
荒漠区植物雾冰藜光合特性对混合盐碱胁迫的响应
2018-03-22 11:33赵文智
生态学报 2018年4期
李 辛, 赵文智,2,*
1 甘肃农业大学林学院, 兰州 730070 2 中国科学院西北生态环境资源研究院,中国生态系统研究网络临泽内陆河流域研究站, 兰州 730000
光合作用是所有植物、藻类和某些细菌同化作用的基础,也是生物圈物质循环最重要的环节[1],在光能的吸收、固定、分配与转化中发挥关键作用[2]。光合作用对逆境胁迫伤害最为敏感,其中盐碱胁迫是影响光合作用的一个重要外在因素[3]。目前,土壤盐碱化是全球面临的主要生态环境问题之一,全世界可利用的土地面积约为1.5×109hm2,其中约有23%的土地存在盐化,另有37% 的土地存在碱化[4]。我国约有2×107hm2的盐荒地和6.67×106hm2的盐碱化土壤[5],其盐分含量一般为0.07%—1.3%,pH范围一般为6.9—10.8,盐碱化程度和盐碱化面积有逐年增大的趋势[6]。由于盐碱地土壤中的盐分种类、数量、比例十分复杂[7],盐碱化土地不同地块或同一地块的不同区域上盐分含量和pH的差异均会影响植物的正常生长和种群分布,因此盐碱混合胁迫是植物在盐碱地上生存所面临的主要问题[8]。近年来许多学者研究了盐碱胁迫对苜蓿[9]、向日葵[10]、杨树[11]、棉花[12]、绣线菊[13]、燕麦[14]、葡萄[15]、虎尾草和星星草[16]等植物生长、生理变化以及光合作用的影响,证明盐碱胁迫对植物的致害作用很强。但是已有研究多以人工栽培的植物品种为研究对象探讨盐碱胁迫的影响,以盐碱地原生植物为研究对象探讨其盐碱生境适应性的研究报道极少。
一年生植物是全球荒漠植物区系的重要组成部分,丰富度很高[17],在各种类型的荒漠生态系统中具有重要的生态功能[18]。一年生植物作为先锋植物或建群种植物,广泛分布于荒漠区土质、沙质、砾质、石质及盐碱土等生境,在长期进化和自然选择的过程中很好的适应了荒漠地区的生物和非生物环境因素,形成了一整套完善的生存策略来应对荒漠生境的随机性和不确定性[19]。基于一年生植物独有的生物学特性,使其成为许多植物生态学关键问题研究的最佳试验材料。
雾冰藜(Bassiadasyphylla)为藜科雾冰藜属一年生草本植物,是半固定或固定沙丘、平坦沙地以及中度盐碱地的先锋植物之一[20],经常散生或群生于我国北方草原、半荒漠和荒漠地区的土质、沙质或沙砾质盐碱土壤上。目前,关于雾冰藜的研究仅见极少量群落结构[21]、空间分布格局分析[22]及生物指示作用[23]的文献报道。但是雾冰藜作为盐碱地先锋植物的生长环境适应性研究尚未开展。本研究以沙培雾冰藜幼苗为研究对象,将两种碱性盐Na2CO3和NaHCO3及两种中性盐NaCl和Na2SO4按不同比例混合,模拟出30种盐度和碱度各不相同的盐碱条件对其进行胁迫处理,探讨其光合特性对盐碱混合胁迫的响应机理,以期从光合生理角度阐明雾冰藜盐碱生境的适应机理,进而为开展盐碱地治理及固沙植被的恢复与重建积累科学数据。
1 材料与方法
1.1 材料培养
