
水位变动影响下三峡库区汉丰湖鸟类群落及多样性
2018-03-22 11:33:34刁元彬袁兴中张乔勇黄亚洲岳俊生周李磊
生态学报 2018年4期
刁元彬,刘 红,*, 袁兴中, 张乔勇, 熊 森, 黄亚洲, 岳俊生, 周李磊
1 煤矿灾害动力学与控制国家重点实验室, 重庆 400030 2 三峡库区生态环境教育部重点实验室, 重庆 400030 3 重庆大学资源及环境科学学院, 重庆 400030 4 重庆开州澎溪河湿地自然保护区管理局, 重庆 405400
三峡水库蓄水后,由于防洪、清淤及航运等需求,三峡水库实行“蓄清排浑”的运行方式,即每年汛期(6—9月)将水位降至145m,放水排沙;汛期过后将水位抬升至175m,拦蓄清水。因此,三峡水库正常蓄水运行后,在145—175m高程的库区两岸,形成与天然河流涨落季节相反、水位落差高达30m、面积达348.9km2的水库消落带[1]。反季节、高水位落差导致三峡库区原有生态系统发生显著变化,对生物多样性产生强烈胁迫[2]。鸟类作为生态系统中的重要组成部分,是生态系统健康水平的指示类群[3- 5]。目前三峡水库生物多样性研究多集中于藻类、底栖动物、鱼类、高等维管植物等生物类群[6- 8]。对三峡水库鸟类生态学的研究比较匮乏,迄今为止,仅有苏化龙等[9- 11]、张家驹等[12]、冉江洪等[13]等对库区鸟类进行过本底调查,但对三峡水库蓄水后鸟类群落的变化、季节性水位变化与鸟类群落结构之间的相互关系缺乏研究。本文以位于三峡库区腹心的重庆开州区汉丰湖为研究区域,选择不同水位时期,进行鸟类群落结构及其多样性的调查研究,探讨大型水库蓄水后鸟类群落及多样性对季节性水位变化的响应关系,旨在积累三峡库区生物多样性变化的长期数据,为国内外大型水库的生物多样性保护及其管理提供科学依据。
1 研究区域与研究方法
1.1 研究区域
研究区域汉丰湖位于三峡库区腹心的重庆开州区,是长江一级支流澎溪河回水末端,地理坐标为108°18′41.35′′—108°′27′41.84″E,31°09′10.09′′—31°11′16.39′′N。研究区域属亚热带季风气候,多年平均气温18.5℃,多年平均降水量1385mm。受三峡水库水位调控影响,环汉丰湖形成了水位变幅接近22.5m的消落带[14]。为减缓汉丰湖大面积消落带对开州城区生态环境的不利影响,2007年,在重庆开州新城区下游4.5km处修建水位调节坝,拟将消落带水位变幅由22.5m降至4.72m,并形成具有独特水位变动的“城市内湖”—汉丰湖。汉丰湖水位调节坝的调控方式为:冬季,当三峡水库水位上涨至175m,调节坝闸门打开,汉丰湖与三峡水库同水位,即175m高水位运行;夏季,当三峡水库坝前水位下降至145m时,闸门关闭,汉丰湖水位维持在170.28m。汉丰湖水位调节坝自2012年建成后,迄今为止尚未下闸蓄水,因此一直保持着与三峡水库同步的水位变化。2017年汉丰湖将首次尝试在夏季三峡水库低水位时期下闸蓄水到170.28m水位高程。
针对汉丰湖的水位季节性变化,为了保障汉丰湖水环境安全,改善汉丰湖周城市人居环境质量,环汉丰湖消落带及其上的湖岸区实施了系列湿地修复工程,包括基塘工程(汉丰湖南岸石龙船大桥至游泳馆、汉丰湖南岸芙蓉坝、汉丰湖北岸石龙船大桥至头道河口)、湿地多塘工程(头道河河口)、林泽工程(东河河口、乌杨坝)及多带多功能生态护坡(乌杨坝)。
1.2 调查方法
2015年3月至2016年2月对汉丰湖鸟类群落及其生境进行了调查,根据水位变动特点、代表性生境类型,共设置了6个固定定量调查样点(表1, 图1)。S1—S4样点的调查面积设置为10hm2,S5—S6为20hm2。 同时根据汉丰湖生境状况,对整个湖区进行了鸟类的定性调查。
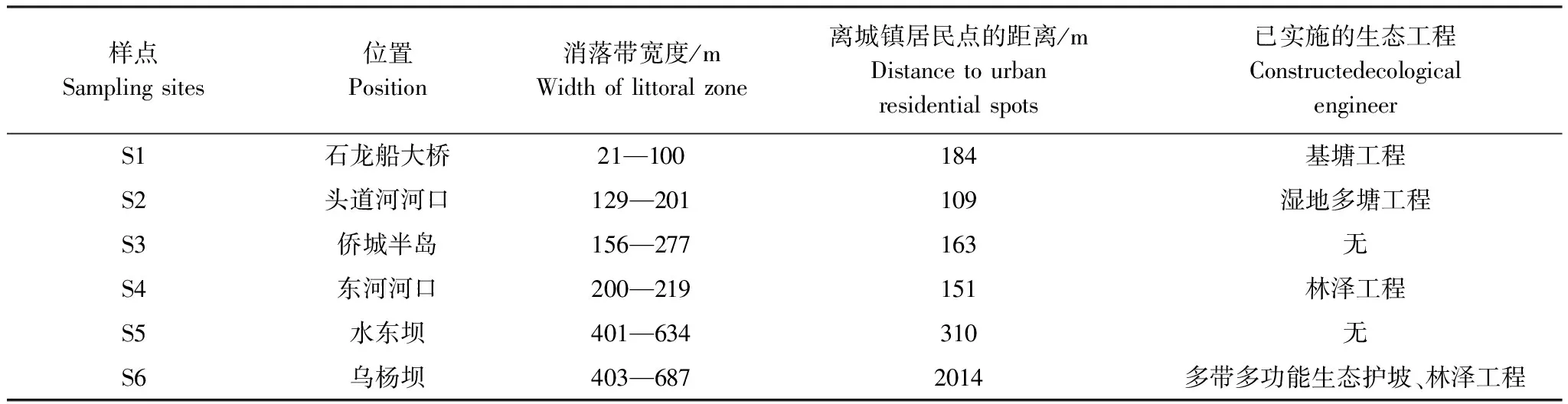
表1 汉丰湖调查样点环境特征
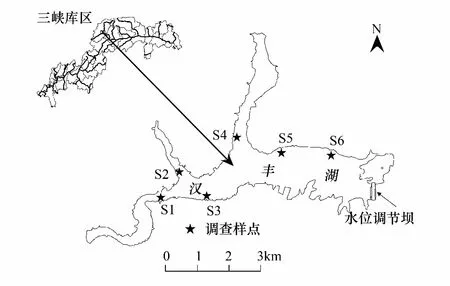
图1 调查样点位置Fig.1 The location of sampling sites in Hanfeng Lake
冬季调查分两次进行,即冬季Ⅰ(2015年12月最高水位期,水位高程为174.50m)和冬季Ⅱ(2016年2月开始退水后,代表水位消退初期,水位高程为170.01m)。春、夏及秋季定量调查分别选择2015年4、8、10月进行。每个月调查3次。选择晴朗无风的天气,在鸟类活动的高峰段(7:00—10:00;16:00—18:00),使用8×42倍双筒望远镜及20—60倍单筒望远镜进行观察,记录鸟类的种类、数量、行为及生境特征,鸟类识别主要参照《中国鸟类野外手册》[15],鸟类分类参照《中国鸟类分类与分布名录》[16]。
