
枫杨种子休眠与萌发特性研究
2018-03-20 09:22王宝龙徐君霞李明月赵彤彤张鹏
森林工程 2018年2期
王宝龙,徐君霞,李明月,赵彤彤,张鹏,*
0 引言
枫杨(Pterocarya stenoptera C. DC.)属于胡桃科(Juglandaceae)枫杨属,速生落叶阔叶乔木,长江流域和淮河流域重要的优良乡土改造树种,以种子繁殖为主[1-3];枫杨耐湿性极强, 根系发达对堤岸具有保护作用[4-8]。枫杨滞灰能力较强,姿态优美,是很好的行道绿化树种[9-10]。枫杨对氟离子和镉的耐受力和积累能力很强,适宜水体氟、镉污染严重区域水体净化和植被的恢复[11-14]。近几年,有研究发现,枫杨属植物的乙醇提取液还具有很强的抗肿瘤活性,对油茶炭疽病有较强抑制作用[15-17]。枫杨叶因含没食子酸、槲皮素等成分而具有杀死钉螺的作用,因此是长江中下游地区江湖滩地兴林灭螺及综合开发的主要造林树种[18],但20世纪70年代后由于城乡建设枫杨大量被砍伐,使得枫杨数量急剧减少[19]。国外对枫杨的研究很少,国内研究主要集中在种源的分布,抗逆性,灭螺生态系统的应用,枫杨提取物的纯化、成分分析及医疗应用等方面[13-15,18,20-25],对其休眠原因及解除方法缺乏研究。近几年,随着人们对枫杨的重视,枫杨造林用苗量迅速增加,但因其种子具体休眠性,给播种育苗造成了一定困难。因此,系统研究枫杨种子休眠与萌发特性对其育苗生产实践意义重大。本文以成熟的枫杨种子为材料,研究其种子休眠原因、解除方法及适宜萌发温度条件,为其种子繁殖提供理论和技术依据。
1 研究材料与方法
1.1 试验材料
试验用枫杨种子(翅果)采自东北林业大学校内植物园中的成年母树上。种子在室内阴干后置于冰箱中(3℃)冷藏备用,种子千粒重56.67 g。
1.2 研究方法
1.2.1 种子透水性测定
取10粒种子(去种翅)称重后置于烧杯中用自来水浸泡,每24 h取出种子用滤纸擦干水分后称重,种子鲜重不再增加时结束。绘制种子吸水曲线。