供试雾冰藜种子于2015年10月采集于中国生态系统研究网络临泽内陆河流域研究站附近盐碱地,播种于直径和深度均为30 cm盛有洗净细河沙和蛭石的塑料花盆内,河沙和蛭石按照体积比2∶1配制,每盆种植种子20粒。花盆置于室外,人工遮雨,利用自然条件培养雾冰藜苗。出苗后每周用Hoagland营养液透灌1次,每日用称质量法确定每盆的失水量,用蒸馏水补充,至胁迫试验开始时间苗至长势均匀一致的5株。
1.2 盐碱混合胁迫模拟
依据当前河西走廊盐碱地的盐分组成和土壤pH的分布范围[24- 25],将NaCl、Na2SO4两种中性盐和Na2CO3、NaHCO3两种碱性盐按照不同比例混合,按照碱性盐配制比例逐步增大的顺序设计A、B、C、D、E、F 6个不同的处理组(表1),每个处理组内设50、100、150、200、250 mmol/L 5个浓度梯度,总计30个不同盐浓度和pH的处理,以盐浓度为0 mmol/L的双蒸水为对照(CK)。
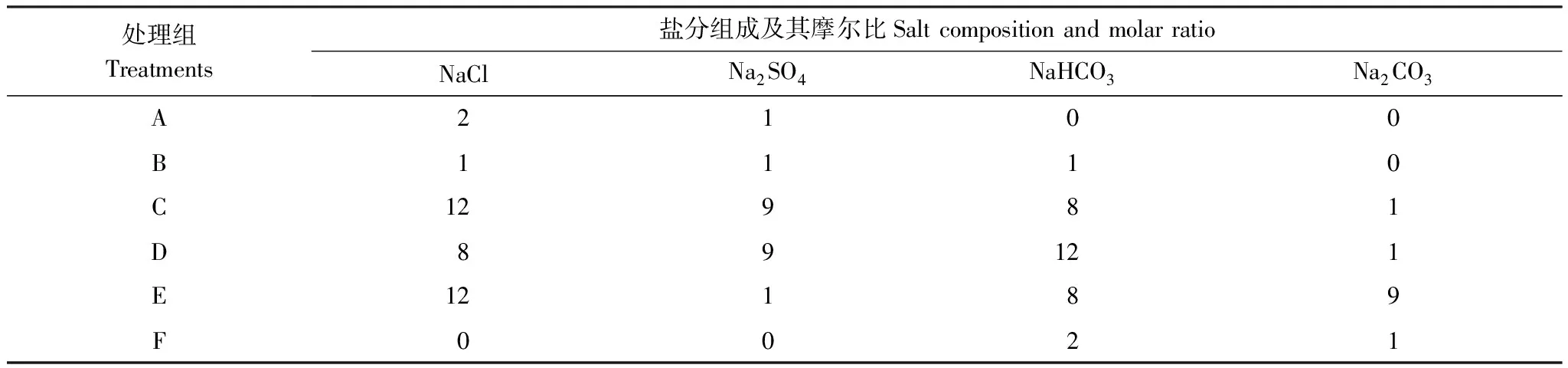
表1 不同处理组的盐分组成及其配比
A、B、C、D、E、F为不同碱性盐配制比例的处理
1.3 盐碱胁迫处理
雾冰藜种子于2016年4月10日播种,苗龄4周时,选取长势均匀一致的雾冰藜苗93盆随机分成31组。其中1组为对照,其余30组为不同程度的盐碱胁迫处理,按照5个浓度梯度依次标为A1、A2、… F4、F5,每组3盆为3次重复,处理于17:00—20:00时进行。以含有相应浓度混合盐碱组合的配比液为处理液,每盆500 mL分3次透灌花盆,对照分3次浇灌等量双蒸水。从次日起用称质量法测定每盆失水量,并用蒸馏水补充。为防止盆内盐分随水分流失,每个盆下放置托盘,将流出的溶液重新倒回盆内,使植株完全处于胁迫状态。在胁迫7 d 后进行光合色素含量、气体交换参数及叶绿素荧光参数的测定。
1.4 指标测定
1.4.1 光合色素含量的测定
叶片的叶绿素 (Chl)含量和类胡萝卜素 (Caro)含量的测定参照李合生[26]的方法。
1.4.2 光合气体交换参数测定
选择胁迫7 d后天气晴朗的一天9: 00—11: 00时段进行光合参数测定。使用 LI- 6400 便携式光合仪(LI-Cor,Inc,美国) 测定雾冰藜幼苗在光强 1000 μmol m-2s-1下的净光合速率(Pn)、气孔导度(Gs)、胞间CO2摩尔分数(Ci)、蒸腾速率(Tr)等指标。雾冰藜水分利用效率(WUE)以植株光合速率与蒸腾速率的比值表示,即WUE=Pn/Tr,式中Pn和Tr分别为同一植株的净光合速率和蒸腾速率的测定值。测定过程中的环境状况为空气温度(22.45±2.31)℃,相对湿度(57.13±2.45)%,空气中CO2浓度(379.51±13.25) μmol/mol。对照和每个处理各测定5次。
1.4.3 叶绿素荧光参数测定
1.5 数据处理
数据分析采用SPSS 19.0软件完成,用Duncan显著性检验方法比较不同处理间的差异性,并进行多元回归分析,利用Microsoft Excel 2010软件做图。
2 结果与分析
2.1 盐碱胁迫处理溶液的盐度和pH
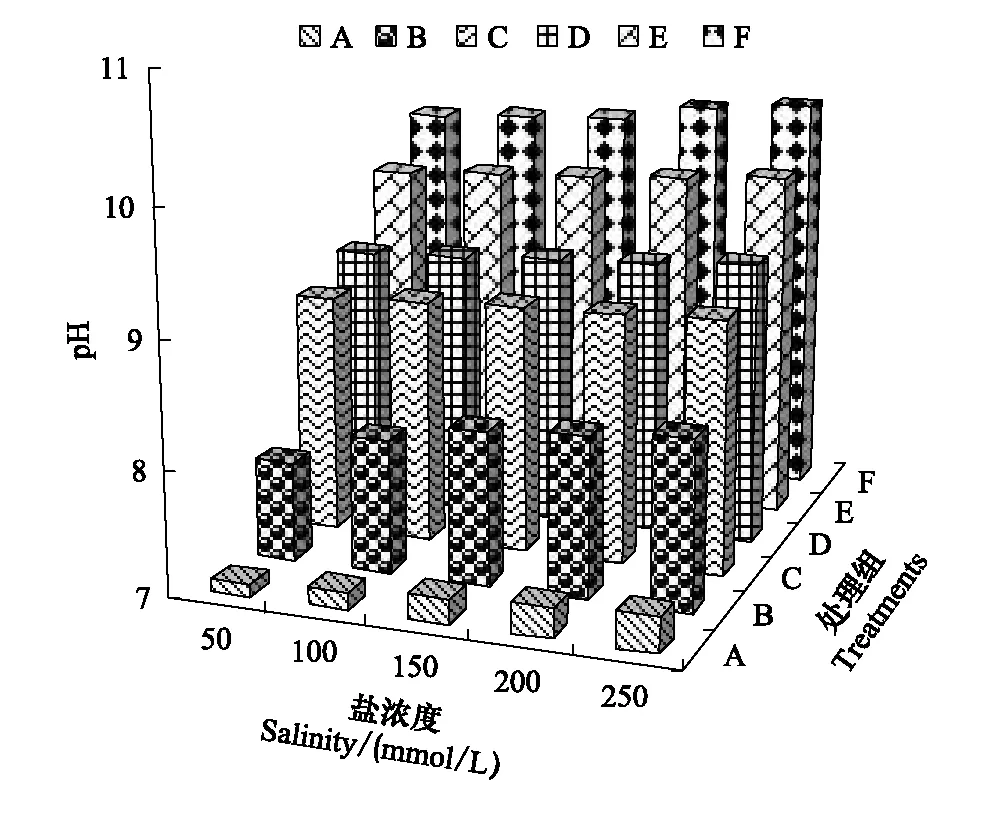