1.3 功能群划分
参考Paszkowski和Tonn[3]、Gatto等[17]、Ortega-lvarez等[18]的研究,并结合野外对鸟类取食习性的实际观察,将鸟类划分为4种取食功能群:(1)肉食性鸟,取食鱼、螺、贝类及小型脊椎动物等;(2)食虫鸟,主要取食昆虫;(3)植食性鸟,主要取食植物体果实、种子、谷物、根茎、块茎等;(4) 杂食性鸟,取食小型动物、植物体等多种食物。
1.4 数据分析
优势度测定采用Berger-Parker 优势度指数(I)[19],计算公式为:
I=Ni/N
(1)
优势度等级按照如下标准划分:优势种(>10%),常见种(1%—10%),稀有种(<1%)[20]。
鸟类多样性采用Shannon-Wiener指数和Pielou均匀度指数(J)测定[19],计算公式为:
(2)
Pielou均匀度指数:
J=H/lnS
(3)
式中,S为物种数;N为总个体数;Ni为第i个物种的个体数量;Pi为第i种个体数量占群落总个体数量的比例。
鸟类群落相似性采用Sørenson 相似性系数测定,计算公式为:
c=2j/(a+b)
(4)
式中,j为2个季节共有的鸟类种类数,a为第一个季节的鸟类种类数,b为第二个季节的鸟类种类数[21]。
用One-way ANOVA对群落数量指标进行差异显著性检验。若差异显著,采用Duncan法进行多重比较。数据采用SPSS 18.0进行统计处理。显著性水平取0.05。
采用除趋势对应分析法(Detrended correspondence analysis, DCA)分析越冬鸟类的空间分布特征及其栖息的生境特点[22-23]。剔除相对多度小于0.1%的物种,以6个调查样点在冬季Ⅰ和冬季Ⅱ的28个物种的多度数据为基础,构建28×12二维矩阵进行除趋势对应分析。DCA分析及制图用CANOCO 4.5软件完成。
2 结果与分析
2.1 群落组成
本研究共记录鸟类97种,隶属12目32科。其中,雀形目鸟类最多,有43种,占鸟类种数的44.33%;其次为雁形目和鸻形目,有18种和13种,分别占鸟类种数的18.56%、13.40%。其他类群鸟类种数均低于10种。湿地鸟类49种,主要包括雁鸭类(18种)、鸻鹬类(13种)、鹭类(7种)、秧鸡类(5种)。在居留类型上,留鸟41种,占鸟类种数的42.27%;冬候鸟30种,占鸟类种数的30.93%;夏候鸟和旅鸟种类较少,所占鸟类种数的比例分别为14.43%和11.34%;迷鸟1种,为斑头雁(Anserindicus)。国家Ⅱ级野生保护动物3种,分别为普通鵟(Buteojaponicus)、鸳鸯(Aixgalericulata)及小天鹅(Cygnuscolumbianus)。红胸田鸡(Porzanafusca)、蓝胸秧鸡(Gallirallusstriatus)为重庆市新记录种。
优势种种类及其个体数量随季节变化明显,并表现出与水位变动一致的变化关系。8月汉丰湖低水位期,优势种为白鹭(Egrettagarzetta)和棕头鸦雀(Sinosuthorawebbiana),其中白鹭在汉丰湖小规模集群,仅在S3样点就观测到75只。10月到次年2月汉丰湖处于高水位期,秋季与冬季Ⅰ、冬季Ⅱ的优势种不同;秋季的优势种为斑嘴鸭(Anaszonorhyncha)和麻雀(Passermontanus);冬季Ⅰ的优势种为绿头鸭(A.platyrhynchos)、罗纹鸭(A.falcata)和骨顶鸡(Fulicaatra);冬季Ⅱ的优势种为绿头鸭和骨顶鸡。冬季Ⅰ的优势种个体数量最高,占当月鸟类总数量的比例为80.05%。
汉丰湖鸟类物种丰度和多度随季节变化明显。夏季的物种丰度最高,为34种;其次为秋季32种;冬季Ⅰ30种,冬季Ⅱ28种;春季的物种丰度最低,仅27种。冬季Ⅱ和春季的物种丰度分别与夏季、秋季、冬季Ⅰ的差异显著(P<0.05);冬季Ⅱ和春季之间差异不显著(P>0.05);夏季、秋季和冬季Ⅰ之间的差异不显著(P>0.05)。
冬季Ⅰ汉丰湖蓄水到最高水位,水域面积为14.8km2,调查到鸟类数量1048只,其中水鸟1024只;鸭类是主要生态类群,有848只。夏季汉丰湖低水位期,水域面积大大缩小,约4km2,调查到水鸟121只;雀形目鸟类及鹭类是主要的生态类群,分别为254只、101只。冬季Ⅱ与冬季Ⅰ的鸟类多度差异不显著(P>0.05);冬季Ⅰ和冬季Ⅱ与春季、夏季、秋季之间鸟类多度差异显著(P<0.05);春季、夏季、秋季之间的多度差异不显著(P>0.05)。
2.2 群落多样性及相似性
