
高羊茅FaHsfC1b启动子克隆及外源激素处理下FaHsfC1b的表达分析
2018-03-14 06:20杨志民庄黎丽
江苏农业科学 2018年3期
曹 薇, 杨志民, 庄黎丽, 王 剑, 孙 岩
(南京农业大学草业学院,江苏南京 210095)
作为一种多年生冷季型草坪草,高羊茅(Festucaarundinacea)具有抗病虫害、成坪快、耐修剪等多种优点,因此在我国温带南部和亚热带地区得到了广泛的种植。在温带以北,如北京地区,高羊茅在越冬时遭受冷胁迫,导致其通常活不过2年,被称为“短命草”,而在亚热带地区,夏季高温以及高温带来的缺水也严重影响着高羊茅的生长,提高高羊茅对非生物胁迫的抵抗能力有益于节约草坪管理成本,进一步扩大高羊茅草坪草在我国的使用范围。
植物热激转录因子(heat stress transcription factors,Hsfs)是存在于植物细胞内调节热激蛋白表达的一类转录调节基因,在植物热胁迫信号转导和耐热性的产生过程中具有重要作用[1]。热激转录因子在结构上可以分为3类:A类、B类、C类。A类热激转录因子是热激基因表达的激活剂,在不同的植物中具有不同的应激反应[1],多项研究表明,此类热激转录因子在植物抗逆境尤其是抗热方面起到重要作用[2-4]。B类热激转录因子本身没有转录激活功能,在番茄抗热性形成的不同阶段中,HsfB1和HsfA1a可以形成一个增效的共激活子[5]。另外,在热胁迫下HsfB1可以跟其他热激转录因子结合控制或抑制看家基因的表达[6]。与A类Hsfs相类似,热胁迫下的转录组分析显示,C类热激转录因子HsfCs在水稻(Oryzasativa)、小麦(Triticumaestivum)、白菜(Brassicapekinensis)、椪柑(CitrusreticulataBlanco)、胡萝卜(Daucuscarota)、豌豆(Pisumsativum)、簸箕柳(Salixsuchowensis)中普遍上调表达,在其中某些物种中,HsfCs也响应渗透胁迫、冷胁迫、ABA[7-13]。到现在为止,对于C类热激转录因子的研究较少,仅发现水稻缺失OsHsfC1b后,对盐胁迫、干旱胁迫及ABA敏感,在非胁迫条件下,OsHsfC1b对水稻的生长是不可缺少的[14]。目前对多年生草坪草尤其是高羊茅中C类热激转录因子的功能及其上游调控因子缺乏研究。
启动子是DNA分子与RNA聚合酶特异结合的部位,负责下游基因的转录激活,对于基因的表达非常重要。伊淑莹[15]等研究获得了番茄线粒体小分子热激蛋白基因LeMTsHSP起始密码子上1 915 bp的调控序列,可能对该基因的表达具有增强作用。高温下小麦(Triticumaestivum)Hvhsp17启动子能驱动Hvhsp17在小麦中表达[16]。在不同类型的启动子中,有一类被称为激素诱导型启动子,激素首先与植物体内的特定受体相结合,受体蛋白被激活后与启动子结合,从而激活下游基因的表达,使植物发生一系列生理反应。如果植物缺少相应的受体基因或启动子,就不会对激素起反应[17]。目前研究较多的有水杨酸(SA)、脱落酸(ABA)、赤霉素(GA)、生长素(IAA、NAA等)等激素响应启动子。不同激素诱导型启动子需要具有相应的激素响应元件,比如在拟南芥RD29B启动子中存在2个ABA响应元件ABRE,只有这2个元件相互作用形成ABA应答复合体,才能使其所在的启动子表现出ABA诱导活性[18]。
