
不同时间和空间尺度上台湾水青冈群落谱系结构动态变化
2018-03-22 11:33:29吴倩倩梁宗锁刘金亮翁东明张宏伟于明坚
生态学报 2018年4期
吴倩倩,梁宗锁,刘金亮,翁东明,张宏伟,于明坚,胡 广
1 浙江理工大学生命科学学院, 杭州 310018 2 浙江大学生命科学学院, 杭州 310058 3 浙江清凉峰国家级自然保护区管理局, 临安 311300 4 浙江理工大学建筑工程学院, 杭州 310018
群落内现有物种组成是进化过程和生态过程共同作用的结果,群落谱系结构结合了物种的进化历史去反映群落的组成特征,对于了解长期的群落构建、种间关系和群落聚群等过程有着重要作用[1-2]。2000年,Webb首次运用谱系树来分析森林群落的构建机制[3]。2002年,Webb等又进一步系统地阐述了群落谱系结构研究的具体方法,如利用净谱系亲缘关系指数(Net Relatedness Index,NRI)、净最近种间亲缘关系指数(Nearest Taxon Index,NTI)等指数来描述群落的谱系结构格局[4]。群落谱系结构格局的聚集或者发散与环境过滤或者竞争排斥作用等生态学过程密切相关[4- 6],可据此探索调控群落聚群的生态学过程与机制(表1)。在鼎湖山森林中开展的植物群落谱系结构研究发现环境过滤作用是该地区主要的群落构建机制[7]。Whitfeld等人在研究热带森林的谱系结构中发现热带森林的演替符合环境过滤的构建过程[8]。

表1 群落聚群过程与谱系结构的关系
MPD:平均谱系距离,Mean Phylogenetic Distance;MNTD:最近种间平均谱系距离,Mean Nearest Taxon Distance;NRI:净谱系亲缘关系指数,Net Relatedness Index;NTI:净最近种间亲缘关系指数,Nearest Taxon Index
由于植物一些重要的功能特征具有较强的谱系保守性,因此谱系结构在一定程度上也体现了群落物种的功能结构[9-10]。而局部群落的谱系结构又受到生物和非生物过程的影响,从而谱系结构也反映了局域生境对于物种进化历史的选择[4]。现有研究表明群落谱系结构的变化受到很多方面的影响,如尺度、生态特征、环境因子等[11- 14]。Swenson等发现小尺度上植物群落谱系结构趋于发散,反之聚集[15]。Baldeck等发现谱系β多样性与土壤及地形差异有关[16]。因此可以通过研究影响群落谱系结构的调控因素,来进一步深入了解环境变化和尺度依赖性对植物群落的影响。
珍稀植物对生境条件、气候变化较为敏感,具有很大的研究及保护价值。而以往对珍稀植物的研究多集中在繁育、种群等方面的研究[17-18],对于整个群落的研究较少。台湾水青冈(Fagushayatae)属壳斗科(Fagaceae)水青冈属(Fagus)的落叶乔木,为国家Ⅱ级重点保护野生植物[19]。浙江清凉峰国家级自然保护区内保存有一定规模,发育良好的台湾水青冈群落,较为罕见,具有较高的保护和研究价值[20]。然而近年来由于极端天气现象多发,对该地区植被造成较大影响。因此,通过分析台湾水青冈群落的谱系结构可以探寻其发展趋势及濒危机制,对于今后的保护工作具有重要的意义。故而本文以清凉峰1hm2台湾水青冈群落动态监测样地为研究对象,于2006年,2011年,2016年3次对样地内木本植物进行调查,分析群落内物种在不同尺度下的动态谱系结构,旨在了解:(1)10年间台湾水青冈群落的谱系结构变化;(2)在不同尺度下,台湾水青冈群落的谱系结构差异;(3)谱系结构研究在台湾水青冈种群保护策略制定中的作用。
