
混养鲮对凡纳滨对虾养殖池塘浮游生物群落结构的影响
2018-03-14 11:24李志斐李家磊王金林余德光王广军
河南农业科学 2018年1期
李志斐,李家磊,王金林,谢 骏,余德光,王广军
(中国水产科学研究院珠江水产研究所/农业部热带亚热带水产资源利用与养殖重点实验室,广东广州510380)
凡纳滨对虾(Litopenaeus vannamei)俗称南美白对虾,是当前世界产量最高的甲壳类养殖品种之一。2015年,我国凡纳滨对虾养殖总产量达 73.15 万 t[1]。凡纳滨对虾适盐度广,可在咸淡水和淡水地区养殖,加之生长迅速、出肉率高,在沿海和内陆地区养殖广泛[2]。然而在对虾精养池塘中,高投饵量造成了大量的残余饵料和代谢物质的沉积,池塘中的有机污染严重[3-4]。因养殖水体的富营养化,进而导致对虾直接死亡或者诱发疾病,给养虾业带来了严重的经济损失,严重阻碍了对虾产业的发展[5-8]。
研究发现,对虾与其他经济品种的混养可有效缓解池塘污染[9-10]。鱼虾混养模式是通过在虾类养殖水体中放养一定数量和密度的鱼类,使养殖水体符合虾类生长条件,又可让鱼类健康生长。Yuan等[11]在凡纳滨对虾养殖池塘中混养不同密度和不同规格的罗非鱼(Oreochromis mossambicus),研究混养罗非鱼对对虾养殖池塘水质、产量和营养利用效率的影响,提出了对虾与罗非鱼混养可以作为实现对虾养殖可持续发展的一种养殖模式。孔谦[12]将凡纳滨对虾与鲻(Mugil cephalus)进行混养发现,在养殖的中后期,混养条件能显著提高养殖水体中的浮游植物数量和多样性,致病菌数量也显著低于对虾单养,并且混养模式较单养模式经济效益提高15.2%。此外,还有点带石斑鱼(Epinephelus coioides)[13]和鲢 (Hypophthalmichthys molitrix)、鳙(Aristichthys nobilis)[14]等与虾混养的模式,都带来了可观的经济效益。但以上混养的鱼类多为池塘中上或中下层鱼类,对于混养底层刮食性鱼类的研究并不多见。
对虾养殖的残饵和粪便会在养殖水体中不断累积[15],底层刮食性鱼类可以直接通过它们的摄食活动,清除池塘中的残饵及有机质,提高饵料利用率,改善池塘环境条件;也可通过它们的摄食活动,起到搅动底层池水[16]、改变养殖池塘底部氧化还原电位、促进养殖池塘底层物质循环及能量流通的作用,对养殖动物生长和产量的提高均能起到积极作用[17]。鲮(Cirrhinus molitorella)作为底栖刮食性鱼类,长期生活在池塘底部,以池塘底部营养物质为食,可以减少池塘营养物质积累,形成稳定的生态食物链,被称为池塘的“清道夫”。鉴于此,将鲮应用于凡纳滨对虾养殖池塘中,以期改善对虾池塘养殖环境,同时通过测定混养前后池塘水质、浮游生物群落等评价混养对养殖池塘生态的影响,旨在构建一种复合型养殖模式。
1 材料和方法
1.1 试验地点及设计
试验在广东省江门市某养殖场进行。采样池塘2 口,面积约 0.266 hm2,长宽比约为 7 ∶4,南北朝向。2口池塘前期2个月单养凡纳滨对虾,单养2个月后开始混养鲮。采样期间池塘的平均水位为2 m。单养凡纳滨对虾第45天时采样作为单养样品,放养鲮后在第15天时采样作为混养样品。每口池塘采用五点采样法,即池塘中心和四角5个点采样,然后混合为1个样,每个点重复3次。
每口池塘每0.067 hm2放养凡纳滨对虾5万尾,混养鲮水花20万尾,整个试验过程只投喂凡纳滨对虾商品饲料,投喂量约为对虾总质量的2%~3%,以2 h内摄食量酌情增减投喂量,投喂时间为8:00—10:00 和 16:00—18:00。
1.2 测定指标
水体温度、溶解氧、电导率、pH值、溶解性固体总量等指标采用YSI多功能水质分析仪现场测定,水体总氮、总磷、硝酸盐、亚硝酸盐、磷酸盐和总有机碳含量等指标按照标准方法测定[11]。凡纳滨对虾上市时,统计对虾及鲮产量。
浮游动物样品分定性和定量采集,定性采集使用25号浮游生物网捞取,加2%福尔马林固定后带回实验室,参照《淡水浮游生物图谱》进行种类鉴定[18];定量样品采集20 L水样,经25号浮游生物网滤缩后放入小塑料瓶中,加福尔马林固定后鉴定[19]。浮游植物样品定量采集,取1 L水样加入鲁哥氏液固定,然后倒入有刻度的沉淀器定容,静置24 h后,用虹吸管吸取上清液,浓缩至30 mL的塑料瓶中再进行鉴定。
根据浮游动植物不同物种的出现频率(fi)和个体数量(Ni)与总数量(N)的比例来计算优势度,公式为:优势度Y=(Ni/N)fi。当 Y≥0.02时,确定为优势种[19]。
采用Shannon-Weiner多样性指数(H)计算混养前、后多样性的变化,计算公式为[20]:
式中,s为浮游动/植物的种类数,Pi为第i种浮游动/植物所占总数的比例。
1.3 数据分析
数据用SPSS 16.0统计软件进行统计分析,Oneway ANOVA分析差异显著性,结果以平均值±标准误表示。
2 结果与分析
2.1 混养对养殖池塘水质的影响
由表1可知,混养后,养殖水体电导率、溶解性固体总量、总氮、氨氮、硝酸盐、亚硝酸盐含量显著降低(P<0.05),磷酸盐含量显著升高(P<0.05),水体温度、溶解氧、总有机碳含量和pH值无显著变化(P>0.05),说明混养可有效降低养殖水体中有直接毒害作用的氨氮及亚硝酸盐含量。