图1 各处理的盐浓度和pH值Fig.1 Total salt concentration and pH of various treatmentsA、B、C、D、E、F为不同碱性盐配制比例的处理

2.2 雾冰藜光合色素含量对盐碱胁迫的响应
从图2可以看出,雾冰藜各盐碱胁迫处理的叶绿素和类胡箩卜素含量均低于双蒸水(CK)处理,即在溶液盐浓度为0 mmol/L、pH 7.00时的光合色素含量最高,叶绿素a(Chla)含量为(240.25±5.75) mg/g,叶绿素b(Chlb)含量为(155.91±5.22) mg/g,类胡萝卜素(Caro)含量为(42.90±5.10) mg/g,叶绿素总量(Chla+Chlb)为(396.16±10.97) mg/g。在同一处理组中,随着溶液盐浓度的不断增大,雾冰藜叶绿素和类胡箩卜素的含量均呈现下降趋势,且在高盐浓度下的下降幅度明显大于低盐浓度。在同一盐浓度下,随着溶液pH的升高,即随碱性盐比例的增大,叶绿素a、叶绿素b和叶绿素总量先增大后减小,类胡箩卜素含量不断降低。叶绿素a、叶绿素b、类胡箩卜素和叶绿素总量均在F5处理下出现最小值,分别比对照降低51.71%、49.53%、61.47%、50.85%。Chla/Chlb的值随着盐浓度和pH的不断增大其变化趋势不同,B1、B2、B4、C1、C2处理下Chla/Chlb的值均高于对照处理,D、E、F组的比值在所有处理中均低于对照;总体而言,A组和E组呈“W”型变化,B、D、F组则先下降后升高再下降,C组呈现先升高后降低的趋势;就变化幅度而言,A组和B组的相对较小。
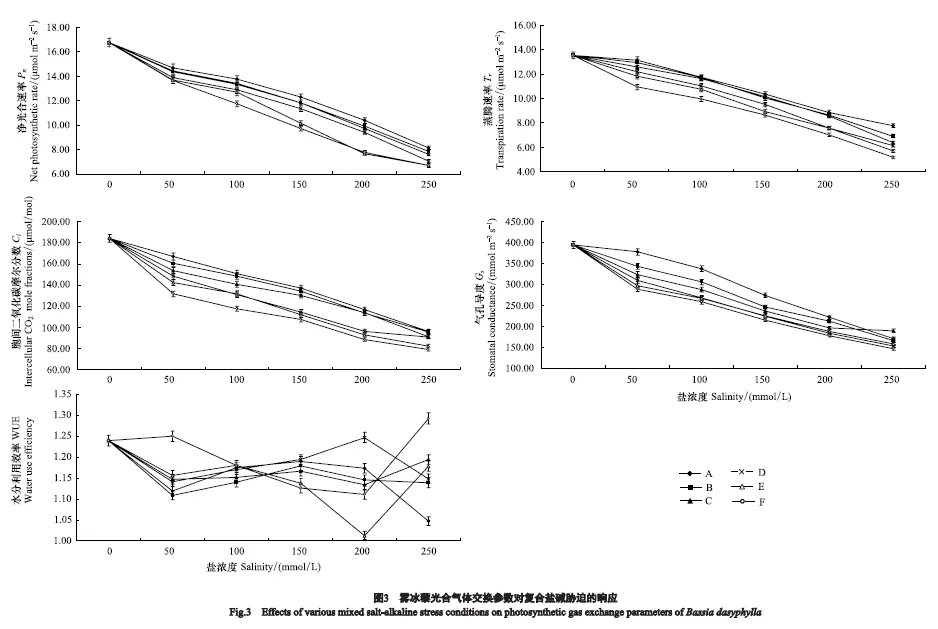
2.3 雾冰藜光合气体交换参数对盐碱胁迫的响应
各胁迫处理对雾冰藜光合气体交换参数的影响如图3所示。雾冰藜各盐碱胁迫处理的净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2摩尔分数(Ci)、气孔导度(Gs)均低于对照处理,即在溶液盐浓度为0 mmol/L、pH 7.00时上述各个参数的值最高,分别为Pn(16.79±0.93) μmol m-2s-1,Tr(13.54±0.42) μmol m-2s-1,Ci(184.06±6.77) μmol m-2s-1,Gs(394.22±11.84) mmol m-2s-1。在同一处理组中,随着溶液盐浓度的不断增大,雾冰藜的Pn、Tr、Ci、Gs均呈现下降趋势,且在高盐浓度时的下降幅度明显大于低盐浓度。在同一盐浓度下,随着溶液pH的升高,即随碱性盐比例的增大,雾冰藜的Pn、Tr、Ci、Gs也不断降低,且均在F5处理下出现最小值,分别比对照降低60.10%、61.74%、57.05%和62.96%。水分利用效率(WUE) 随着溶液盐浓度和pH的不断增大其变化趋势不同,在溶液盐浓度最低50 mmol/L和最高250 mmol/L的处理中,随着溶液pH的不断增大,WUE 也呈现增大的趋势,并在溶液pH最高的F组出现最大值;就其变化趋势而言,A、B、D组呈现先下降后升高再下降的变化趋势,C、E组呈“W”型变化,F组则呈先升高后降低再升高的趋势。