1 研究地区与研究方法
1.1 研究地自然概况
研究区域位于浙江省临安市西部的清凉峰国家级自然保护区(118°52′—119°11′E,30°5′—30°17′N),与安徽省的绩溪、歙县毗邻,是白际山脉北段的一部分。龙塘山地处中亚热带北缘,具有明显的亚热带中山山地季风特征。全年降水量1500—1900mm,并随着海拔高度的变化而变化,在海拔900—1100m范围达最大;同时降水量随季节分配不均匀,夏、秋两季雨量较大,冬、春季节则相对较少。相对湿度在78%—82%之间,夏季较冬季湿度大。温度随海拔高度和季节差异的变化较大,高低海拔间平均气温变化范围为7.8—15.3℃,年积温为2200—4800℃[20- 22]。
1.2 样地设置与调查
2006年在清凉峰保护区龙塘山保护站(118°07.270′E,29°15.098′N)建立以台湾水青冈为建群种,面积为1hm2(100m×100m)的森林动态样地,样地具体描述见[23-24]。对样地内胸径(DBH)≥1cm的木质非藤本植物均进行了物种鉴定、胸径、树高和坐标位置的测量,并挂牌标记。2011年和2016年两次对该动态样地进行复查,对样地内原挂牌纪录物种进行胸径的测定,记录死亡情况,并对新增的DBH≥1cm的木本个体调查记录种名、胸径、坐标并挂牌标记。
1.3 群落谱系结构
物种定名参考《中国在线植物志》(www.eflora.cn)以及《浙江种子植物检索鉴定手册》[25],并根据TPL(the Plant List)网站(www.theplantlist.org)对定名进行规范。采用Shannon-Wiener指数估计物种的α多样性。谱系树构建基于Zanne等发表的谱系数据库,该数据库包含全球30000多种种子植物的谱系数据[26]。通过结合样地内获取的植物物种信息,参考Qian和Jin介绍的“Scenario 3”方法,使用软件S.PhyloMaker 获取各物种谱系位点并构建谱系树[27],相当于BLADJ(branch length adjuster)方法[28]。
研究选择了在谱系结构研究中广泛使用的平均谱系距离(Mean Phylogenetic Distance,MPD)、最近种间平均谱系距离(Mean Nearest Taxon Distance,MNTD)、净谱系亲缘关系指数(Net Relatedness Index,NRI)和净最近种间亲缘关系指数(Nearest Taxon Index,NTI)来代表群落谱系结构[4]。MPD为群落各物种对之间谱系位点距离的平均值,MNTD为物种谱系位点与其最近物种谱系位点距离的平均值[3],这两个指数体现了群落谱系结构的绝对变化[29],可用于同一群落时间动态上的比较。NRI是标准化的MPD,对检验环境过滤更为有效;NTI则是指标准化的MNTD,对检验竞争具有更大的统计能力[1,6,15]。NRI和NTI均可用于比较不同取样强度群落的谱系结构差异。
为探讨不同的空间尺度对谱系多样性的影响,本文分别在10、20和50m 3个空间取样尺度上计算群落谱系结构指数。根据样地内每木的空间坐标在1hm2样地范围中利用移动窗口法随机选取10m×10m,20m×20m,50m×50m的小样方,计算相关面积小样方内的谱系结构指数,各取样尺度随机取样999次。首先通过计算出随机选取的小样方中所有物种对的MPD和MNTD的实际观察值;保持小样方内物种数量及物种个体数不变,从物种库(即整个1hm2样地中所有的物种)中随机抽取物种构建随机零模型并重复999次,从而获得该小样方中物种在随机分布条件下的MPD和MNTD的分布,最后利用随机模拟结果将观察值标准化,从而获得NRI和NTI。具体计算公式为[4,30]:
其中n为群落内物种数,δij为物种i和物种j之间的谱系距离,minδij为物种i与群落内其他物种间的最短谱系距离,MPDs和MNTDs为每个样方中的观察值,MPDr和MNTDr为每个样方中通过随机模型所获得值的平均值。NRI 和NTI如果小于0,表明样方中的物种在谱系结构上是发散的,如果大于0说明样方中的物种在谱系结构上是聚集的,如果等于0则说明谱系随机。
所有计算使用R 3.2.3软件完成。使用R软件中的Vegan包[31]计算物种多样性,用Picante包[32]计算相关谱系结构指数;采用t检验进行显著性检验。
2 结果与分析
2.1 群落物种组成动态变化
总体来看,台湾水青冈群落物种多样性随时间增加而减少(表2)。台湾水青冈种群龄级结构呈反“J”形,为增长型(图1),在2006年到2016年的10年森林动态变化过程中,Ⅰ、Ⅱ龄级数量减少,其他龄级数量较为稳定。2011年,台湾水青冈种群死亡率为4.24%,补员率为4%;2016年,台湾水青冈种群死亡率为6.95%,补员率为1.9%。

表2 2006—2016年台湾水青冈群落物种组成动态变化
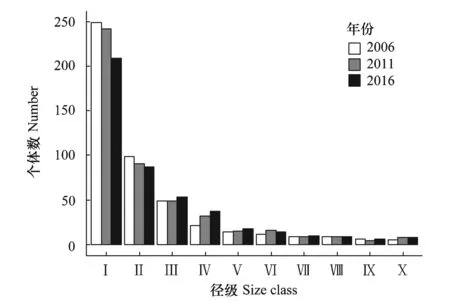
图1 台湾水青冈种群径级结构Fig.1 Diameter structure of Fagus hayatae population Ⅰ级,1cm≤DBH<5cm;Ⅱ级,5cm≤DBH<10cm;Ⅲ级,10cm≤DBH<15cm;Ⅳ级,15cm≤DBH<20cm;Ⅴ级,20cm≤DBH<25cm;Ⅵ级,25cm≤DBH<30cm;Ⅶ级,30cm≤DBH<35cm;Ⅷ级,35cm≤DBH<40cm;Ⅸ级,40cm≤DBH<45cm;Ⅹ级,DBH≥45cm
2.2 群落谱系结构动态变化
群落MPD和MNTD的动态变化趋势相反。随着时间的增长,MPD值均呈下降趋势,而MNTD呈现上升趋势(图2)。
NRI和NTI的动态变化趋势要弱于MPD和MNTD。10m尺度下,NRI和NTI随时间变化没有表现出显著差异;而20m和50m尺度下, NRI和NTI均随时间增加有显著增加(图3)。
2.3 空间尺度与谱系结构动态的关系
随着取样空间尺度的增大,对应年份的群落MPD增加,而MNTD值减小。3个空间尺度下,群落内的NRI和NTI均显著大于0,谱系结构表现聚集格局,且随着空间尺度增大,NRI和NTI数值显著增大(图3),聚集格局愈加明显。
空间尺度的变化对谱系结构的时间动态变化有明显的影响。随着取样空间尺度的增加,MPD和MNTD的动态变化趋势逐渐增强(图2)。在10m尺度,2016年MPD值与其他年份MPD存在显著差异,2006年MNTD与其他年份存在显著差异;50m尺度下,2006年、2011年、2016年之间MPD和MNTD均有显著差异。NRI和NTI的动态变化趋势受空间尺度的影响和MPD和MNTD类似,大尺度下的变化趋势要强于小尺度(图3)。
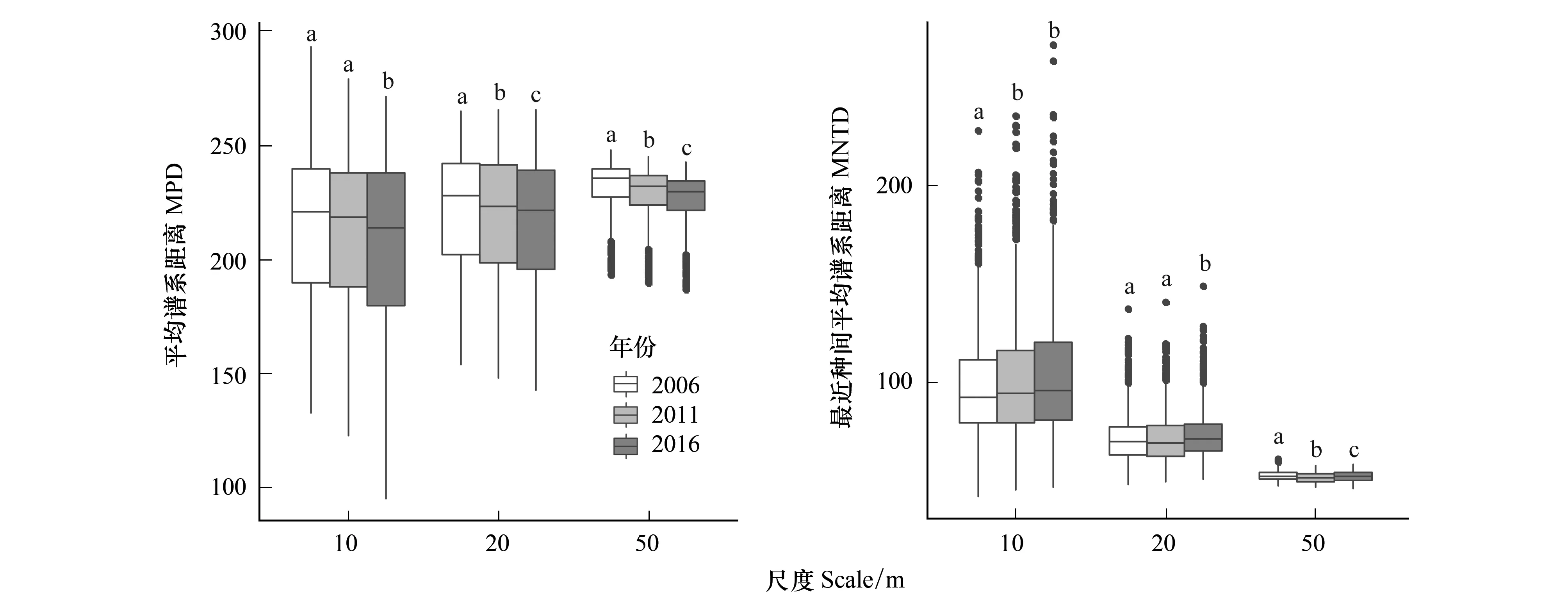
图2 样地群落内所有物种在不同尺度下的平均谱系距离(MPD)和最近种间平均谱系距离(MNTD)Fig.2 The mean phylogenetic distance (MPD) values and the mean nearest taxon distance (MNTD) values for different scales同一尺度下,字母不同表示数据间存在显著差异,相同表示没有显著差异