植物激素在植物抗逆性中发挥着重要的作用,ABA含量在大豆热激恢复阶段会明显快速升高[19-20],而ABA不敏感型突变体abi1和abi2则表现为耐热性变差[21]。水稻中的OsHsfC1b被证实依赖ABA提高水稻对盐胁迫的抗性[14],并且已经在A类的热激转录因子中发现了富集ABA应答原件(ABRE)的顺式调控模块,这表明了ABA在一些热激转录因子转录调控中的介导作用[22]。SA能够在植物遭受逆境时及时清除ROS,维持抗氧化酶的活性,从而保护胁迫下细胞膜的稳定性[23]。生长素和赤霉素作为植物激素普遍参与了植物在正常条件和胁迫条件下的各种生理机制[24-27]。研究发现,大麦和拟南芥在热胁迫下,伴随着内源生长素含量的降低,编码生长素合成的关键基因表达量也出现下调,并导致雄性不育,这些性状可以通过外施生长素得到恢复[28]。赤霉素还参与调解生长素/细胞分裂素的拮抗作用[29]。
基于已有高羊茅品种凌志的全长序列,我们设计 5′RACE 引物,通过染色体步移法分离FaHsfC1b上游启动子并克隆,随后利用启动子分析网站对该基因启动子区域的顺式作用调控元件及其功能进行分析,并根据启动子中出现的激素响应元件,对激素处理下FaHsfC1b的表达量进行了测定,从而为高羊茅C类热激转录因子的分子调控机制研究提供科学依据。
1 材料与方法
1.1 植物材料与菌株
试验于2015年9—12月在南京农业大学草业科学学院草坪分子生物学实验室完成。高羊茅(Festucaarundinacea)品种凌志材料为笔者所在实验室草坪种质资源圃自有品种。选取实验室资源圃高羊茅植株,洗去根部泥土并去掉枯叶后将植株分蘖从整株上分离下来,剪去根部,将分蘖种植于霍格兰营养液[30]中,营养液上置泡沫板以海绵包裹分蘖节支撑植株,按照1/3原则修剪叶片,培养条件为白天、夜晚各12 h,白天温度25 ℃,夜晚温度20 ℃。其后每周换1次营养液,待 2~3周后,分蘖节发育成较大植株,侧根也长出10~15 cm,即可更换营养液并对高羊茅进行处理。
pMD19SimpleT载体购自TaKaRa公司,大肠杆菌菌株DH5α为笔者所在实验室冻存。引物合成及DNA测序均在南京思普金生物公司完成。
1.2 方法
1.2.1 高羊茅FaHsfC1b启动子的克隆 高羊茅FaHSFC1b启动子的克隆采用染色体步行法,具体步骤参考李广平等的方法[31]和BD Genome WalkerTMUniversal Kit User Manual说明书。2条基因特异反向引物分别为:NGSP1:5′-TCGT-GCTCTGCTCGGCGTGGT-3′;NGSP2:5′-TACCTCGCTCTGT-GCGTGCCG-3′。重新验证启动子所设计的引物序列为:F:5′-CAGAGCAGAAGTGGTGATGGT-3′;R:5′-GAGGAG-GAAGTCGGAGAAGGC-3′。
1.2.2FaHsfC1b启动子序列分析 采用GenBank在线比对工具BLASTn(http://blast. ncbi. nlm.nih. gov/)[32]进行了序列同源性分析,启动子序列调控元件分析利用植物顺势调控元件数据库PlANTCARE(http://bio informatics. psb. ugent. be/webtools/plantcare/html/)[33]和PLACE(http://www. dna. affrc. go. jp/PLACE/signalscan.html)[34]完成。
1.2.3 外源激素处理 将水培2周左右的高羊茅植株转移到分别含有ABA(100 μmol/L)、GA(90 mg/L)、IAA(60 mg/L)、SA(0.5 mmol/L)、ZT(0.5 mg/L)的营养液中处理24 h,在0、4、12、24 h取根部和叶片,每个处理取3个重复,培养条件与处理前相同。