2.4 雾冰藜叶绿素荧光参数对盐碱胁迫的响应
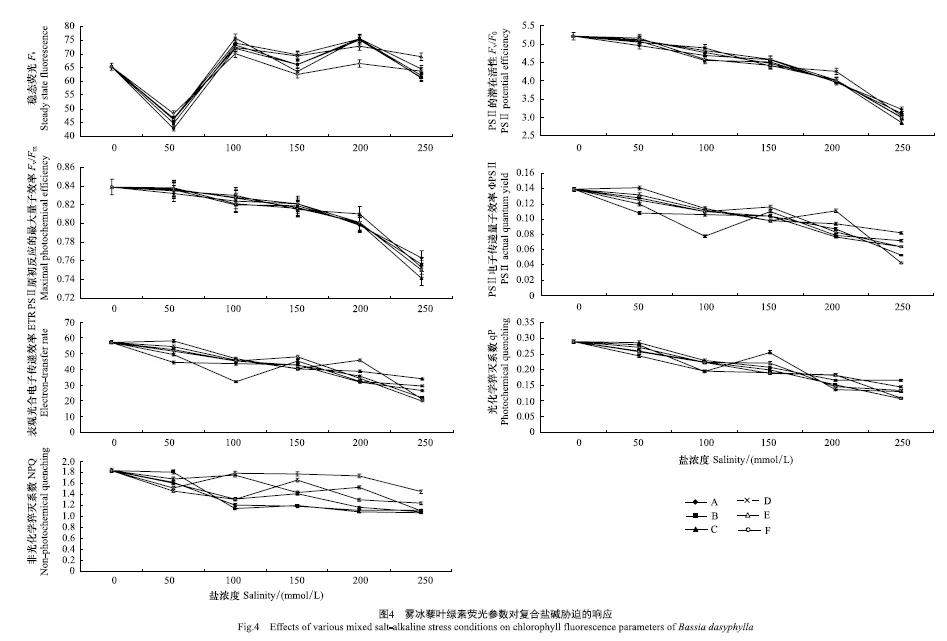
各胁迫处理对雾冰藜叶绿素荧光参数的影响如图4所示。各盐碱胁迫处理组的稳态荧光(Fs)在溶液盐浓度为0—250 mmol/L的范围内先降低,后呈现“M”型变化趋势,各处理组均在盐浓度为50 mmol/L时出现最小值。PSⅡ的潜在活性(Fv/F0)和PSⅡ 原初反应的最大量子效率(Fv/Fm)具有相同的变化趋势,即在溶液盐浓度为0 mmol/L、pH 7.00时(CK)有最大值(5.22±0.12)和(0.84±0.01),并随着处理溶液盐浓度的升高而逐渐降低。PSⅡ电子传递量子效率(ΦPSII)、表观光合电子传递效率(ETR)和光化学猝灭系数(qP)的变化趋势基本一致,均在盐浓度为0 mmol/L、pH 7.00时(CK)有最大值,分别为(0.14±0.05)、 (57.28±2.45)和(0.29±0.04),并均随盐浓度的升高而下降,而在相同盐浓度不同pH的条件下变化趋势没有明显的规律性。非光化学猝灭系数(NPQ)在盐浓度为0 mmol/L、pH 7.00时(CK)有最大值(1.84±0.09),并随着处理溶液盐浓度的升高而基本呈现先降低后升高再降低的变化趋势;当溶液盐浓度高于50 mmol/L时,在相同盐浓度处理组下的NPQ随处理溶液pH的升高而升高。
2.5 盐分离子组成与雾冰藜光合特征各项指标间的相关性
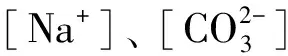

表2 盐碱离子组成与雾冰藜光合指标间的相关系数
*表示P< 0.05,**表示P< 0.01
3 讨论
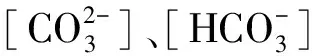
通常情况下,植物的净光合速率(Pn)会随着外界胁迫强度的增大而降低[36- 37]。盐碱混合胁迫下植物Pn的降低不仅与光合能力的损伤和气孔导度(Gs)的下降有关,而且也与植物细胞内矿物质元素的不平衡密切相关[38]。Pn降低的原因有气孔部分关闭导致的气孔限制和叶肉细胞光合活性的下降导致的非气孔限制,当胞间CO2摩尔分数(Ci)、Gs同时下降时,说明Pn的下降是由气孔限制造成的[39]。已有研究表明,在盐碱胁迫下,Pn受到气孔因素和非气孔因素的双重影响[12],盐碱胁迫程度较低时Pn的下降受气孔因素的影响,盐碱胁迫程度较高时Pn的下降主要是受非气孔因素以及气孔因素与非气孔因素的协同影响[40]。本研究表明,随胁迫溶液盐浓度和pH的增大,雾冰藜的Pn、Tr、Ci、Gs等气体交换参数均呈下降趋势,且雾冰藜光合色素含量的变化趋势和气体交换参数的变化趋势一致,说明雾冰藜在盐碱复合胁迫下Pn的下降是由气孔限制和非气孔限制共同造成的,这与薛延丰等利用NaCl和Na2CO3在菊芋上获得的研究结果一致[41]。本研究发现,在盐浓度和pH最高和最低的处理组中,雾冰藜WUE的变化趋势基本一致,但在其他处理组中其变化趋势比较复杂,造成这一现象的机理有待进一步的研究。
