数。研究结果对于填补棉蚜周年生活史中的寄主转移规律和指导棉田苗蚜的防治具有重要意义。
猜你喜欢
农药学学报(2022年6期)2022-12-27 12:00:28
湖北农业科学(2021年22期)2021-12-13 13:00:36
环境昆虫学报(2021年1期)2021-03-30 02:25:20
新疆农业科学(2020年11期)2020-10-27 09:05:32
现代园艺·综合版(2017年1期)2017-07-14 16:37:59
江苏农业科学(2017年7期)2017-05-23 23:10:25
中国稻米(2017年2期)2017-04-28 08:00:06
农业与技术(2016年19期)2016-12-12 02:23:50
中国稻米(2016年2期)2016-06-29 09:53:29
现代农业(2016年4期)2016-02-28 18:42:12