2015年夏季汉丰湖为低水位期,2015年10月至2016年2月为高水位期,水位高程随季节变化明显。夏季、秋季、春季的Shannon-Wiener指数和Pielou均匀度指数显著高于冬季Ⅰ和冬季Ⅱ(图2,图3)。冬季Ⅰ和冬季Ⅱ的多样性指数分别与春季、夏季、秋季之间的差异显著(P<0.05);冬季Ⅰ和冬季Ⅱ之间差异不显著(P>0.05);春季、夏季、秋季之间差异不显著(P>0.05)。
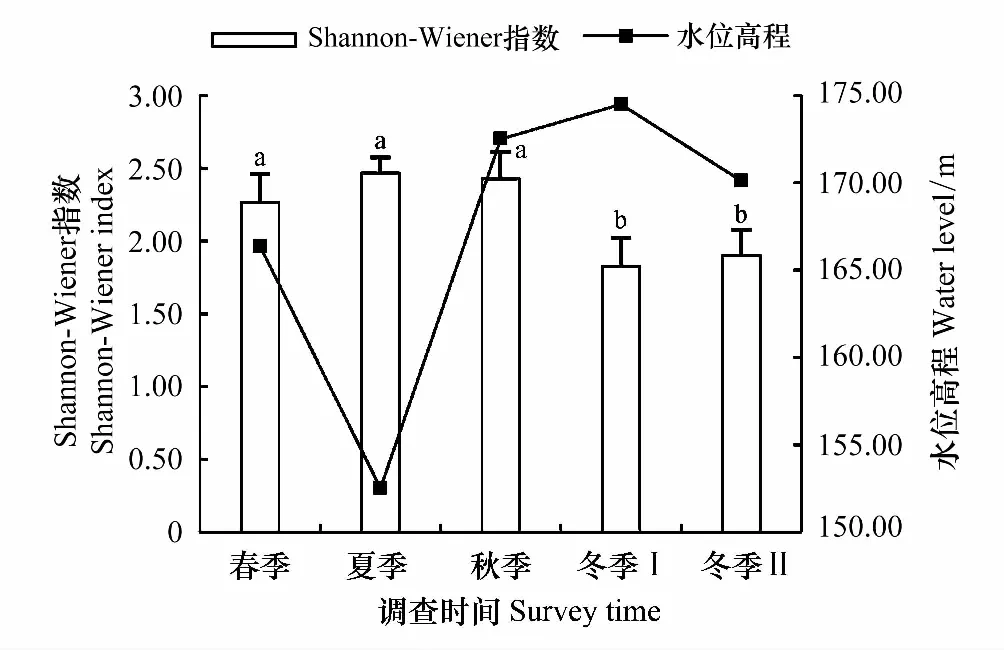
图2 不同季节汉丰湖鸟类群落Shannon-Wiener指数(均值±标注差)及对应的水位高程 Fig.2 Species Shannon-Wiener index in different seasons of Hanfeng Lake (Mean±SE) and water level不同字母代表显著性水平0.05的差异性
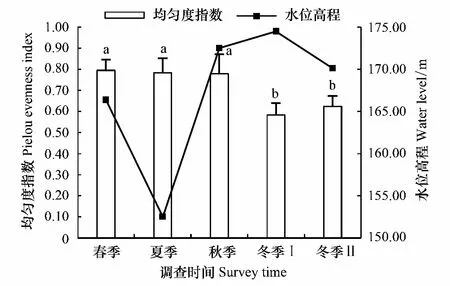
图3 不同季节汉丰湖鸟类群落Pielou均匀度指数(均值±标注差)及对应的水位高程Fig.3 Pielou evenness index of avian communities in different seasons of Hanfeng Lake (Mean±SE) and water level不同字母代表显著性水平0.05的差异性
由表2可知,冬季Ⅰ汉丰湖高水位(174.50m)期,与春季、夏季、秋季的鸟类群落结构相似性系数低。夏季与冬季Ⅰ仅有10种鸟类相同,鸟类群落结构相似性系数最低,为0.31。8月低水位期(152.50m)主要为雀形目鸟类和鹭类;12月高水位期,主要为雁鸭类和雀形目鸟类,鸟类群落结构组成差异明显。12月和2月群落相似性系数较高,为0.83。
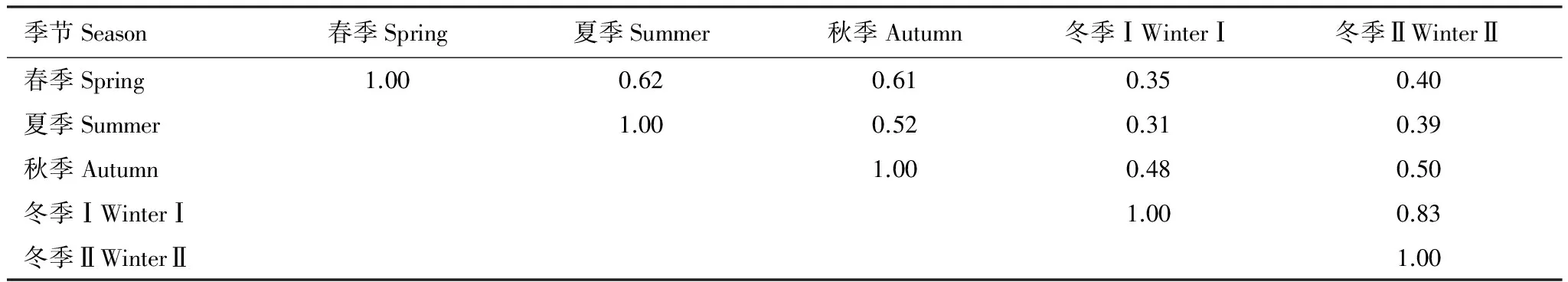
表2 汉丰湖不同季节鸟类群落相似性系数
冬季Ⅰ为2015年12月,冬季Ⅱ为2016年2月
2.3 越冬鸟类空间分布
DCA排序轴能够反映一定的环境梯度,通过对环境因子与排序轴的相关分析,可以识别与排序轴显著相关的环境因子,反映物种分布与环境因子之间的关系[24-25]。图4为汉丰湖鸟类12月和2月的DCA二维排序图。越冬鸟类的空间分布格局有差异。第一排序轴从左至右基本反映了在冬季S1—S6样点水面宽度增加和距离城镇居民点的距离降低的趋势。第一排序轴反映了越冬的雁鸭类集中分布在水面最为开阔、人为干扰强度最低的S6样点。鸭科鸟类冬季在S6区域集群,个体数量占越冬鸟类总数量的80.5%。第一排序轴特征值分为0.712,解释了36.8%的鸟类分布特征。第二排序轴从下至上基本反映了人为干扰强度从低到高的变化趋势。麻雀、白鹡鸰(Motacillaalba)、纯色山鹪莺(Priniainornata)等雀形目鸟类零散分布在人为干扰较强的S1—S4样点。第二排序轴特征值分为0.231,第二排序轴解释了11.9%的分布特征。在冬季Ⅰ与冬季Ⅱ,S5的鸟类组成有差异。
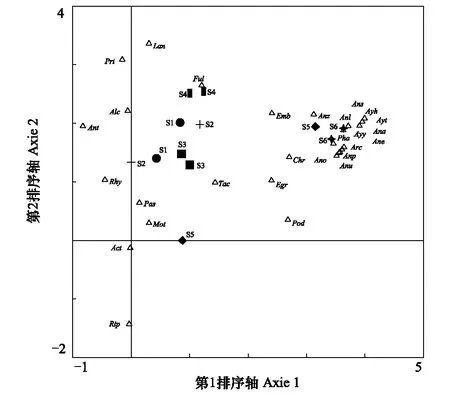
图4 汉丰湖越冬鸟类DCA分析二维排序图 Fig.4 Detrended correspondence analysis ordination of wintering birds in Hanfeng Lake图中不同的缩写对应不同鸟类的拉丁名:Pha: 普通鸬鹚, Phalacrocorax carbo; Alc: 普通翠鸟, Alcedo atthis; Tac: 小, Tachybaptus ruficollis; Pod: 凤头, Podiceps cristatus; Ful: 骨顶鸡, Fulica atra; Chr: 红嘴鸥, Chroicocephalus ridibundus; Act: 矶鹬, Actitis hypoleucos; Arc: 苍鹭, Ardea cinerea; Egr: 白鹭, Egretta garzetta; Ana: 赤膀鸭, Anas strepera; Ans: 罗纹鸭, Anas falcate; Anp: 赤颈鸭, Anas Penelope; Anl: 绿头鸭, Anas platyrhynchos; Anz: 斑嘴鸭, Anas zonorhyncha; Anu: 针尾鸭, Anas acuta; Ano: 花脸鸭, Anas Formosa; Ane: 绿翅鸭, Anas crecca; Ayt: 红头潜鸭, Aythya ferina; Ayh: 白眼潜鸭, Aythya nyroca; Ayy: 凤头潜鸭, Aythya fuligula; Pas: 麻雀, Passer montanus; Emb: 小鹀, Emberiza pusilla; Lan: 棕背伯劳, Lanius schach; Pri: 纯色山鹪莺, Prinia inornata; Rip: 崖沙燕, Riparia riparia; Mot: 白鹡鸰, Motacilla alba; Ant: 树鹨, Anthus hodgsoni; Rhy: 红尾水鸲, Rhyacornis fuliginosa