1.2.2 种子浸提物的生物测定
取部分种子(5 g)置于80%甲醇溶液中,用塑料薄膜封瓶口,置于0~4℃条件下避光浸提48 h,经减压蒸馏后定容至5 mL蒸馏水中作为原液(1 g/ml),再将部分原液稀释5倍(0.2 g/ml)和10倍(0.1 g/ml)。用不同浓度种子浸提溶液进行白菜种子的生物测定。白菜种子置于垫有一层滤纸的直径为9 mm塑料培养皿中,每皿滴入3 mL浸提液,以加入3 mL蒸馏水处理为对照,每种处理50粒种子,4次重复。将培养皿置于25℃恒温培养箱中(8 h光照)培养,24 h后计算白菜种子发芽率。
1.2.3 种子解除休眠处理
采种当年11月上旬,取部分种子浸种48 h,然后用0.5%的高锰酸钾溶液消毒30 min,消毒后的种子分别进行以下3种解除休眠处理。处理1:种子置于网袋中,不与任何基质混和,埋藏于室外深度为10 cm的土壤中(以下简称直接埋藏);处理2:种子与雪混和置于网袋中,埋藏于室外10 cm土壤中催芽(以下简称为混雪后埋藏);处理3:于室内5℃培养箱中进行混沙层积(低温层积)。所有处理种子于翌年4月下旬取出,于培养箱中25℃黑暗条件下进行发芽试验。
11月上旬,取部分种子,经浸种消毒后与细河沙均匀混和后于室内5℃培养箱中进行低温层积催芽。于种子催芽的30、60、90、120d分别取出部分种子,置于培养箱中25℃黑暗条件下进行发芽试验,观察低温层积处理时间对种子萌发的影响。
1.2.4 种子萌发温度测验
11月上旬,经浸种消毒后的种子于室内5℃培养箱中进行混沙低温层积催芽。种子层积120 d时,取出部分种子于培养箱中黑暗条件下进行发芽试验。共设置4个恒温萌发温度:10、15、20、25℃。
1.2.5 种子发芽测定方法
发芽实验时,各处理种子置于直径为9 cm的塑料培养皿中,其底部垫有一层滤纸和脱脂棉。培养皿放入培养箱中进行萌发试验,每种处理200粒种子,每皿50粒,4次重复,每日观察并记录种子发芽情况,以胚根突破种子并超过2 mm作为种子萌发的标志。发芽指标计算公式如下:
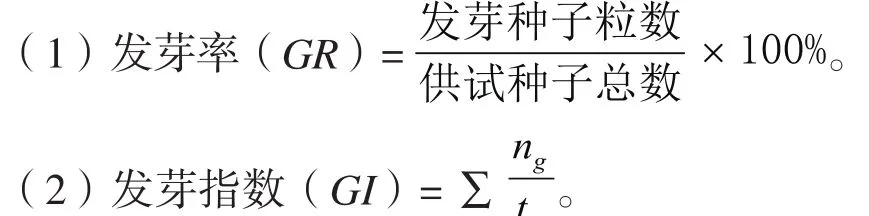
式中:tg为发芽时间,d;ng为与tg相对应的每天发芽种子数。
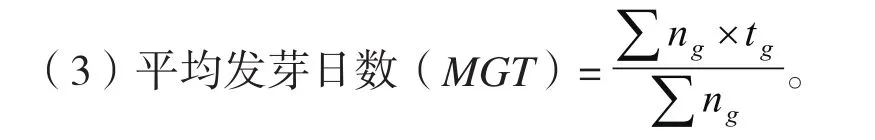
式中:tg为发芽时间,d,ng为与tg相对应的每天发芽种子数。
1.2.6 数据统计分析方法
试验数据利用SPSS13.0进行数据统计分析,并利用Excel 2003软件绘图。
2 结果与分析
2.1 枫杨种子的透水性
枫杨种子吸水后的鲜重变化(图1)显示,种子浸种后1 d内鲜重迅速增加,比浸种前增加了72.85%,第2~6天种子吸水量迅速减少,基本不再吸收更多的水分。由此可见,枫杨种皮并不能阻碍种子吸水,且种子可以在24 h内吸收足够的水分,种皮透水性不存在问题,不是引起种子休眠的原因。
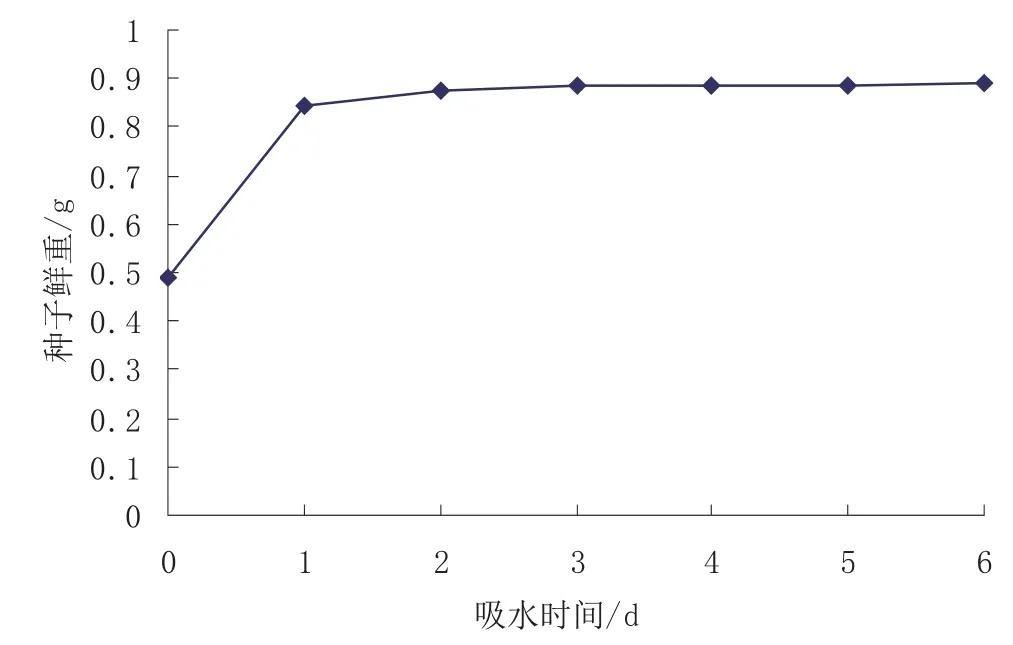
图1 枫杨种子吸水后鲜重变化Fig.1 Fresh weight changes of Pterocarya stenoptera seeds after water absorption
2.2 枫杨种子浸提液的抑制物质活性
白菜种子在不同浓度枫杨种子浸提液中的发芽率差异极显著(p<0.01)。多重比较结果(图2)显示,白菜种子在枫杨种子不同浓度浸提液中的发芽率随着浸提液浓度的降低而升高,但都显著低于对照。这表明枫杨种子浸提液中存在发芽抑制物质,抑制物质可能是造成种子不能直接萌发的主要原因。
2.3 不同催芽方法对枫杨种子萌发的影响
2.3.1 不同催芽方法对枫杨种子萌发的影响
不同催芽方法处理的种子发芽率差异极显著(p<0.01)。多重比较结果(图3)显示,低温层积催芽处理种子发芽率最高为84%,显著高于直接埋藏(34%)和混雪后埋藏(13%),混雪后埋藏处理种子的发芽率显著低于其它处理。
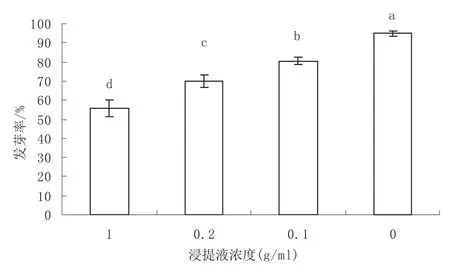