图3 样地群落内所有物种在不同尺度下的净谱系亲缘关系指数(NRI)和净最近种间亲缘关系指数(NTI)Fig.3 The net relatedness index (NRI) values and the nearest taxon index (NTI) values for different scales同一尺度下,字母不同表示数据间存在显著差异,相同表示没有显著差异
3 讨论
3.1 不同时间尺度下群落谱系结构的变化
在时间尺度上的群落谱系结构动态反映了群落的发展趋势。在2006年到2016年的10年森林动态变化过程中,台湾水青冈群落MPD减小,可知群落整体谱系距离减小。由于台湾水青冈群落补员率较低,该群落内物种丰富度随时间增加而降低(图1),结合MPD数据应该是群落中谱系亲缘关系较远、生境需求较为特殊、环境耐受性较差的物种大量丧失,这就为具有较高环境耐受性,谱系关系接近的植物的更新提供了大量空间。已有研究发现,温度和降水是影响亚热带地区森林群落物种多样性和谱系多样性的主导因子[13]。我们猜测当地生境质量下降、抗干扰性较弱的物种无法生存,进而导致台湾水青冈群落谱系关系在近10年表现出聚集格局,主要是由气候变化引起的近几年极端恶劣天气的频发,如近年频发的冰雪灾害[33]和台风影响[34]所导致的。NRI随时间的不断增加(谱系多样性下降)和物种丰富度的降低也说明了生境过滤作用是调控群落聚群的主导过程(表1),且这种过程在10年的森林动态变化中不断增强。对于古田山和鼎湖山大型森林动态样地的研究也表明生境过滤作用对当地生物多样性和森林更新过程具有重要作用[7,35]。如果没有采取一定措施,群落的物种和谱系多样性在未来可能会持续降低,影响整个群落的稳定和抗干扰能力,对台湾水青冈种群造成持续的不良影响。
MNTD在近10年间持续增大,说明尽管群落的整体谱系结构呈现不断聚集的过程,但是各物种的近缘种也逐渐被替代成谱系关系较远的物种,呈现发散的格局。可能是由于生境质量的下降导致小尺度下近缘种对同种资源的争夺加强,种间竞争作用增强。清凉峰的早期研究指出该样地的8个优势树种中有6个物种的聚集程度随着树木的生长而减小,并且聚集程度的减幅在5m的取样空间尺度内效果最强[36]。NTI的变化趋势随时间逐渐增大,说明了生境过滤作用仍是调控群落聚群的主导过程,但由于在小尺度下近缘种之间竞争的强化,近10年近缘种谱系结构动态变化趋势要弱于群落整体谱系结构的变化,相对处于比较稳定的水平。
3.2 不同空间尺度群落谱系结构的变化
空间尺度是影响谱系结构变化的重要因子[37-38]。一般来说,随着空间尺度的增加,谱系结构有逐渐聚集的趋势[15]。本研究中,随着研究尺度的增加,群落的整体谱系结构和近缘种谱系结构均表现为聚集(图3),且聚集程度持续增加。在古田山常绿阔叶林群落不同空间尺度(取样半径为5、25、50、75、100m)的谱系结构研究中,也均发现显著的谱系聚集(先增加后下降),并且在5—50m尺度下聚集程度增加[2],这一现象主要是由环境过滤过程所调控的,与本研究结果一致。可见生境过滤作用在台湾水青冈群落构建和发展中起主导作用,空间尺度越大,生境过滤作用越强。同时我们发现谱系结构的动态变化及其生态过程的效应强度也受到研究尺度的影响。在50m尺度下,群落的整体谱系结构和近缘种谱系结构均随时间增长而愈加紧密,环境过滤作用也在不断增强。而在小尺度下,则没有明显表现出这种时间序列上的变化趋势(图3),可能是由于小尺度下的随机过程,种间竞争作用以及临近母体的幼树表现出的Janzen-Connell效应所导致的[39]。