2.4 功能群动态变化
汉丰湖鸟类取食功能群组成随季节不同而变化显著,由此反映了功能群组成与水位变化之间的关系(图5,图6)。食虫鸟主要由雀形目鸟类和鸻鹬类组成,在春季和秋季的相对丰度(即各功能群的鸟类种类组成)最高,分别为59.26%和46.88%;夏季和冬季略低。肉食性鸟类由鹭类、类、鸬鹚等组成,相对丰度随季节变化不明显。杂食性鸟类主要由雀形目鸟类和秧鸡类组成,相对丰度随季节变化不明显。植食性鸟类主要由鸭科鸟类和雀形目鸟类组成,随季节变化极为显著,冬季相对丰度最高,两个水位高程时期(即12月174.50m水位高程,和2月170.01m水位高程)分别为26.67%和35.71%;春季最低,仅为3.70%。
汉丰湖鸟类功能群的相对多度(即各功能群的鸟类数量组成)随季节变化的规律与相对丰度的变化明显不同。肉食性鸟类相对多度夏季最高,为31.35%,其余季节均较低。食虫鸟的相对多度春季和夏季较高,分别为49.77%和40.34%,冬季下降至最低,分别为3.39%和3.43%。植食性鸟相对多度春季最高,其余季节略低。植食性鸟相对多度冬季最高,两个水位高程时期分别为72.53%、68.13%;春季和夏季极低,分别为1.17%和8.14%。夏季和冬季的鸟类功能群的相对多度差异极为明显。
3 讨论
3.1 水位变化与鸟类多样性
明水面、滩涂、植被是影响湿地鸟类群落多样性及空间分布格局的3个重要因子[26]。水位变动是三峡水库蓄水后汉丰湖生态系统变化的主要驱动因子。与三峡水库水位变化相一致,汉丰湖的季节性水位变化导致明水面、泥滩、植被等主要因子随水位高程涨落而发生动态变化,进而影响汉丰湖鸟类多样性及空间分布格局。
8月汉丰湖低水位期,鸟类群落多样性指数和物种丰富度最高、功能群类型最多,主要原因如下:(1)研究范围内鸟类生境在一年中的不同季节发生明显的变化,这种变化与季节性水位变化密切相关。夏季当三峡水库水位降到145m时,汉丰湖水位降至最低,水域面积缩小至一年中最小的时期,约4km2。相比其他月份,大面积的河漫滩和沙洲出露,并有洼地、沟渠、小型水塘、小型岛屿、砾石滩等多种微生境结构。多样的微生境及其组合类型为白鹭、彩鹬(Rostratulabenghalensis)、红胸田鸡、矶鹬(Actitishypoleucos)等肉食性鸟和食虫鸟提供了食物资源和栖息环境。(2)以狗牙根(Cynodondactylon)、苍耳(Xanthiumstrumarium)、胡枝子(Lespedezatomentosa)等为优势种组成的消落带植被,在夏季低水位时期多处于花期或果期,草籽和植物群落内的昆虫资源是一年中最为丰富的时期,为棕头鸦雀、纯色山鹪莺、黑卷尾(Dicrurusmacrocercus)等食虫鸟提供了优良的栖息地和食物资源。 (3)汉丰湖部分湖岸(如汉丰湖南岸石龙船大桥至游泳馆、汉丰湖南岸芙蓉坝、汉丰湖北岸石龙船大桥至头道河河口)营造有面积不等、形态各异的湿地基塘,塘内鱼类、螺类、水生昆虫等食物资源丰富,挺水植物团块状种植,形成良好的庇护地,是蓝胸秧鸡、黑水鸡(Gallinulachloropus)、白胸苦恶鸟 (Amaurornisphoenicurus)等涉禽的繁殖场所。
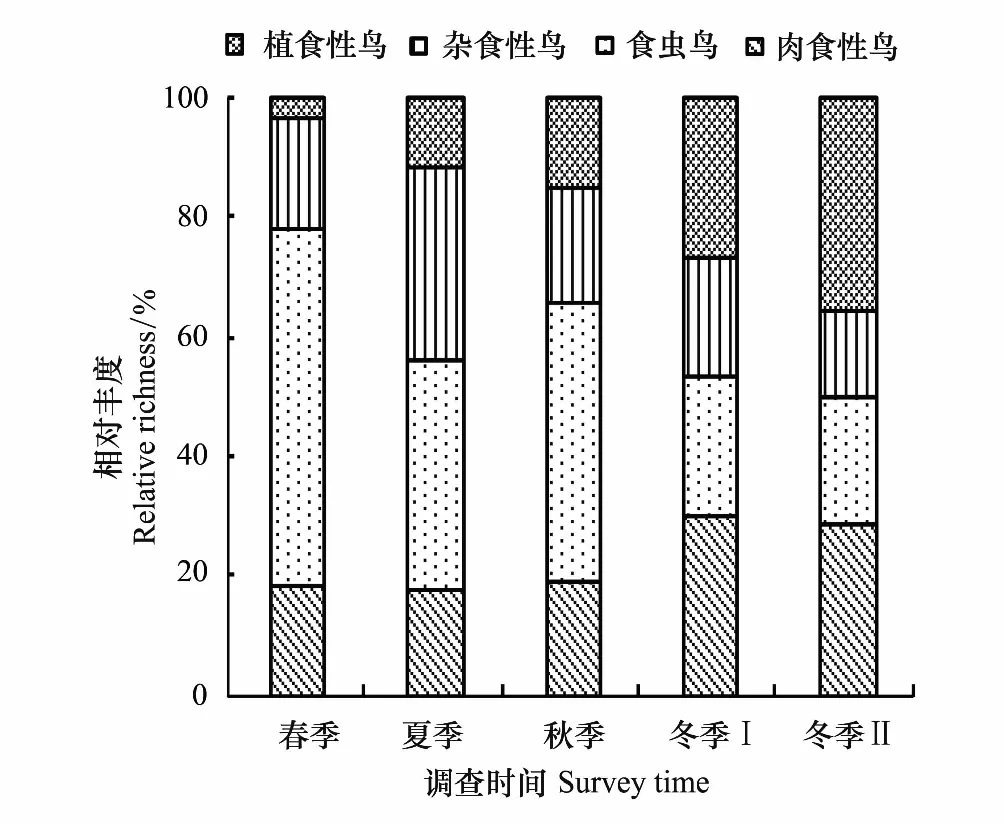
图5 汉丰湖鸟类取食功能类群相对丰度的季节变化 Fig.5 Temporal changes of avian feeding functional groups′ relative richness in different seasons of Hanfeng Lake
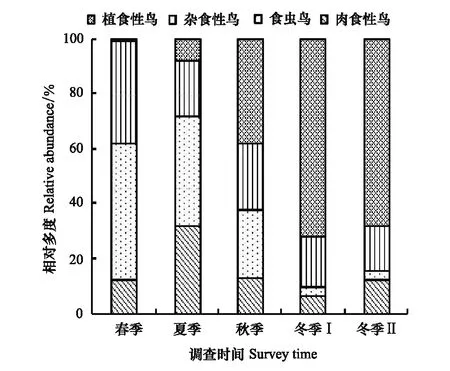
图6 汉丰湖鸟类取食功能类群相对多度的季节变化 Fig.6 Temporal changes of avian feeding functional groups′ relative abundance in different seasons of Hanfeng Lake
汉丰湖12月达到最高水位175m,水域面积宽阔,水面达14.8km2,滩涂被淹没,大部分植被淹没水下,生境结构单一,物种丰富度最低。夏季和冬季鸟类群落组成及结构差异明显。从2015年8月到2015年12月、2016年2月,研究区域内在草丛中活动的食虫鸟、植食性鸟和肉食性鸟类物种丰度和多度明显下降;到了冬季,这3种功能群类型的鸟类趋向周边农耕地分布取食越冬。冬季汉丰湖形成大水面吸引绿头鸭、罗纹鸭、绿翅鸭等植食性鸟类集群越冬。骨顶鸡善于潜水,能觅食水下25cm的植物体、小型鱼类及水生昆虫,适宜深水环境,因此也是冬季的优势种[27-28]。