图2 白菜种子在不同浓度枫杨种子浸提液中的发芽率Fig.2 Germination rate of cabbage seeds in different concentration extracts of Pterocarya stenoptera seed
不同催芽方法处理种子的发芽指数差异极显著(p<0.01)。多重比较结果(图3)显示,室内低温层积催芽处理种子发芽指数最高为6.50,显著高于直接埋藏处理(1.20)和混雪后埋藏处理(0.35),混雪后埋藏处理种子的发芽指数显著低于其它处理。
不同催芽方法处理种子的发芽时间差异极显著(p<0.01)。多重比较结果(图3)显示,室内低温层积催芽处理种子发芽时间最短为3.82 d,显著短于直接埋藏处理(7.49 d)和混雪后埋藏处理(9.96 d),混雪后埋藏处理种子的发芽时间显著长于室内低温层积和直接埋藏处理。
2.3.2 不同低温层积时间对枫杨种子萌芽的影响
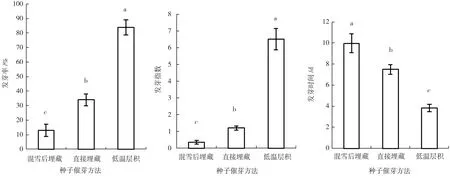
图3 不同催芽方法处理枫杨种子的发芽率、发芽指数和发芽时间Fig.3 The germination rate, germination index and germination time of Pterocarya stenoptera seeds under different pretreatment method
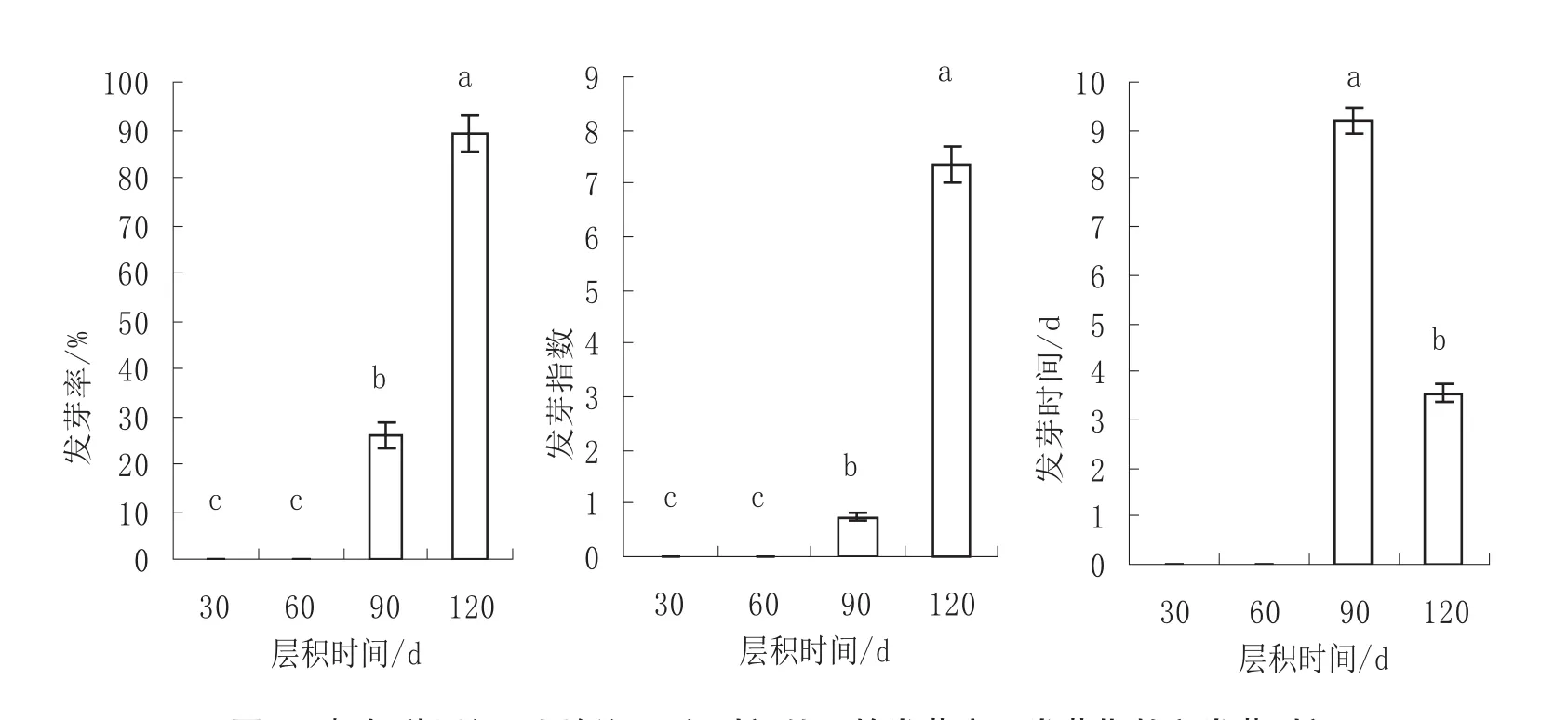
图4 枫杨种子经不同低温层积时间处理的发芽率、发芽指数和发芽时间Fig.4 The germination rate, germination index and germination time of Pterocarya stenoptera seeds after different time of stratification
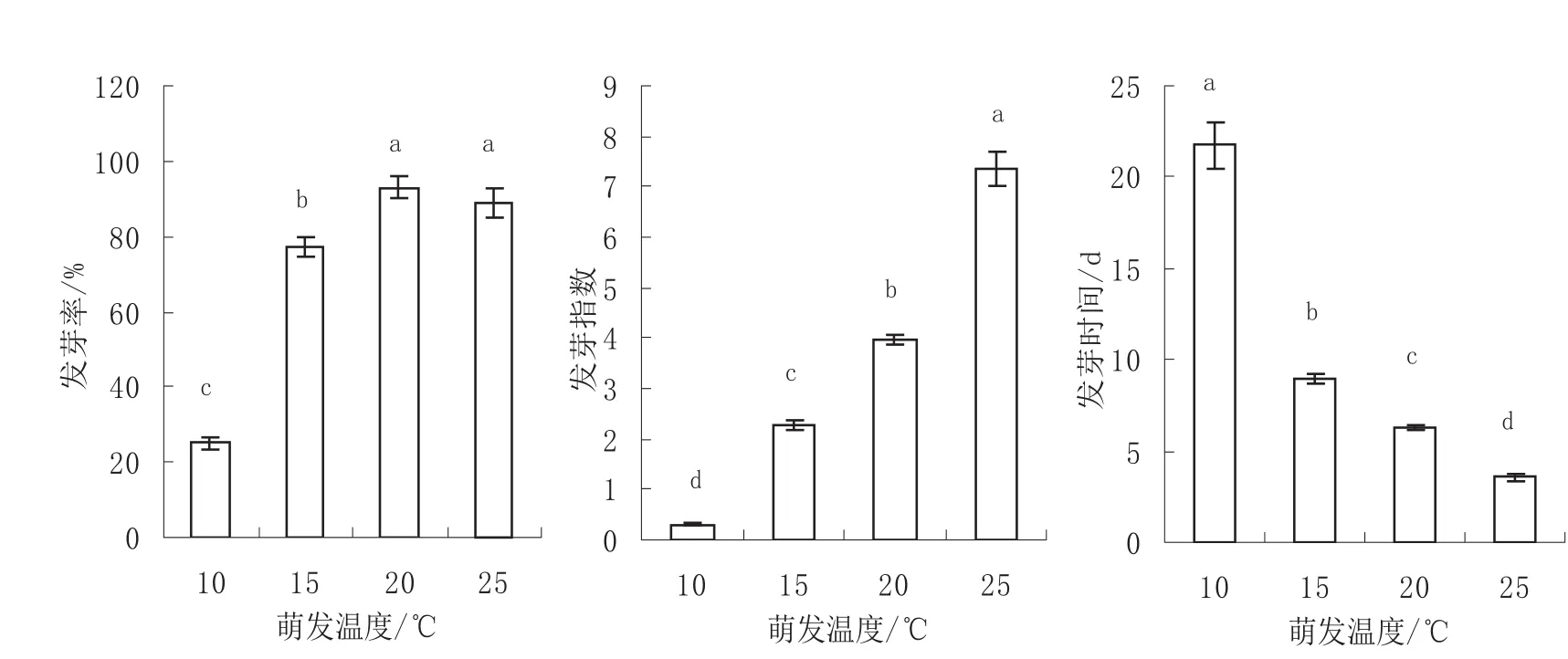