光合作用中电子激发能的捕获和能量转换由PSⅠ和PSⅡ协调完成,当PSⅠ和 PSⅡ间的激发能捕获与能量转换失衡时,会导致两个光系统间电子传递或激发状态的不平衡,从而影响光合作用的效率[42]。PSⅡ 原初反应的最大量子效率(Fv/Fm)和PSⅡ电子传递量子效率(ΦPSII)是判断植物是否受到光抑制的重要指标,是研究植物对逆境胁迫响应的理想参数[43]。在非胁迫条件下,植物通常表现出较高的Fv/Fm和ΦPSII值,可以使植物获得较高的光能转化效率,从而有利于能量的同化积累[44]。当植物受到营养元素缺乏、病害、温度、光照、盐分等胁迫时,PSⅡ复合体受到破坏,抑制了电子传递速率和光化学量子效率,使Fv/Fm和ΦPSII的值下降[45- 49]。光化学猝灭系数(qP) 反映了 PSⅡ原初电子受体的还原状态,而非光化学猝灭系数(NPQ) 则是衡量过剩激发能耗散的指标[50]。本研究表明,随着胁迫溶液盐浓度和pH的增大,雾冰藜的Fv/Fm和ΦPSII显著降低,ETR和qP等指标也随之降低,说明盐碱胁迫导致了雾冰藜叶绿体PSⅡ反应中心的损伤和光合活性的下降。刘建新等[51]分别用0、50、100、150、200 mmol/L的NaCl和Na2CO3处理燕麦时发现,随着处理浓度的增加,其Fv/Fm、ΦPSII和qP明显下降,研究结果与本文一致。雾冰藜非光化学淬灭系数(NPQ)随盐碱胁迫强度的增大基本呈现先降低后升高再降低的变化趋势,且在溶液盐浓度相同的条件下溶液pH较高的处理NPQ也较高,这表明雾冰藜能够保持较高的NPQ 从而提高热耗散消耗来适应盐碱胁迫,且碱性盐胁迫对雾冰藜的伤害作用大于中性盐胁迫,这与陈志强等在另一种耐盐碱植物西伯利亚白刺上利用苏打盐溶液处理所得研究结果一致[52]。然而在盐碱胁迫生境下,雾冰藜是否能够通过假环式电子传递保护过剩光能对光合机构的伤害,ROS清除系统是否能够做出相应的生理变化需进一步证实。
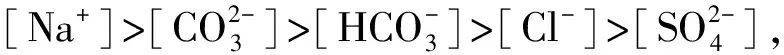
[1] Foyer C H, Noctor G. Tansley review No. 112: oxygen processing in photosynthesis: regulation and signalling. New Phytologist, 2000, 146(3): 359- 388.
[2] Robert H, Le Marrec C, Blanco C, Jebbar M. Glycine betaine, carnitine, and choline enhance salinity tolerance and prevent the accumulation of sodium to a level inhibiting growth ofTetragenococcushalophila. Applied and Environmental Microbiology, 2000, 66(2): 509- 517.
[3] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Reviews of Plant Physiology, 2011, 33: 317- 345.