4月是汉丰湖水位消退的末期,10月是汉丰湖水位上涨期,此阶段为水位变化动态时期。4月和10月生境与8月相似,因此鸟类群落多样性指数、物种丰度、多度等与8月接近。4月与10月是鸻鹬类迁徙时期,滩涂是鸻鹬类在迁徙时重要的停歇地和觅食地[26,29]。但在研究期内仅观测到5种鸻鹬,鸻鹬类种类及种群数量少,主要原因有:(1)汉丰湖4月水位下降,水域面积缩小,新生成大面积泥滩。由于光热条件适宜,泥滩水分蒸发,含水量减少,泥滩内的狗牙根、苍耳、合萌(Aeschynomeneindica)等植物的繁殖体迅速萌发生长,极快覆盖泥滩生境。汉丰湖泥滩出现的时间较短,不能够给鸻鹬类提供稳定的食物资源。(2)2015年4月汉丰湖流域降雨量比往年大,形成洪水,加大了对泥滩的冲刷、侵蚀,不适宜鸻鹬类停歇、觅食。(3)2015年10月由于水位抬升,夏季的滩涂被淹没;水位高程大于170m时滩涂生境极少,零星分布,难以吸引鸻鹬类停歇觅食。
3.2 冬季鸟类空间分布格局
鸟类多样性及其空间分布格局与生境结构复杂性密切相关,受水域面积、滩涂、湿地植被及周边土地利用类型等多种环境因子影响[30]。冬季高水位期,汉丰湖有库湾、河口、半岛、开阔大水面等生境结构。汉丰湖6个样点由于生境结构与周边环境不同,冬季鸟类空间分布显著不同。
冬季汉丰湖的鸟类集中分布于S6样点,越冬水鸟种群数量显著高于其他样点,主要原因有:(1)S6形成水域面积近100hm2、宽度超过1km的大水面,是河鸭属、麻鸭属、潜鸭属适宜的越冬栖息地。库岸坡度平缓,高程172m以上的植物残体和小型水生生物成为越冬雁鸭类的重要食物资源。(2)庇护性是鸟类在生境内进行觅食、求偶、繁殖、休憩等行为的重要条件。在汉丰湖北岸的乌杨坝,以香樟(Cinnamomumchingii)和斑茅(Saccharumarundinaceum)为优势种构成的多带多功能生态护坡,其长度超过1km,宽度超过50m,是优良的生境隔离带,削弱了外界人为干扰强度。在生态护坡以下,水位高程为172m的消落带区域实施了以池杉(Taxodiumdistichumvar.imbricatum)、落羽杉(Taxodiumdistichum)、江南桤木(Alnustrabeculosa)为优势种的林泽工程,形成了宽度超过10m、与库岸平行的带状林泽(耐水淹的木本群落)。在蓄水高程达到175m时,林泽露出水面超过3m。林泽树冠层为鸬鹚(Phalacrocoraxcarbo)、白鹭、苍鹭等水鸟提供了停栖地,而林内水面则是斑嘴鸭、罗纹鸭、绿头鸭等的庇护、休憩场所。S6是生态护坡和林泽的复合庇护生境,有利于屏蔽周边的人为干扰,极大提高了生境的隐蔽性。(3)S6周边为自然荒草地,人为干扰强度很小。
S1—S5样点水面宽度、水域面积低于乌杨坝,周边属于城市滨湖公园、居民区,人为干扰强度高,鸟类物种丰度和种群数量均较低。
汉丰湖2月份水位已经开始降低,水域面积减少,在169—175m高程区内形成大小不一的小型水塘,滞留了不少小型鱼类,是苍鹭、白鹭等涉禽重要的食物资源,因此鹭类种群数量增加。由于水域面积降低和食物资源减少,罗纹鸭种群数量减少。2月份由于水位下降约5m,S5生境结构改变,从大水面变为小型岛屿、泥滩及浅水区等多种结构构成的复合生境,吸引大量绿头鸭、骨顶鸡等水鸟栖息,物种丰度和个体数量显著增加。
3.3 汉丰湖鸟类生境恢复与营造
从2013年6月至2016年2月,在汉丰湖记录到鸟类近130种,湿地鸟类超过一半,包括中华秋沙鸭(Mergussquamatus)、小天鹅(Cygnuscolumbianus)、鸳鸯(Aixgalericulata)等珍稀濒危水鸟,成为三峡库区乃至长江上游重要的水鸟越冬地。2017年汉丰湖水位调节坝将正式下闸蓄水,将最低水位维持在170.28m,水位落差由过去的22.5m降低到4.72m。由此,过去夏季低水位期的河漫滩、沙洲、洼地、砾石滩等生境将大部分消失,将导致鹭科、秧鸡科、彩鹬科等涉禽的栖息地减少或丧失。如何针对汉丰湖鸟类进行生境恢复和重建,降低水位变化对鸟类的影响是必须高度重视的生物多样性保护问题。
结合汉丰湖鸟类群落组成及时空变化特征,重点针对繁殖鸟和越冬鸟类设计生境。繁殖鸟与栖息地动态变化最为密切[31]。红胸田鸡、蓝胸秧鸡是汉丰湖内主要的繁殖鸟,可以沿湖岸营造形态不同,面积不一、水位深度不同的基塘,团块状种植乡土湿地植物,维持三分之二的明水面面积,为秧鸡科、鹭科鸟类营造繁殖庇护生境。