1.2.4 高羊茅总RNA的提取与基因表达分析 总RNA提取采用试剂盒Plant RNA Kit(50)(Omega Bio-Tec,American),用于荧光定量PCR的cDNA采用试剂盒PrimeScriptTMRT reagent Kit with gDNA Eraser (Perfect Real Time)(TaKaRa,Otsu,Japan)合成,通过qRT-PCR检测FaHsfC1b在不同激素处理下的表达情况。采用SYBR Green Ⅰ Master反应体系(Roche Diagnostic,Rotkreuz,Switzerland),在LightCycler 480 Ⅱ(Roche,Rotkreuz,Switzerland)仪器中进行。以FaTublin作为内参基因,使用如下引物:RT-FaHsfC1b-F:ACTTCAAGCACCGCAACTTT;RT-FaHsfC1b-R:TGAACCGGCCTTCTTCTTCT;RT-FaTublin-F:ATGCTTT-CGTCTTATGCCC;RT-FaTublin-R:CTCTTGGTTTTGATGG-TTGC。
2 结果与分析
2.1 FaHsfC1b启动子克隆
构建的获得克隆经测序后与已有序列进行拼接和比对,以构建的高羊茅叶片基因组酶解DNA片段文库为模板,经过2轮PCR后,从高羊茅的EcoRⅠ酶切体系中扩增到1条 1 562 bp 左右的片段(图1)。将该片段进行克隆,连接到pMD19SimpleT并测序,结果显示,获得的片段实际长度为 1 514 bp。与已有的高羊茅FaHSFC1b基因(登录号:KY475613)序列比对后发现所得到的序列与高羊茅FaHSFC1b序列有95 bp的重叠区,表明所得到的序列是其上游的DNA序列。进一步设计1对验证引物,克隆分析整理后,共得到高羊茅FaHSFC1b基因上游长度为1 657 bp的DNA序列。

2.2 FaHsfC1b启动子序列生物信息学分析
通过顺式作用元件数据库PLACE和PlANTCARE对FaHsfC1b启动子的分析,结果发现FaHsfC1b启动子很多个位点上都有启动子核心元件TATA-box,即RNA聚合酶结合位点,确保了转录起始位点的高度灵活性,多个CAAT-box则控制着转录起始的频率(表1)。
除了核心元件,FaHsfC1b启动子中包含的元件按照功能大致可以分为6类(表1)。第一类为光信号相关元件,在启动子中数量最多、范围最广,多达16种,有-10PEHVPSBD、box Ⅱ、chs-Unit 1 m1、G-box、GAG-motif、GBOXLERBCS、I-box、INRNTPSADB、MNF1、MRE、PRECONSCRHSP70A、REBETALGLHCB21、SORLIP2AT、SORLIP1AT、Sp1、TBOXATGAPB。第二类是与抵抗逆境相关的元件,集中在干旱、冷/寒、盐及病害方面,包括综合性防御元件(同时具备抵抗2种或2种以上逆境相关元件):DOFCOREZM(多种胁迫防御)、DPBFCOREDCDC3(病原与高盐胁迫)、GT1GMSCAM4(病害与盐胁迫)包括脱水响应元件:DRECRTCOREAT、MYCATRD22;低温响应元件:LTRECOREATCOR15、LTRE1HVBLT49;抗病相关元件:GT1CONSENSUS、SEBFCONSSTPR10A、WBOXATNPR1。还有一种缺氧相关的缺氧特异性诱导增强元件:GC-motif,猜测可能与涝害相关。第三类是组织特异性元件,它们参与调控基因在不同植物组织中的表达,包括分生组织:CAT-box;叶肉细胞:CACTFTPPCA1;花粉:GTGANTG10、POLLEN1LELAT52;根瘤组织:OSE1ROOTNODULE、OSE2ROOTNODULE;根毛:RHERPATEXPA7;种子:RYREPEATBNNAPA;胚乳:Skn-1_motif。第四类为特殊物质响应元件,有蔗糖应答元件:RAV1AAT、SREATMSD;铜应答元件:CURECORECR;硫应答元件:SURECOREATSULTR11。第五类是一些与基因、酶及蛋白结合的位点。