图5 不同萌发温度下枫杨种子的发芽率、发芽指数和发芽时间Fig.5 The germination rate, germination index and germination time of Pterocarya stenoptera seeds germinated in different temperatures
不同低温层积时间处理种子的发芽率差异极显著(p<0.01)。多重比较结果(图4)显示,低温层积催芽120 d时种子发芽率最高为89%,显著高于低温层积30 d(0)、60 d(0)和90 d(26%)的催芽处理。低温层积催芽30 d和60 d时,种子仍然处于休眠状态,没有种子萌发,低温层积90 d时只有少部分种子解除了休眠。
不同低温层积时间处理种子的发芽指数差异极显著(p<0.01)。多重比较结果(图4)显示,低温层积催芽120 d时种子发芽指数最高为7.34,显著高于低温层积30 d(0)、60 d(0)和 90 d(0.75)的催芽处理。
由于低温层积催芽30 d和60 d时,种子仍然处于休眠状态,没有种子萌发,所以无法用公式计算其平均发芽时间,因此只计算比较了低温层积催芽90 d和120 d时的平均发芽时间。不同低温层积时间下枫杨种子发芽时间的方差分析结果表明,低温层积催芽90 d和120 d时种子的发芽时间差异极显著(p<0.01)。图4显示,低温层积120 d时种子发芽时间最短为3.15 d,显著短于低温层积90 d时的发芽时间9.20 d。
2.4 不同温度对枫杨种子萌发的影响
不同萌发温度下枫杨种子的发芽率差异极显著(p<0.01)。多重比较结果(图5)显示,恒温20℃条件下种子发芽率最高为93%,其次是20℃条件下种子发芽率较高为89%,两者差异不显著,但都显著高于15℃(77%)和10℃(25%)条件下种子的发芽率。10℃条件下种子的发芽率最低,显著低于其它温度处理。