岛屿是河流、湖泊、水库等水体中的重要结构单元,是涉禽和游禽优良的庇护、繁殖生境。结合汉丰湖水鸟的时空分布特征,在S6样点林泽外营造生境岛,沿高程梯度从湖内到湖岸高地形成水域-岛链-林泽-生态护坡的复合生境结构,为鹭类和越冬雁鸭类在汉丰湖营造隐蔽的生境空间。
[1] 张虹. 三峡库区消落带土地资源特征分析. 水土保持通报, 2008, 28(1): 46- 49.
[2] Wu J G, Huang J H, Han X G, Gao X M, He F L, Jiang M X, Jiang Z G, Primack R B, Shen Z H. The Three Gorges Dam: an ecological perspective. Frontiers in Ecology and the Environment, 2004, 2(5): 241- 248.
[3] Paszkowski C A, Tonn W M. Foraging guilds of aquatic birds on productive boreal lakes: environmental relations and concordance patterns. Hydrobiologia, 2006, 567(1): 19- 30.
[4] 陆健健. 河口生态学. 北京: 海洋出版社, 2003.
[5] Wu C F, Lin Y P, Lin S H. A hybrid scheme for comparing the effects of bird diversity conservation approaches on landscape patterns and biodiversity in the Shangan sub-watershed in Taiwan. Journal of Environmental Management, 2011, 92(7): 1809- 1820.
[6] New T, Xie Z Q. Impacts of large dams on riparian vegetation: applying global experience to the case of China′s Three Gorges Dam. Biodiversity Conservation, 2008, 17(13): 3149- 3163.
[7] Park Y S, Chang J B, Lek S, Cao W X, Brosse S. Conservation strategies for endemic fish species threatened by the three gorges dam. Conservation Biology, 2003, 17(6): 1748- 1758.
[8] Yan Q Y, Yu Y H, Feng W S, Yu Z G, Chen H T. Plankton community composition in the Three Gorges Reservoir Region revealed by PCR-DGGE and its relationships with environmental factors. Journal of Environmental Sciences, 2008, 20(6): 732- 738.
[9] 苏化龙, 林英华, 张旭, 于长青, 冉江洪, 隆廷伦, 刘绍英. 三峡库区鸟类区系及类群多样性. 动物学研究, 2001, 22(3): 191- 199.
[10] 苏化龙, 马强, 胥执清, 张小蓉. 三峡水库蓄水139m前后江面江岸冬季鸟类动态. 动物学杂志, 2005, 40(1): 92- 95.
[11] 苏化龙, 肖文发, 王建修, 胥执清, 李望洪, 刘富国, 刘小云, 张小蓉, 王英. 三峡库区蓄水前后冬季小江水面及河岸鸟类种群波动调查. 西南师范大学学报: 自然科学版, 2012, 37(11): 41- 48.
[12] 张家驹, 熊铁一, 罗佳, 罗明澎, 吴大均. 三峡工程对库区鸟类资源的影响评价. 自然资源学报, 1991, 6(3): 262- 273.
[13] 冉江洪, 刘少英, 林强, 刘世昌, 王跃招. 重庆三峡库区鸟类生物多样性研究. 应用与环境生物学报, 2001, 7(1): 45- 50.
[14] 李波, 袁兴中, 熊森, 刘红, 岳俊生, 陶德均. 城市消落带景观基塘系统设计初探——以重庆开县汉丰湖为例. 重庆师范大学学报: 自然科学版, 2013, 30(6): 51- 54.