3.3 谱系结构动态对台湾水青冈群落保护的意义
结合台湾水青冈群落谱系结构的动态变化和空间尺度效应,发现环境过滤作用是影响该植物群落未来发展的主导效应。近年来全球气候变化导致的区域性极端天气频发,改变了植物群落的组成和结构[40-41],据此导致的强烈的环境过滤作用使得植物群落的功能和谱系结构趋向单一化。对于具有生境专一性的珍稀濒危植物来说,其负面效应更是影响巨大。谱系和功能的单一导致群落稳定性和生态系统功能下降,一旦环境条件不能满足该类植物的需要,且珍稀植物大多种群规模较小,小种群效应极有可能导致该物种的灭绝。同时群落谱系结构的紧密化(图2),也导致了小尺度上种间竞争效应的增强(表1),严重影响到台湾水青冈的群落补员率,进而影响其种群更新。环境过滤作用降低了台湾水青冈群落的整体稳定性,而小尺度上近缘种的竞争则阻碍了其种群的更新和发展,因此台湾水青冈群落的发展前景不容乐观,保护形势十分艰巨。
4 结论
通过研究发现,在2006—2016年间(1)台湾水青冈群落总体谱系结构表现出不断聚集的趋势,而近缘种的谱系关系则不断疏远;(2)随着空间尺度的增加,台湾水青冈群落的谱系结构愈加聚集,且谱系结构动态变化的聚集程度也随空间尺度的增加而增强。上述结论说明清凉峰国家级自然保护区的台湾水青冈群落的谱系结构在目前的气候背景下逐步趋向单一化,发展前景不容乐观。因此根据相关研究结果,针对该群落可以提出一些保护建议:(1)由于常绿树种具有更强的环境耐受性,在全球变暖和环境恶化的背景下具有优势,有很大的可能挤占落叶树种台湾水青冈的生存空间,因此需要进行一定的人为调控,维持该群落内落叶树种的优势;(2)进一步研究分析台湾水青冈幼树和幼苗高死亡率的直接原因,采取有效的针对性措施提高群落补员率。
致谢:美国亚利桑那州立大学Maxwell Wilson博士帮助写作,特此致谢。
[1] 牛红玉, 王峥峰, 练琚愉, 叶万辉, 沈浩. 群落构建研究的新进展: 进化和生态相结合的群落谱系结构研究. 生物多样性, 2011, 19(3): 275- 283.
[2] 黄建雄, 郑凤英, 米湘成. 不同尺度上环境因子对常绿阔叶林群落的谱系结构的影响. 植物生态学报, 2010, 34(3): 309- 315.
[3] Webb C O. Exploring the phylogenetic structure of ecological communities: an example for rain forest trees. The American Naturalist, 2000, 156(2): 145- 155.
[4] Webb C O, Ackerly D D, McPeek M A, Donoghue M J. Phylogenies and community ecology. Annual Review of Ecology and Systematics, 2002, 33(1): 475- 505.
[5] 卜文圣, 许涵, 臧润国, 丁易, 张俊艳, 林明献. 不同采伐干扰方式对热带山地雨林谱系结构的影响. 林业科学, 2014, 50(4): 15- 21.
[6] 柴永福, 岳明. 植物群落构建机制研究进展. 生态学报, 2016, 36(15): 4557- 4572.