不同萌发温度下枫杨种子的发芽指数差异极显著(p<0.01)。多重比较结果(图5)显示,各温度条件下种子发芽指数差异均显著,恒温25℃条件下种子发芽指数最高为7.34,显著高于其它温度处理。20℃和15℃条件下种子发芽指数分别为3.96和2.28。10℃条件下种子的发芽指数最低为0.30,显著低于其它温度处理。
不同萌发温度下枫杨种子的发芽时间差异极显著(p<0.01)。多重比较结果(图5)显示,各温度条件下种子发芽时间差异均显著,恒温25℃条件下种子发芽时间最短为3.55 d,显著短于其它温度处理。20℃和15℃条件下种子发芽时间分别为6.29 d和8.96 d。10℃条件下种子的发芽时间最长为21.73d,显著长于其它温度处理。
3 结论
枫杨种子透水性良好,不存在物理休眠;枫杨种子中存在发芽抑制物质,抑制物质的存在可能是造成种子休眠的主要原因;混沙低温层积(120d)解除枫杨种子休眠效果比直接埋藏和混雪后埋藏催芽好,种子经层积处理后发芽率可达到89%;温度显著影响枫杨种子的萌发,25℃的恒温条件下种子萌发效果好。
[1]徐有明, 邹明宏, 史玉虎, 等.枫杨的生物学特性及其资源利用的研究进展[J].东北林业大学学报, 2002, 30(3): 42-48.
[2]欧斌, 卜明生, 李远章. 枫杨种子秋播育苗技术[J]. 林业科技开发, 2004, 18(4): 58-59.
[3]阳著平, 秦景寞. 枫杨的繁殖与栽培[J]. 中国林业, 2009 (19):49.
[4]王朝英, 李昌晓, 张晔.水淹对枫杨幼苗光合生理特征的影响[J].应用生态学报, 2013, 24(3): 675-682.
[5]谢小红, 魏虹, 李昌晓, 等.水淹胁迫下枫杨(Pterocarya stenoptera C.DC.)幼苗叶片高光谱特征的研究[J].西南大学学报(自然科学版), 2011, 33(4): 93-98.
[6]王振夏, 魏虹, 吕茜, 等.枫杨幼苗对土壤水分“湿-干”交替变化光合及叶绿素荧光的响应[J].生态学报, 2013,33(3): 888-897.