此次研究重点观察到的第六类是激素响应元件,包括细胞分裂素调节基因ARR1结合位点:ARR1AT;脱落酸响应元件:ABRE、MYB2CONSENSUSAT;生长素响应元件:CATATGGMSAUR;乙烯响应元件:GCCCORE、WBOXNTERF3;水杨酸响应元件:TCA-element;赤霉素响应元件:PYRIMIDINEBOXOSRAMY1A、WRKY710S,另外WRKY710S元件也与创伤有关;调节类黄酮生物合成元件:MYBCORE。
综合上述分析,可知克隆到的启动子序列可能与光照周期、光照度有关,并且可能通过响应激素诱导,调控下游基因表达,在植物生长、抗病、抗逆方面发挥作用。
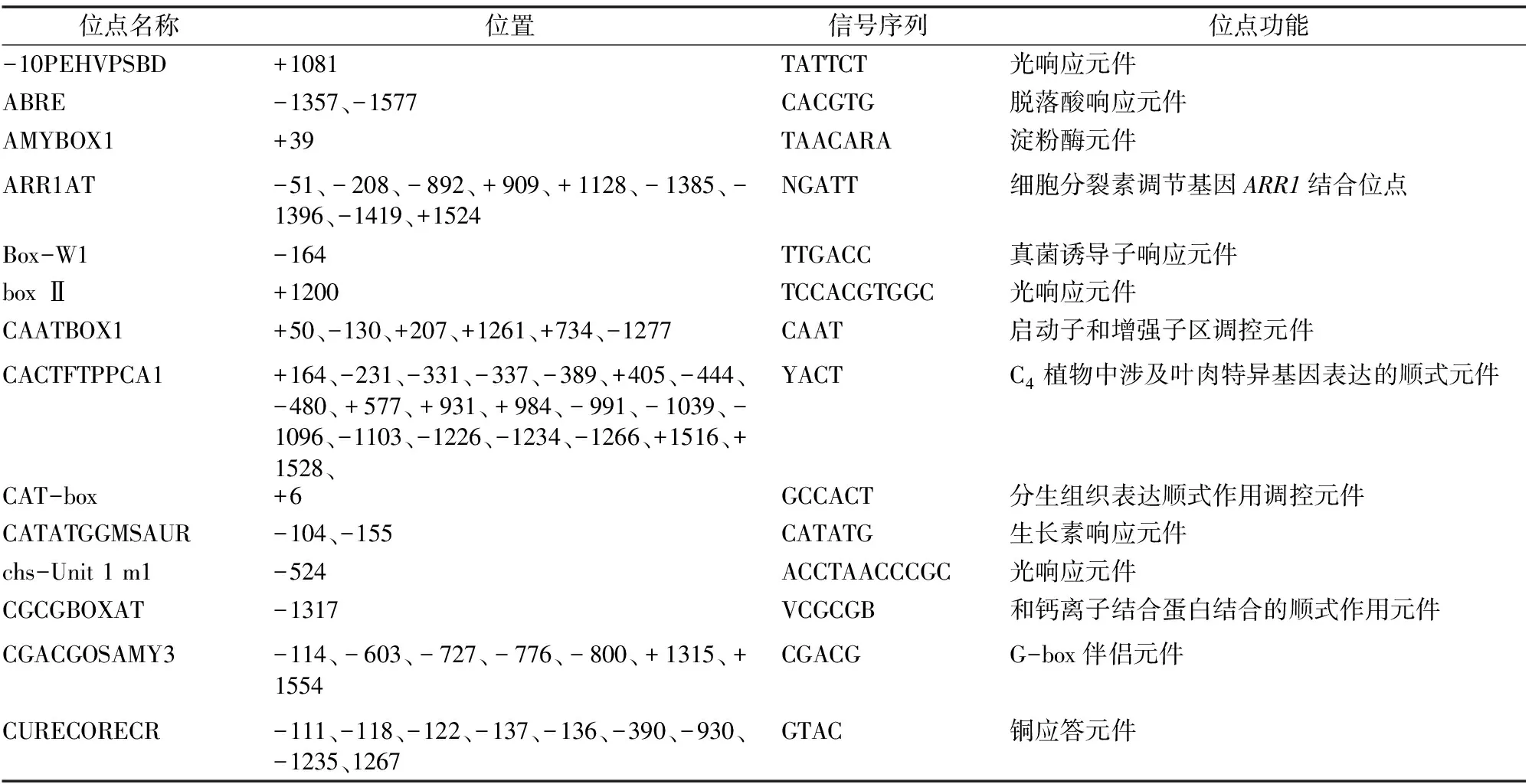
表1 应用PLANTCARE预测FaHsfC1b启动子顺式作用元件

位点名称位置信号序列位点功能DOFCOREZM+22、+238、-407、+857、-1078、+1101、-1134、-1144、+1285、-1296、+1460、-1623、AAAG防御与胁迫响应元件DPBFCOREDCDC3+147、-875、-1250、+1343、+1357、+1574ACACNNG病原和高盐胁迫响应元件DRECRTCOREAT-801、-1481RCCGAC脱水响应元件EBOXBNNAPA-96、-104、-989、-1242、-1344、1346、-1358、-1432、-1486、CANNTG储藏蛋白napA的E-boxEECCRCAH1+128、+340、+748、-1120、-1521GANTTNCMyb转录因子LCR1结合位点ELRECOREPCRP1-320TTGACC激发响应元件EMBP1TAEM+1358CACGTGGC抗旱、耐寒相关的反式作用因子GCCCORE-860、-882、+899GCCGCC乙烯响应元件G-Box-1202、+1421CACGTG光顺式作用调控元件GATABOX-47、-844、-1158、+1514、-1563、-1629GATAASF-2结合位点GAG-motif+310、+1149、471GGAGATG光响应元件GBOXLERBCS+1357MCACGTGGC光响应基因的上游保守序列GT1CONSENSUS-45、+1150、+1276、+1282、+1558GRWAAW调控抗病基因PR-1a表达,影响水杨酸诱导的基因表达GT1GMSCAM4+17、+1282GAAAAA在病原或盐诱导的SCaM-4基因表达中发挥作用GTGANTG10+30、+68、-87、+99、+315、-328、+338、-412、-821、-959、+1227、-1409GTGA花粉晚期基因启动子的元件GC-motif+1394CCCCCG缺氧特异性诱导增强元件Ⅰ-box-1626、-46GATAA部分光响应元件INRNTPSADB+355、-1220、-1277YTCANTYY光响应元件LECPLEACS2-191TAAAATATLeCp结合的顺式元件LTRE1HVBLT49+854CCGAAA低温响应元件LTRECOREATCOR15-810、-812、+1482、-1491、+1553CCGAC低温响应元件MNF1+56GTGCCC(A/T)(A/T)光响应元件MRE-341AACCTAA光响应MYB结合元件MYBCORE-561、-1464CNGTTR调节类黄酮(一类植保素)生物合成MYB2CONSENSUSAT+561YAACKG脱落酸信号转录激活因子MYCATRD22-96、+1344CACATG脱水响应元件OSE1ROOTNODULE+238AAAGAT根瘤组织特异性表达元件OSE2ROOTNODULE+427、+1076CTCTT根瘤组织特异性表达元件RAV1AAT+915、-1116、-1436CAACA蔗糖应答元件REBETALGLHCB21-1629CGGATA光敏色素调控元件RHERPATEXPA7-210、-605、+1606KCACGW根毛特异元件POLLEN1LELAT52+16、-408、-507、-674、-712、-1071、-1131、-1145、-1642AGAAA花粉特异表达的顺式作用元件ROOTMOTIFTAPOX1+191、-1561ATATTrolD的启动子元件PRECONSCRHSP70A-758、+886、+1372、+1553SCGAYNRNNNNNNNNNNNNNNNHD光响应元件PROLAMINBOXOSGLUB1+1098TGCAAAG葡聚糖酶1醇溶谷蛋白元件PYRIMIDINEBOXOSRAMY1A-1284CCTTTT赤霉素响应元件RYREPEATBNNAPA+1034CATGCA在种子特异性表达中发挥作用SEBFCONSSTPR10A+761、-1324YTGTCWC抗病相关元件SEF4MOTIFGM7S+263、-1402RTTTTTRSEF4结合位点Skn-1_motif+956GTCAT胚乳高水平表达顺式作用调控元件SORLIP2AT+160、-863、-947、+1030、+1139、-1363GGGCC光响应元件SORLIP1AT+162、-272、-550、-633、-689、-734、-880、+1311、-1349、-1361、+1538GCCAC光响应元件

位点名称位置信号序列位点功能Sp1-385、+1285、-587、-1391、-389、-1386、-744CC(G/A)CCC光响应元件SREATMSD+1628TTATCC蔗糖应答元件SURECOREATSULTR11-8、+646、-763、-956、+1324GAGAC硫应答元件TAAAGSTKST1-1078、-1134TAAAGstdof1蛋白反式作用控制保卫细胞特异基因表达的靶位点TATA-box-663、+33、+102、-39、-1014、-37、+339、+41、-1375、+34、+109、-40、-1015、-38、+361、+46ATATAT转录起始-30核心启动子元件TBOXATGAPB-1100ACTTTG涉及基因转录的光诱导激活TCA-element-434CAGAAAAGGA水杨酸响应顺式作用元件WBOXATNPR1-321TTGAC调控抗病基因NPR1表达,影响水杨酸诱导的基因表达WBOXNTERF3+31、+100、-152、-617、-1111TGACY参与乙烯诱导的ERF3基因的激活WRKY710S+75、-321TGACW-box核心区,赤霉素信号抑制因子
2.3 不同激素处理下FaHsfC1b的表达水平分析
为了验证FaHsfC1b启动子序列上存在的激素响应元件是否与FaHsfC1b在激素下的表达调控有关,我们对高羊茅进行了激素处理试验(图2),发现在ABA、GA、IAA、SA、ZT处理下,24 h内FaHsfC1b表达量均呈现上升再下降的趋势。ABA处理下,高羊茅根部FaHsfC1b表达量在4 h开始上升并在 12 h 达到峰值,为对照的49倍,叶片表达水平比根部低,同样在4 h开始上升并在12 h达到峰值,为对照的19倍。GA处理下根部和叶片中FaHsfC1b表达量都在4 h达到峰值,为对照的7~8倍,随后叶片中基因表达量虽有下降但依旧显著高于对照,而根部表达量则下降到处理前水平。IAA处理下高羊茅根部FaHsfC1b表达量明显快于也高于叶片中基因表达量,根部基因表达量在4 h达到峰值,并且是对照的42倍,而叶片则在12 h开始出现基因表达量的上升,与根部相比上升幅度较小,峰值为对照的11倍。SA和ZT处理下FaHsfC1b表达量在根部和叶片中的上升/下降趋势很相似,但SA处理下高羊茅根部基因表达量为所有处理中最高水平,在4 h达到峰值,为对照的65倍,叶片中FaHsfC1b表达量则在12 h开始上升并在24 h达到峰值,为对照的11倍;在ZT处理下,高羊茅根部FaHsfC1b表达量同样在4 h达到峰值,为对照的15倍,叶片在24 h达到峰值,是对照基因表达量的24倍。