[4] Tanji K K. Agricultural salinity assessment and management. New York: American Society of Civil Engineers, 1990: 1- 18.
[5] 程广有, 侯杰, 唐晓杰, 周喜云. 3种枸杞耐盐碱性的比较. 东北林业大学学报, 2007, 35(11): 47- 49.
[6] 赵可夫. 盐生植物资源及盐碱土改良利用研究动态. 资源与环境, 1989, 1(1): 40- 43.
[7] 石德成, 盛艳敏, 赵可夫. 不同盐浓度的混合盐对羊草苗的胁迫效应. 植物学报, 1998, 40(12): 1136- 1142.
[8] 张科, 田长彦, 李春俭. 一年生盐生植物耐盐机制研究进展. 植物生态学报, 2009, 33(6): 1220- 1231.
[9] Yang J Y, Zheng W, Tian Y, Wu Y, Zhou D W. Effects of various mixed salt-alkaline stresses on growth, photosynthesis, and photosynthetic pigment concentrations ofMedicagoruthenicaseedlings. Photosynthetica, 2011, 49(2): 275- 284.
[10] Liu J, Shi D C. Photosynthesis, chlorophyll fluorescence, inorganic ion and organic acid accumulations of sunflower in responses to salt and salt-alkaline mixed stress. Photosynthetica, 2010, 48(1): 127- 134.
[11] Wang H M, Wang W J, Wang H Z, Wang Y, Xu H N, Zu Y G. Effect of inland salt-alkaline stress on C4enzymes, pigments, antioxidant enzymes, and photosynthesis in leaf, bark, and branch chlorenchyma of poplars. Photosynthetica, 2013, 51(1): 115- 126.
[12] Chen W, Feng C, Guo W, Shi D, Yang C. Comparative effects of osmotic-, salt-and alkali stress on growth, photosynthesis, and osmotic adjustment of cotton plants. Photosynthetica, 2011, 49(3): 417- 425.