[15] 约翰·马敬能, 卡伦·菲利普斯. 中国鸟类野外手册. 何芬奇, 译. 长沙: 湖南教育出版社, 2000.
[16] 郑光美. 中国鸟类分类与分布名录(第二版). 北京: 科学出版社, 2011.
[17] Gatto A, Quintana F, Yorio P. Feeding behavior and habitat use in a waterbird assemblage at a marine wetland in Coastal Patagonia, Argentina. Waterbirds, 2008, 31(3): 463- 471.
[19] 马克平, 刘玉明. 生物群落多样性的测度方法I:α多样性的测度方法(下). 生物多样性, 1994, 2(4): 231- 239.
[20] 徐玲. 崇明东滩湿地植被演替不同阶段鸟类群落动态变化的研究[D]. 上海: 华东师范大学, 2004.
[21] 孙儒泳. 动物生态学原理(第三版). 北京: 北京师范大学出版社, 2006.
[22] 张金屯. 数量生态学. 北京: 科学出版社, 2003.
[23] 阮得孟, 孙勇, 程嘉伟, 刘大伟, 鲁长虎. 盐城自然保护区新洋港河口不同生境冬季鸟类群落组成及其梯度变化. 生态学报, 2015, 35(16): 5437- 5448.
[24] 王强, 袁兴中, 刘红. 山地河流浅滩深潭生境大型底栖动物群落比较研究——以重庆开县东河为例. 生态学报, 2012, 32(21): 6726- 6736.
[25] 苏日古嘎, 张金屯, 张斌, 程佳佳, 田世广, 张钦弟, 刘素军. 松山自然保护区森林群落的数量分类和排序. 生态学报, 2010, 30(10): 2621- 2629.
[26] 葛振鸣. 长江口滨海湿地迁徙水禽群落特征及生境修复策略[D]. 上海: 华东师范大学, 2007.
[27] 哈丽亚, 程鲲, 宗诚, 李金波. 骨顶鸡日行为活动对游憩干扰的反应. 生态学杂志, 2014, 33(7): 1860- 1866.
[28] 桑莉莉, 葛振鸣, 裴恩乐, 徐骁俊, 姜姗, 王天厚. 崇明东滩人工湿地越冬水禽行为观察. 生态学杂志, 2008, 27(6): 940- 945.
[29] 葛振鸣, 王天厚, 施文彧, 周晓. 长江口杭州湾鸻形目鸟类群落季节变化和生境选择. 生态学报, 2006, 26(1): 40- 47.
[30] 袁玉洁, 梁婕, 黄璐, 龙勇, 申升, 彭也茹, 武海鹏, 曾光明. 环境因子对东洞庭湖优势冬季水鸟分布的影响. 应用生态学报, 2013, 24(2): 527- 534.
[31] Germaine S S, Rosenstock S S, Schweinsburg R E, Richardson W S. Relationships among breeding birds, habitat, and residential development in Greater Tucson, Arizona. Ecological Applications, 1998, 8(3): 680- 691.