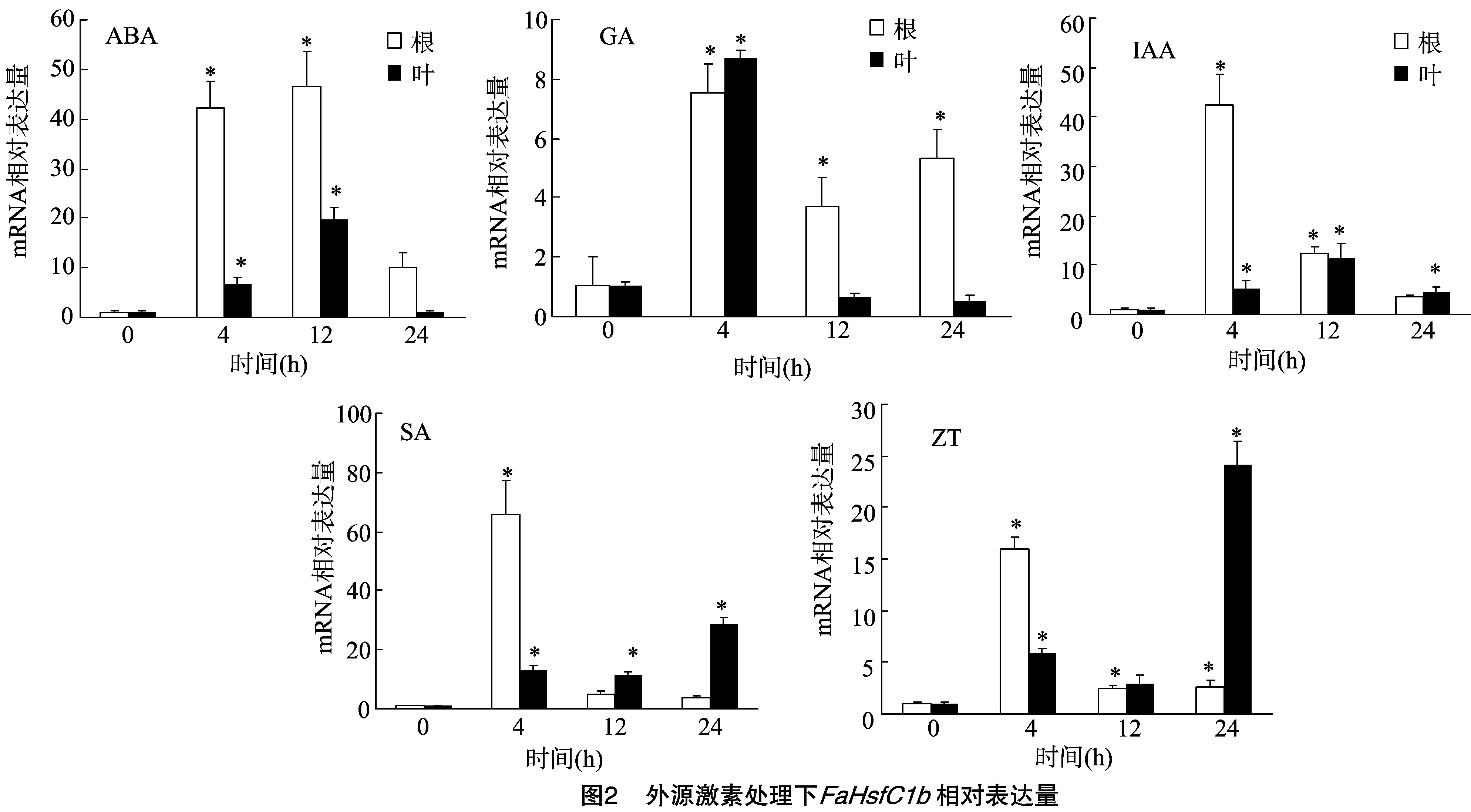
FaHsfC1b对以上5种激素的响应与启动子元件中5种激素响应元件相吻合,证明启动子所包含的响应元件与下游基因的调控表达有一定联系。
3 讨论与结论
FaHsfC1b启动子分析表明,该启动子序列中含有丰富的顺势作用元件和其他应答元件。参与低温响应的元件有6个,抗旱相关元件有4个,盐胁迫相关元件有8个,抗病相关元件有3个,推测FaHsfC1b可能与高羊茅抗寒、抗旱及抗盐相关,而已有的研究结果表明,HsfC1在渗透胁迫、盐胁迫和冷胁迫中的表达都很突出[35,37],将水稻中的OsHsfC1b敲除后,转基因水稻对渗透胁迫的抗性明显弱于野生型[14]。
在所有顺式作用元件中,数量最多的是涉及C4植物叶肉特异表达的顺式元件CACTFTPPCA1,此元件是维持C4植物叶肉特异性,因而得以更高效率地利用光照和氮资源的关键元件[38],另一个发现是关于光照的元件,在所有元件中光响应元件无论在种类还是数量上都是最多的,比较前人对其他基因启动子的分析,我们可以推测FaHsfC1b可能存在复杂的光应激反应表达模式,在植物正常的光合作用中起到重要作用。此外,FaHsfC1b启动子中还包含有多个蔗糖应答元件、铜应答元件、硫应答元件、真菌诱导子元件,猜测外源施加营养、金属元素或真菌感染可能会对FaHsfC1b的转录发生影响。启动子中含有的其他基因、酶及蛋白结合的位点说明植物对外界的应激反应是一个复杂的机制,需要多种物质共同协同作用。
对FaHsfC1b启动子序列分析的另一结果显示,激素响应元件所响应的激素不仅涵盖了五大类激素:生长素、细胞分裂素、赤霉素、脱落酸、乙烯,还包括水杨酸、类黄酮,其中细胞分裂素调节基因ARR1结合位点多达8个,在启动子的末梢还有个分生组织表达顺式作用调控元件CAT-box,猜测FaHsfC1b的转录表达可能对植物的生长有影响。大多数干旱应答基因都受脱落酸的诱导,已经在A类热激转录因子中发现了富集脱落酸应答元件的启动子序列[39],这表明脱落酸在C类热激转录因子转录调控中可能也有一定的介导作用。组织特异性元件的富集可推测FaHsfC1b的转录调控遍布高羊茅植株的各个部位。
在本试验中,FaHsfC1b响应IAA、ZT、GA、ABA和SA,在FaHsfC1b启动子中,生长素、赤霉素、脱落酸响应元件都有不止1个,而研究表明,启动子中多拷贝应答元件可能参与下游基因的增强表达[40]。当干旱胁迫应答基因上游启动子序列中含有ABRE元件时,该基因的表达调节依赖ABA调控途径,本课题组前期对FaHsfC1b的研究表明FaHsfC1b响应干旱胁迫、冷胁迫和盐胁迫,而对OsHsfC1b的研究表明其维持水稻耐旱性依赖于ABA调控途径,因此进一步猜测FaHsfC1b可能也具有相同功能[14]。另外研究指出生长素诱导型启动子往往同时含有干旱胁迫或其他胁迫相关的作用元件,表明启动子下游基因的调控依赖于激素和逆境胁迫的共同作用[41]。
目前除ABA以外,C类热激转录因子在其他植物外源激素处理下的表达分析尚无报道,因此猜测植物激素可能通过启动子来调控FaHsfC1b的表达,进而在植物抗逆性中发挥作用。由于此类激素尤其是生长素、细胞分裂素和赤霉素和植物生长密切相关,结合水稻中的OsHsfC1b在水稻生长中所扮演的不可缺少的角色[14],进一步猜测FaHsfC1b可能也在高羊茅的生长发育中发挥重要作用。
本研究通过对高羊茅FaHsfC1b启动子序列的分析,推测FaHsfC1b可能在植物激素介导的抗逆性中发挥重要作用,对于植物生长发育及正常的光合作用可能也是必不可少的。
参考文献:
[1]黄小云,陶 鹏,李必元,等. 植物热激转录因子基因家族的研究进展[J]. 浙江农业科学,2014(9):1323-1332,1336.