[13] 刘涛, 耿文春, 李丽, 刘一佳, 刘慧民. 混合盐碱胁迫对两种抗性不同的绣线菊光合特性的影响. 东北农业大学学报, 2009, 40(5): 32- 36.
[14] 王波, 张金才, 宋凤斌, 赵敏, 韩希英. 盐碱胁迫对燕麦光合特性的影响. 中国农学通报, 2007, 23(5): 235- 238.
[15] 王艳杰, 薛达元, 彭羽. 盐碱胁迫对两个葡萄品种光合作用-光响应特性的影响. 北方园艺, 2011, (16): 30- 36.
[16] Guo C Y, Wang X Z, Chen L, Ma L N, Wang R Z. Physiological and biochemical responses to saline-alkaline stress in two halophytic grass species with different photosynthetic pathways. Photosynthetica, 2015, 53(1): 128- 135.
[17] 张德魁, 马全林, 刘有军, 李亚, 陈芳. 河西走廊荒漠区一年生植物组成及其分布特征. 草业科学, 2009, 26(12): 37- 41.
[18] 李雪华, 李晓兰, 蒋德明, 刘志民, 于庆和. 干旱半干旱荒漠地区一年生植物研究综述. 生态学杂志, 2006, 25(7): 851- 856.
[19] Gutterman Y. Environmental factors and survival strategies of annual plant species in the Negev Desert, Israel. Plant Species Biology, 2000, 15(2): 113- 125.
[20] 张林静, 岳明, 张远东, 顾峰雪, 潘晓玲, 赵桂仿. 新疆阜康绿洲荒漠过渡带主要植物种的生态位分析. 生态学报, 2002, 22(6): 969- 972.
[21] 袁建立, 张景光, 王刚. 沙坡头人工固沙区草本层片组成及其动态研究. 中国沙漠, 2002, 22(6): 623- 627.
[22] 梁继业, 王彦阁, 杨晓晖. 鄂尔多斯高原雾冰藜在沙漠-河岸过渡带白刺沙堆上的空间分布特征. 草业科学, 2008, 25(5): 42- 47.
[23] 梁言慈, 郭晓花, 梁汉东, 朱书全. 乌达煤田雾冰藜汞的初步调查. 地质论评, 2015, 61(4): 883- 891.
[24] 吕彪, 许耀照, 赵芸晨. 河西走廊内陆盐渍土生物修复与调控研究. 水土保持通报, 2008, 28(3): 198- 200.
[25] 王遵亲. 中国盐渍土. 北京: 科学出版社, 1993: 38- 76.
[26] 李合生.植物生理生化试验原理与技术[M].北京:高等教育出版社,2000:134- 137.
[28] 吴旭红, 董雪芹. 碱胁迫对甜瓜幼苗生长及光合色素含量和抗氧化作用的影响. 种子, 2011, 30(8): 62- 64.
[29] Parida A, Das A B, Das P. NaCl stress causes changes in photosynthetic pigments, proteins, and other metabolic components in the leaves of a true mangrove,Bruguieraparviflora, in hydroponic cultures. Journal of Plant Biology, 2002, 45(1): 28- 36.
[30] Li C Y, Fang B, Yang C W, Shi D C, Wang D L. Effects of various salt-alkaline mixed stresses on the state of mineral elements in nutrient solutions and the growth of alkali resistant halophyteChlorisvirgata. Journal of Plant Nutrition, 2009, 32(7): 1137- 1147.
[31] Yang C W, Chong J N, Li C Y, Kin C, Shi D C, Wang D L. Osmotic adjustment and ion balance traits of an alkali resistant halophyteKochiasieversianaduring adaptation to salt and alkali conditions. Plant and Soil, 2007, 294(1/2): 263- 276.
[32] Koyro H W. Effect of salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophytePlantagocoronopus(L.). Environmental and Experimental Botany, 2006, 56(2): 136- 146.
[33] 闫萌萌, 王铭伦, 王洪波, 王月福, 赵长星. 光质对花生幼苗叶片光合色素含量及光合特性的影响. 应用生态学报, 2014, 25(2): 483- 487.
[34] Aro E M, McCaffery S, Anderson J M. Photoinhibition and D1 protein degradation in peas acclimated to different growth irradiances. Plant Physiology, 1993, 103(3): 835- 843.
[35] Demmig-Adams B, Adams III W W. The role of xanthophyll cycle carotenoids in the protection of photosynthesis. Trends in Plant Science, 1996, 1(1): 21- 26.
[36] Sultana N, Ikeda T, Itoh R. Effect of NaCl salinity on photosynthesis and dry matter accumulation in developing rice grains. Environmental and Experimental Botany, 1999, 42(3): 211- 220.