[7] Huang J X, Zhang J, Shen Y, Lian J Y, Cao H L, Ye W H, Wu L F, Yue B. Different relationships between temporal phylogenetic turnover and phylogenetic similarity and in two forests were detected by a new null model. PLoS One, 2014, 9(4): e95703.
[8] Whitfeld T J S, Kress W J, Erickson D L, Weiblen G D. Change in community phylogenetic structure during tropical forest succession: evidence from New Guinea. Ecography, 2012, 35(9): 821- 830.
[9] Prinzing A, Reiffers R, Braakhekke W G, Hennekens S M, Tackenberg O, Ozinga W A, Schaminée J H J, van Groenendael J M. Less lineages-more trait variation: phylogenetically clustered plant communities are functionally more diverse. Ecology Letters, 2008, 11(8): 809- 819.
[10] 周道玮. 植物功能生态学研究进展. 生态学报, 2009, 29(10): 5644- 5655.
[11] Pausas J G, Verdú M. The jungle of methods for evaluating phenotypic and phylogenetic structure of communities. BioScience, 2010, 60(8): 614- 625.
[12] 刘巍, 曹伟. 长白山植物群落谱系结构及环境因子对其的影响. 干旱区资源与环境, 2013, 27(5): 63- 68.
[13] Liu J L, Qian H, Jin Y, Wu C P, Chen J H, Yu S Q, Wei X L, Jin X F, Liu J J, Yu M J. Disentangling the drivers of taxonomic and phylogenetic beta diversities in disturbed and undisturbed subtropical forests. Scientific Reports, 2016, 6: 35926.
[14] 宋凯, 米湘成, 贾琪, 任海保, Bebber D, 马克平. 不同程度人为干扰对古田山森林群落谱系结构的影响. 生物多样性, 2011, 19(2): 190- 196.
[15] Swenson N G, Enquist B J, Thompson J, Zimmerman J K. The influence of spatial and size scale on phylogenetic relatedness in tropical forest communities. Ecology, 2007, 88(7): 1770- 1780.
[16] Baldeck C A, Kembel S W, Harms K E, Yavitt J B, John R, Turner B L, Madawala S, Gunatilleke N, Gunatilleke S, Bunyavejchewin S, Kiratiprayoon S, Yaacob A, Supardi M N N, Valencia R, Navarrete H, Davies S J, Chuyong G B, Kenfack D, Thomas D W, Dalling J W. Phylogenetic turnover along local environmental gradients in tropical forest communities. Oecologia, 2016, 182(2): 547- 557.
[17] 袁春明, 孟广涛, 方向京, 柴勇, 李贵祥, 蔺汝涛, 王骞, 李品荣. 珍稀濒危植物长蕊木兰种群的年龄结构与空间分布. 生态学报, 2012, 32(12): 3866- 3872.
[18] 乔琦, 秦新生, 邢福武, 陈红锋, 刘东明. 珍稀植物伯乐树一年生更新幼苗的死亡原因和保育策略. 生态学报, 2011, 31(16): 4709- 4716.
[19] 陈子英, 谢长富, 毛俊杰, 赖玉菁, 林世宗, 胡哲明, 徐堉峰, 杨正钏, 孔祥璿, 林哲荣, 陈品邑, 邱宗仪, 巫智斌. 冰河孑遗的夏绿林――台湾水青冈. 台北: 农业委员会林务局, 2011: 1- 26.
[20] 张方钢. 浙江清凉峰台湾水青冈林的群落学特征. 浙江大学学报: 农业与生命科学版, 2001, 27(4): 403- 406.
[21] 宋朝枢. 浙江清凉峰自然保护区科学考察集. 北京: 中国林业出版社, 1997: 86- 115.
[22] 李明华. 清凉峰自然保护区志. 长春: 吉林人民出版社, 2006: 1- 67.
[23] 郭瑞, 翁东明, 金毅, 张宏伟, 丁文勇, 程樟峰, 于明坚. 浙江清凉峰台湾水青冈种群2006—2011年更新动态及其与生境的关系. 广西植物, 2014, 34(4): 478- 483.
[24] 翁东明, 张磊, 陈晓栋, 沈国春, 张宏伟, 张方钢, 于明坚. 清凉峰自然保护区台湾水青冈群落物种多样性研究. 浙江林业科技, 2009, 29(4): 1- 6.