[7]衣英华, 樊大勇, 谢宗强, 等.模拟淹水对枫杨和栓皮栎气体交换、叶绿素荧光和水势的影响[J].植物生态学报, 2006,30(6): 960-968.
[8]李侠. 枫杨在防洪护岸中的应用[J]. 科技与企业, 2012 (18):290-290.
[9]张家洋, 周君丽, 任敏, 等.20种城市道路绿化树木的滞尘能力比较[J].西北师范大学学报(自然科学版), 2013,49(5): 113-120.
[10]李昂.以特色空间为引导的旧城街道更新改造研究——以沈阳枫杨路街道更新规划为例[J].科技视界, 2014,(11): 310-311.
[11]曲木子, 谢会成, 李传荣, 等.枫杨幼苗对氟离子的吸收积累规律及毒性效应[J].林业科学, 2015, 51(4): 156-163.
[12]贾中民, 魏虹, 孙晓灿, 等.秋华柳和枫杨幼苗对镉的积累和耐受性[J].生态学报, 2011, 31(1): 107-114.
[13]贾中民, 冯汉茹, 魏虹.镉对枫杨幼苗生长和光合特性的影响[J].西南大学学报(自然科学版), 2014, 36(8): 27-35.
[14]陈海生, 黄志强, 冯伟荣.水库消落带植物枫杨降污效应研究[J].安徽农学通报, 2013, 19(16): 96-97.
[15]高双.枫杨树皮化学成分的研究[D].广州:暨南大学,2013.
[16]史红安, 魏晓, 王立华, 等.枫杨树各部位对油茶炭疽病的抑制作用研究[J].湖北工程学院学报, 2016, 36(6): 25-28.
[17]罗彭, 王佳佳, 李兵, 等.枫杨树皮抗菌物质的分离鉴定研究[J].天然产物研究与开发, 2015, 27(2): 246-250.
[18]王万贤.长江滩地九种抑螺防病林模型对钉螺的化感作用研究[A].中国植物保护学会植物化感作用专业委员会.中国第六届植物化感作用学术研讨会论文摘要集[C].中国植物保护学会植物化感作用专业委员会: 2013.
[19]李纪元, 饶龙兵, 潘德寿, 等.人工胁迫条件下枫杨种源MDA 含量的地理变异[ J].浙江林业科技, 1999, 19(4): 22-27.
[20]李纪元, 饶龙兵, 杨伟增. 枫杨种源苗期髙生长地理变异的趋势面分析[J].中南林学院学报, 2003, 23(1): 15-19.
[21]袁传武, 胡兴宜, 谢先祎, 等. 枫杨研究进展[J]. 湖北林业科技, 2011(3): 38-42.
[22]曲良谱, 喻方圆, 张新. 枫杨容器苗育苗技术研究[J]. 江苏林业科技, 2008, 35(2): 9-12.
[23]陈湘.枫杨树叶和骨碎补中化学成分的分离及结构鉴定[D].南昌:南昌大学, 2015.
[24]夏鹏飞.枫杨叶鞣质提取纯化工艺的研究[D].合肥:安徽理工大学, 2014.
[25]李文敏, 魏虹, 李昌晓, 等.基于高光谱参数的枫杨叶绿素含量估算模型优化[J].林业科学, 2014, 50(4): 55-59.
猜你喜欢
现代农业科技(2022年10期)2022-12-17
安徽农业科学(2022年18期)2022-10-13
现代仪器与医疗(2021年6期)2022-01-18
潍坊学院学报(2020年6期)2020-11-22
食品工程(2020年3期)2020-01-05
许昌学院学报(2018年8期)2018-09-05
扬子江(2018年1期)2018-01-26
计算机测量与控制(2017年6期)2017-07-01
决策与信息(2015年36期)2015-12-01
湖南农业科学(2015年11期)2015-01-10