[2]Chen H,Hwang J E,Lim C J,et al. Arabidopsis DREB2C functions as a transcriptional activator of HsfA3 during the heat stress response[J]. Biochemical and Biophysical Research Communications,2010,401(2):238-244.
[3]Czarneckaverner E,Yuan C X,Fox P C,et al. Isolation and characterization of six heat shock transcription factor cDNA clones from soybean[J]. Plant Molecular Biology,1995,29(1):37-51.
[4]Yokotani N,Ichikawa T,Kondou Y,et al. Expression of rice heat stress transcription factor OsHsfA2e enhances tolerance to environmental stresses in transgenicArabidopsis[J]. Planta,2008,227(5):957-967.
[5]Bharti K,Koskull-Döring P V,Bharti S,et al. Tomato heat stress transcription factor HsfB1 represents a novel type of general transcription coactivator with a histone-like motif interacting with the plant CREB binding protein ortholog HAC1[J]. The Plant Cell,2004,16(6):1521-1535.
[6]Scharf K D,Berberich T,Ebersberger I,et al. The plant heat stress transcription factor (Hsf) family: structure,function and evolution[J]. Biochimica et Biophysica Acta,2012,1819(2):104-119.
[7]Hu W,Hu G,Han B. Genome-wide survey and expression profiling of heat shock proteins and heat shock factors revealed overlapped and stress specific response under abiotic stresses in rice[J]. Plant Science,2009,176(4):583-590.
[8]Xue G P,Sadat S,Drenth J,et al. The heat shock factor family fromTriticumaestivumin response to heat and other major abiotic stresses and their role in regulation of heat shock protein genes[J]. Journal of Experimental Botany,2014,65(2):539-557.
[9]Huang X Y,Tao P,Li B Y,Wang W H,et al. Genome-wide identification,classification,and analysis of heat shock transcription factor family in Chinese cabbage (Brassicarapapekinensis)[J]. Genetics and Molecular Research,2015,14(1):2189-2204.
[10]Lin Q,Jiang Q,Lin J,et al. Heat shock transcription factors expression during fruit development and under hot air stress in Ponkan (CitrusreticulataBlanco cv. Ponkan) fruit[J]. Gene,2015,559(2):129-136.
[11]Huang Y,Li M Y,Wang F,et al. Heat shock factors in carrot: genome-wide identification,classification,and expression profiles response to abiotic stress[J]. Molecular Biology Reports,2015,42(5):893-905.
[12]Li P S,Yu T F,He G H,et al. Genome-wide analysis of the Hsf family in soybean and functional identification of GmHsf-34 involvement in drought and heat stresses[J]. BMC Genomics,2014,15:998-1009.
[13]Zhang J,Li Y,Jia H X,et al. The heat shock factor gene family inSalixsuchowensis:a genome-wide survey and expression profiling during development and abiotic stresses[J]. Frontiers in Plant Science,2015,6:735-748.
[14]Schmidt R,Schippers J H,Welker A,et al. Transcription factor OsHsfC1b regulates salt tolerance and development inOryzasativassp.japonica[J]. AoB Plants,2012,2012(1):11-23.
[15]伊淑莹,孙爱清,赵春梅,等. 番茄多胁迫诱导型LeMTshsp启动子的分子克隆及其功能分析[J]. 云南植物研究,2007,29(2):223-230.
[16]Freeman J,Sparks C A,West J,et al. Temporal and spatial control of transgene expression using a heat-inducible promoter in transgenic wheat[J]. Plant Biotechnology Journal,2011,9(7):775-788.
[17]孙芳芳,宋洪元. 植物诱导型启动子研究进展[J]. 南方园艺,2014(2):51-56.
[18]Yamaguchi-Shinozaki K,Shinozaki K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters[J]. Trends in Plant Science,2005,10(2):88-94.
[19]Larkindale J,Huang B. Thermotolerance and antioxidant systems inAgrostisstolonifera:involvement of salicylic acid,abscisic acid,calcium,hydrogen peroxide,and ethylene[J]. Plant Physiology,2004,161(4):405-413.