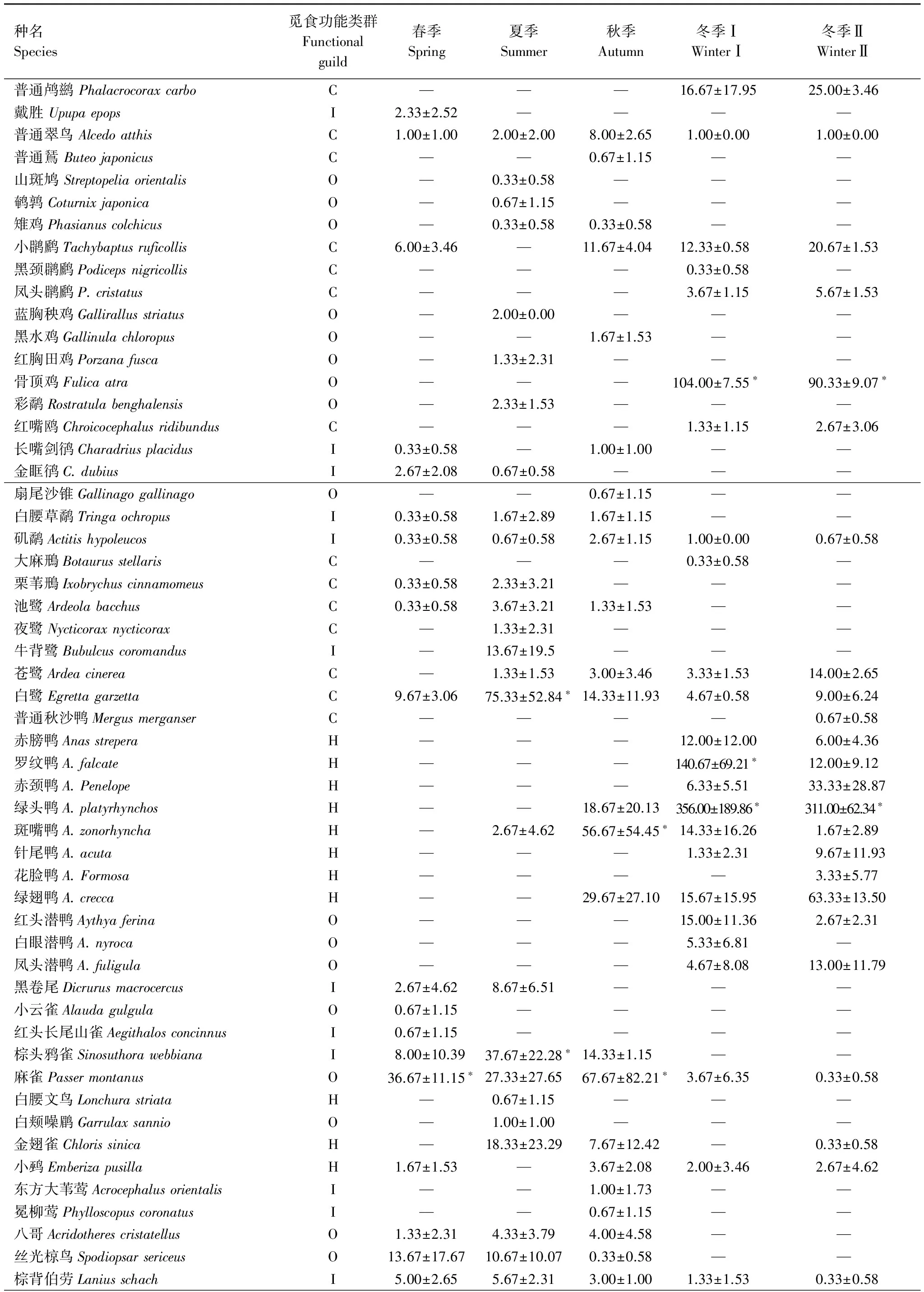
附录1 研究区域不同季节和不同水位时期主要鸟类数量/只
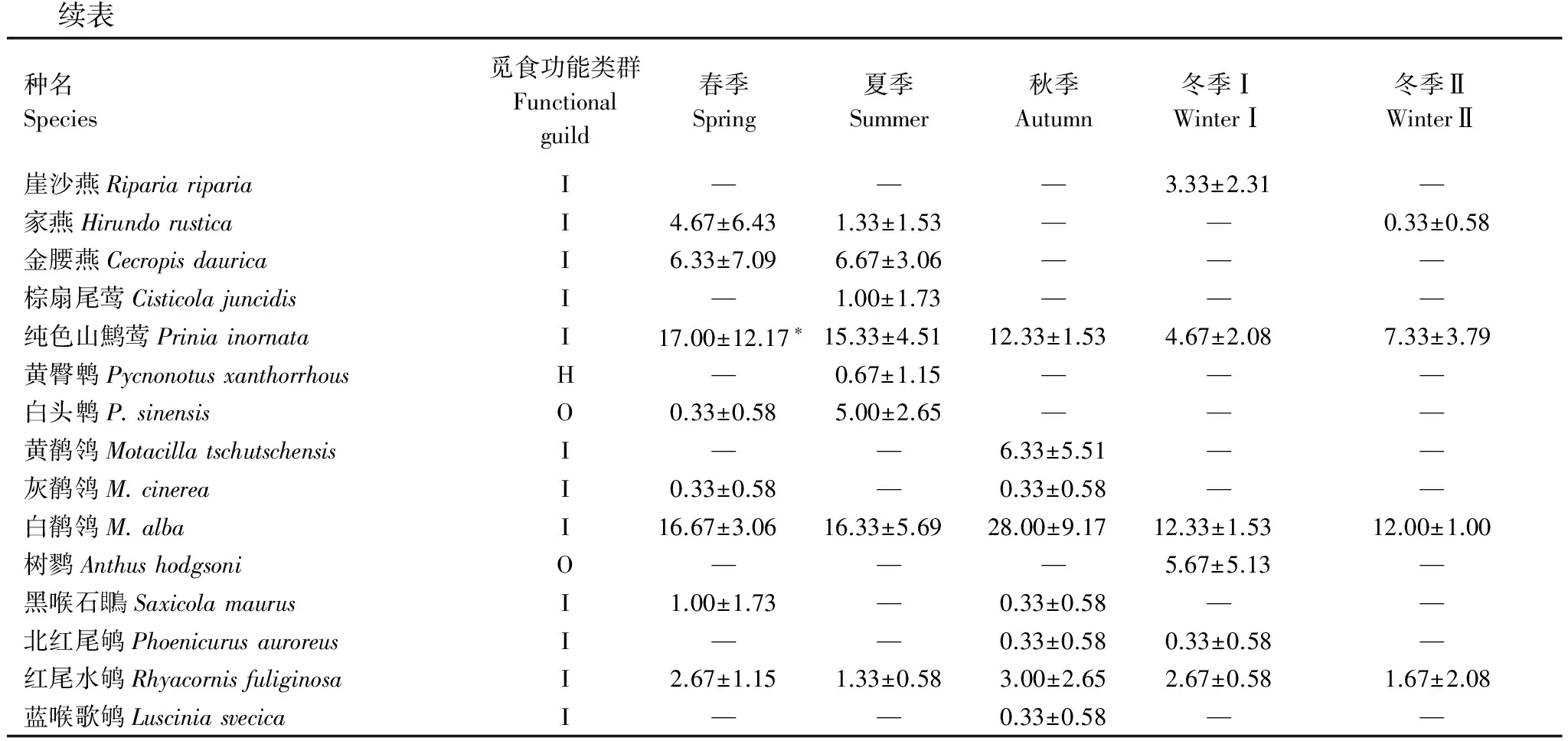
续表种名Species觅食功能类群Functionalguild春季Spring夏季Summer秋季Autumn冬季ⅠWinterⅠ冬季ⅡWinterⅡ崖沙燕RipariaripariaI———3.33±2.31—家燕HirundorusticaI4.67±6.431.33±1.53——0.33±0.58金腰燕CecropisdauricaI6.33±7.096.67±3.06———棕扇尾莺CisticolajuncidisI—1.00±1.73———纯色山鹪莺PriniainornataI17.00±12.17∗15.33±4.5112.33±1.534.67±2.087.33±3.79黄臀鹎PycnonotusxanthorrhousH—0.67±1.15———白头鹎P.sinensisO0.33±0.585.00±2.65———黄鹡鸰MotacillatschutschensisI——6.33±5.51——灰鹡鸰M.cinereaI0.33±0.58—0.33±0.58——白鹡鸰M.albaI16.67±3.0616.33±5.6928.00±9.1712.33±1.5312.00±1.00树鹨AnthushodgsoniO———5.67±5.13—黑喉石即鳥SaxicolamaurusI1.00±1.73—0.33±0.58——北红尾鸲PhoenicurusauroreusI——0.33±0.580.33±0.58—红尾水鸲RhyacornisfuliginosaI2.67±1.151.33±0.583.00±2.652.67±0.581.67±2.08蓝喉歌鸲LusciniasvecicaI——0.33±0.58——
C:肉食性鸟Carnivore; I:食虫鸟Insectivore; O:杂食性鸟Omnivore; H:植食性鸟Herbivore; *:各季节的优势种;—:该季节研究区域内没有观察到该种鸟类
猜你喜欢
学与玩(2022年9期)2022-10-31 02:54:08
落叶果树(2021年6期)2021-02-12 01:29:10
中国水利(2020年14期)2020-08-02 16:19:30
文苑(2020年12期)2020-04-13 00:54:14
小太阳画报(2019年1期)2019-06-11 10:29:48
小学生必读(低年级版)(2017年5期)2017-08-12 03:47:07
体育科技(2016年2期)2016-02-28 17:06:07
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
湖泊科学(2014年4期)2014-02-27 10:30:32
中国工程咨询(2011年11期)2011-02-13 02:35:00