[25] 郑朝宗. 浙江种子植物检索鉴定手册. 杭州: 浙江科学技术出版社, 2005: 1- 558.
[26] Zanne A E, Tank D C, Cornwell W K, Eastman J M, Smith S A, FitzJohn R G, McGlinn D J, O′Meara B C, Moles A T, Reich P B, Royer D L, Soltis D E, Stevens P F, Westoby M, Wright I J, Aarssen L, Bertin R I, Calaminus A, Govaerts R, Hemmings F, Leishman M R, Oleksyn J, Soltis P S, Swenson N G, Warman L, Beaulieu J M. Three keys to the radiation of angiosperms into freezing environments. Nature, 2014, 506(7486): 89- 92.
[27] Qian H, Jin Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. Journal of Plant Ecology, 2016, 9(2): 233- 239.
[28] Webb C O, Ackerly D D, Kembel S W. Phylocom: software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics, 2008, 24(18): 2098- 2100.
[29] Stevens R D, Gavilanez M M. Dimensionality of community structure: phylogenetic, morphological and functional perspectives along biodiversity and environmental gradients. Ecography, 2015, 38(9): 861- 875.
[30] Webb C O, Donoghue M J. Phylomatic: tree assembly for applied phylogenetics. Molecular Ecology Notes, 2005, 5(1): 181- 183.
[31] Oksanen J, Blancher F G, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin P R, O′Hara R B, Simpson G L, Solymos P, Stevens M H H, Szoecs E, Wagner H. Vegan: community ecology package. R package version 2.4- 3. [2016-05-05] http://cran.r-project.org, https://github.com/vegandevs/vegan.
[32] Kembel S W, Cowan P D, Helmus M R, Cornwell W K, Morlon H, Ackerly D D, Blomberg S P, Webb C O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics, 2010, 26(11): 1463- 1464.
[33] 金毅, 陈建华, 米湘成, 任海保, 马克平, 于明坚. 古田山24ha森林动态监测样地常绿阔叶林群落结构和组成动态: 探讨2008年冰雪灾害的影响. 生物多样性, 2015, 23(5): 610- 618.
[34] 王樟华, 王希华, 沈国春. 台风干扰对天童常绿阔叶林凋落物量的影响. 华东师范大学学报: 自然科学版, 2014, (1): 79- 89.
[35] 田锴, 陈磊, 米湘成, 马克平, 陈建华. 亚热带常绿阔叶林木本植物幼苗分布格局及其对生境过滤的响应. 科学通报, 2013, 58(34): 3561- 3569.
[36] 丁文勇, 翁东明, 金毅, 张宏伟, 程樟峰, 郭瑞, 于明坚. 清凉峰自然保护区台湾水青冈群落优势种群密度制约效应分析. 浙江大学学报: 理学版, 2014, 41(5): 583- 592.
[37] Kembel S W, Hubbell S P. The phylogenetic structure of a neotropical forest tree community. Ecology, 2006, 87(S7): S86-S99.
[38] Cavender-Bares J, Keen A, B M. Phylogenetic structure of Floridian plant communities depends on taxonomic and spatial scale. Ecology, 2006, 87(S7): S109-S122.
[39] Petermann J S, Fergus A J F, Turnbull L A, Schmid B. Janzen-Connell effects are widespread and strong enough to maintain diversity in grasslands. Ecology, 2008, 89(9): 2399- 2406.
[40] Walther G R. Community and ecosystem responses to recent climate change. Philosophical Transactions of the Royal Society B: Biological Sciences, 2010, 365(1549): 2019- 2024.
[41] Walck J L, Hidayati S N, Dixon K W, Thompson K, Poschlod P. Climate change and plant regeneration from seed. Global Change Biology, 2011, 17(6): 2145- 2161.
猜你喜欢
艺术品鉴(2022年16期)2022-07-09 03:31:08
昆明医科大学学报(2022年2期)2022-03-29 00:51:58
现代园艺(2021年23期)2021-12-01 07:47:44
食品安全导刊(2021年20期)2021-08-30 06:40:50
河池学院学报(2021年1期)2021-07-10 05:14:16
景德镇陶瓷(2021年1期)2021-03-24 05:47:54
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
东方考古(2017年0期)2017-07-11 01:37:50
水生生物学报(2015年1期)2015-02-28 16:01:05