[37] Wei Y, Xu X, Tao H, Wang P. Growth performance and physiological response in the halophyteLyciumbarbarumgrown at salt-affected soil. Annals of Applied Biology, 2006, 149(3): 263- 269.
[38] Yang C W, Xu H H, Wang L L, Liu J, Shi D C, Wang D L. Comparative effects of salt-stress and alkali-stress on the growth, photosynthesis, solute accumulation, and ion balance of barley plants. Photosynthetica, 2009, 47(1): 79- 86.
[39] Farquhar G D, Sharkey T D. Stomatal conductance and photosynthesis. Annual Review of Plant Physiology, 1982, 33: 172- 345.
[40] Kong D, Shi H B, Li Y L, Zhang Y Q, Yan Y. Studies on photosynthetic rate of oil sunflower under difference salt stresses. Agricultural Research in the Arid Areas, 2005, 23(1): 111- 115.
[41] 薛延丰, 刘兆普. 不同浓度NaCl和Na2CO3处理对菊芋幼苗光合及叶绿素荧光的影响. 植物生态学报, 2008, 32(1): 161- 167.
[42] 高奔, 宋杰, 刘金萍, 隋娜, 范海, 王宝山. 盐胁迫对不同生境盐地碱蓬光合及离子积累的影响. 植物生态学报, 2010, 34(6): 671- 677.
[43] 吴晓丽, 汤永禄, 李朝苏, 吴春, 黄钢. 不同生育时期渍水对冬小麦旗叶叶绿素荧光及籽粒灌浆特性的影响. 中国生态农业学报, 2015, 23(3): 309- 318.
[44] Maxwell K, Johnson G N. Chlorophyll fluorescence-a practical guide. Journal of Experimental Botany, 2000, 51(345): 659- 668.
[45] 李强, 罗延宏, 余东海, 孔凡磊, 杨世民, 袁继超. 低氮胁迫对耐低氮玉米品种苗期光合及叶绿素荧光特性的影响. 植物营养与肥料学报, 2015, 21(5): 1132- 1141.
[46] 杨志晓, 丁燕芳, 张小全, 薛刚, 王轶, 任学良, 任周营, 杨铁钊. 赤星病胁迫对不同抗性烟草品种光合作用和叶绿素荧光特性的影响. 生态学报, 2015, 35(12): 4146- 4154.
[47] 徐兴利, 金则新, 何维明, 王兴龙, 车秀霞. 不同增温处理对夏蜡梅光合特性和叶绿素荧光参数的影响. 生态学报, 2012, 32(20): 6343- 6353.
[48] 王建华, 任士福, 史宝胜, 刘炳响, 周玉丽. 遮荫对连翘光合特性和叶绿素荧光参数的影响. 生态学报, 2011, 31(7): 1811- 1817.
[49] 周丹丹, 刘德玺, 李存华, 陈苗苗, 刘桂民, 杨庆山, 李永涛. 盐胁迫对朴树和速生白榆幼苗光合特性及叶绿素荧光参数的影响. 西北植物学报, 2016, 36(5): 1004- 1011.
[50] Aroca R, Irigoyen J J, Sánchez-díaz M. Drought enhances maize chilling tolerance. Ⅱ. Photosynthetic traits and protective mechanisms against oxidative stress. Physiologia Plantarum, 2003, 117(4): 540- 549.
[51] 刘建新, 王金成, 王瑞娟, 贾海燕. 盐、碱胁迫对燕麦幼苗光合作用的影响. 干旱地区农业研究, 2015, 33(6): 155- 160.
[52] 陈志强, 李庆贱, 时瑞亭, 张建秋, 王国柱, 张玉玲, 李悦. 苏打盐碱胁迫对西伯利亚白刺光合和生长的影响. 北京林业大学学报, 2011, 33(3): 31- 37.
[53] 石德成, 李玉明, 杨国会, 李毅丹, 赵可夫. 盐碱混合生态条件的人工模拟及其对羊草胁迫作用因素分析. 生态学报, 2002, 22(8): 1323- 1332.
猜你喜欢
草业学报(2022年5期)2022-07-08
干旱地区农业研究(2022年1期)2022-01-28
金桥(2021年11期)2021-11-20
现代畜牧科技(2021年4期)2021-07-21
草业学报(2021年3期)2021-03-22
装备制造技术(2019年12期)2019-12-25
科技创新与品牌(2019年12期)2019-02-06
草业学报(2018年10期)2018-10-19
草业科学(2018年7期)2018-07-26
西藏科技(2016年8期)2016-09-26