[20]Liu N,Ko S,Yeh K C,et al. Isolation and characterization of tomatoHsa32 encoding a novel heat-shock protein[J]. Plant Science,2006,170(5):976-985.
[21]Larkindale J,Hall J D,Knight M R,et al. Heat stress phenotypes ofArabidopsismutants implicate multiple signaling pathways in the acquisition of thermotolerance[J]. Plant Physiology,2005,138(2):882-897.
[22]Jin G H,Gho H J,Jung K H. A systematic view of rice heat shock transcription factor family using phylogenomic analysis[J]. Journal of Plant Physiology,2013,170(3):321-329.
[23]Sakata T,Oshino T,Miura S,et al. Auxins reverse plant male sterility caused by high temperatures[J]. Proceedings of the National Academy of Sciences of the United States of America,2010,107(19):69-74.
[24]Sun T P,Gubler F. Molecular mechanism of gibberellin signaling in plants[J]. Annual Review of Plant Biology,2004,55(1):197-223.
[25]Fleet C M,Sun T P. A dellacate balance: the role of gibberellin in plant morphogenesis[J]. Current Opinion in Plant Biology,2005,8(1):77-85.
[26]Colebrook E H,Thomas S G,Phillips A L,et al. The role of gibberellin signalling in plant responses to abiotic stress[J]. Journal of Experimental Biology,2014,217(1):67-75.
[27]Branlard T. Proteomic analysis of the effect of heat stress on hexaploid wheat grain : characterization of heat-responsive proteins from non-prolamins[J]. Proteomics,2004,(4):505-513.
[28]Oshino T,Abiko M,Saito R,et al. Premature progression of anther early developmental programs accompanied by comprehensive alterations in transcription during high-temperature injury in barley plants[J]. Molecular Genetics and Genomics,2007,278(1):31-42.
[29]Vanstraelen M,Benková E. Hormonal interactions in the regulation of plant development[J]. Annual Review of Cell & Developmental Biology,2012,28(28):451-463.
[30]Hoagland D R,Arnon D I. The water-culture method for growing plants without soil[J]. Califagricexpstncirc,1937,347(5406):357-359.
[31]李广平,张长青,章 镇,等. 中国李pgip启动子的克隆及调控元件分析[J]. 园艺学报,2009,36(10):1425-1430.
[32]Altschul S F,Madden T L,Schäffer A A,et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs[J]. Nucleic Acids Research,1997,25(17):389-402.
[33]Lescot M,Déhais P,Thijs G,et al. PlantCARE,a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences[J]. Nucleic Acids Research,2002,30(1):325-327.
[34]Higo K,Ugawa Y,Iwamoto M,et al. Plant cis-acting regulatory DNA elements (PLACE) database[J]. Nucleic Acids Research,1999,27(1):297-300.
[35]Kilian J,Whitehead D,Horak J,et al. The AtGenExpress global stress expression data set: protocols,evaluation and model data analysis of UV‐B light,drought and cold stress responses[J]. Plant Journal,2007,50(2):347-363.
[36]Wang J,Sun N,Deng T,et al. Genome-wide cloning,identification,classification and functional analysis of cotton heat shock transcription factors in cotton (Gossypiumhirsutum)[J]. BMC Genomics,2014,15:955-961.
[37]Reddy P S,Kavi Kishor P B,Seiler C,et al. Unraveling regulation of the small heat shock proteins by the heat shock factor HvHsfB2c in barley: its implications in drought stress response and seed development[J]. PloS One,2014,9(3):119-125.
[38]Udo Gowik J B,Meryem A,Ute S,et al. cis-Regulatory elements for Mesophyll-Specific gene expression in the C4plant flaveria trinervia,the promoter of the C4phosphoenolpyruvate carboxylase gene[J]. The Plant Cell,2004,16(5):1077-1090.
[39]Bechtold U,Albihlal W S,Lawson T,et al.ArabidopsisHEAT SHOCK TRANSCRIPTION FACTORA1b overexpression enhances water productivity,resistance to drought,and infection[J]. Journal of Experimental Botany,2013,64(11):3467-3481.
[40]Nishiuchi T,Shinshi H,Suzuki K. Rapid and transient activation of transcription of theERF3 gene by wounding in tobacco leaves: possible involvement of NtWRKYs and autorepression[J]. Journal of Biological Chemistry,2005,279(53):355-361.
[41]Liu Z B,Hagen G,Guilfoyle T J. A G-Box-Binding protein from soybean binds to the E1 auxin-response element in the soybean GH3 promoter and contains a proline-rich repression domain[J],Plant Physiology,1997,115(2):397-407.
猜你喜欢
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06
金属加工(热加工)(2020年12期)2020-02-06
山西农经(2019年5期)2019-02-22
腹腔镜外科杂志(2016年11期)2016-06-01
现代工业经济和信息化(2016年2期)2016-05-17
磁共振成像(2015年9期)2015-12-26
实用手外科杂志(2015年2期)2015-08-28
电子工业专用设备(2015年4期)2015-05-26
无机化学学报(2014年3期)2014-02-28
作物研究(2012年7期)2012-11-06