表1 混养前、后养殖水体理化指标变化
2.2 混养对凡纳滨对虾生长的影响
凡纳滨对虾上市时,平均每0.067 hm2池塘产虾400 kg、鲮30 kg,单养凡纳滨对虾精养池塘产量一般为每0.067 hm2产虾300 kg。此外,混养的鲮也可作为饵料鱼出售,养殖效益增加。混养条件下,凡纳滨对虾饵料系数在0.6~0.8,相比精养虾塘的饵料系数(一般在1.2~1.5),具有明显的成本优势。
2.3 混养对养殖池塘浮游植物的影响
2.3.1 浮游植物的种类组成 经鉴定,2口池塘混养前浮游植物共6门46种,混养后共6门44种(表2)。其中,混养前绿藻门占 50.00%,蓝藻门占19.56%;混 养 后 绿 藻 门 占 54.54%,蓝 藻 门 占22.73%。对虾池塘混养后的浮游植物种类数少于混养前。

表2 混养前、后养殖水体浮游植物各门种数及其比例
2.3.2 浮游植物优势种 对虾池塘中以绿藻门(Chlorophyta)为主要优势类群,以优势度Y≥0.02为优势种的标准,优势种有蓝藻门的色球藻(Chroococcus sp.)、优美平裂藻(Merismopedia elegans)、细小平裂藻(Merismopedia tenuissima)、断裂颤藻(Oscillatoria fraca),绿藻门的二形栅藻(Scenedesmus dimorphus)、四尾栅藻(Scenedesmus quadricauda)、镰形纤维藻(Ankistrodesmus falcatus)、微小新月藻(Tetraedron minimum)、小空星藻 (Coelastrum microporum),硅藻门的小环藻(Cyclotella sp.),隐藻门的尖尾蓝隐藻(Chroomon asacuta)、倒卵隐藻(Cryptomonas obovata)、马索隐藻(Cryptomonas marssonii),裸藻门的扁裸藻(Phacus sp.)等(表3)。混养后新增了盐泽螺旋藻(Spirulina subsalsa)、斜生栅藻(Scenedesmusobliquus)、球 衣 藻 (Chlamydomonas globosa)、三叶四角藻(Tetraedron tumidulum)、微小四角藻(Tetraedron minimum)、克里藻(Klebsormidium sp.),减少了卷曲鱼腥藻(Anabaena circnalis)、狭形纤维藻(Ankistrodesmus angustus)、单角盘星藻(Pediastrum simplex)、卵囊藻(Oocystis sp.)、平片针杆藻(Synedra tabulata)、剑尾陀螺藻(Strombomonas ensifera)。
2.3.3 浮游植物的密度和生物量 混养前、后对虾池塘浮游植物密度和生物量变化见图1。对虾池塘混养前浮游植物的密度和生物量分别为167.32×106个/L和7.84 mg/L,混养后浮游植物的密度和生物量分别为 53.69×106个/L和 4.24 mg/L。混养前、后2口池塘均以蓝藻门密度最高,混养前为146.81×106个/L,混养后为 36.99×106个/L;而混养前、后生物量均以硅藻门最高,混养前为3.59 mg/L,混养后为 1.70 mg/L。

表3 混养前、后养殖水体浮游植物种类组成变化
2.3.4 浮游植物的生物多样性 图2显示对虾池塘混养前、后浮游植物群落的多样性指数变化,混养前浮游植物的Shannon-Weiner多样性指数是1.42,混养后Shannon-Weiner多样性指数是1.76。对虾池塘混养后浮游植物Shannon-Weiner多样性指数升高。
2.4 混养对养殖池塘浮游动物的影响
2.4.1 浮游动物的种类组成 经鉴定,对虾池塘混养前浮游动物共4门23种,混养后池塘中浮游动物共4门17种,其中轮虫的比例从43.48%增加到47.06%,而枝角类和桡足类比例分别从13.04%降低到11.76%,26.09%降低到 23.53%(表 4)。
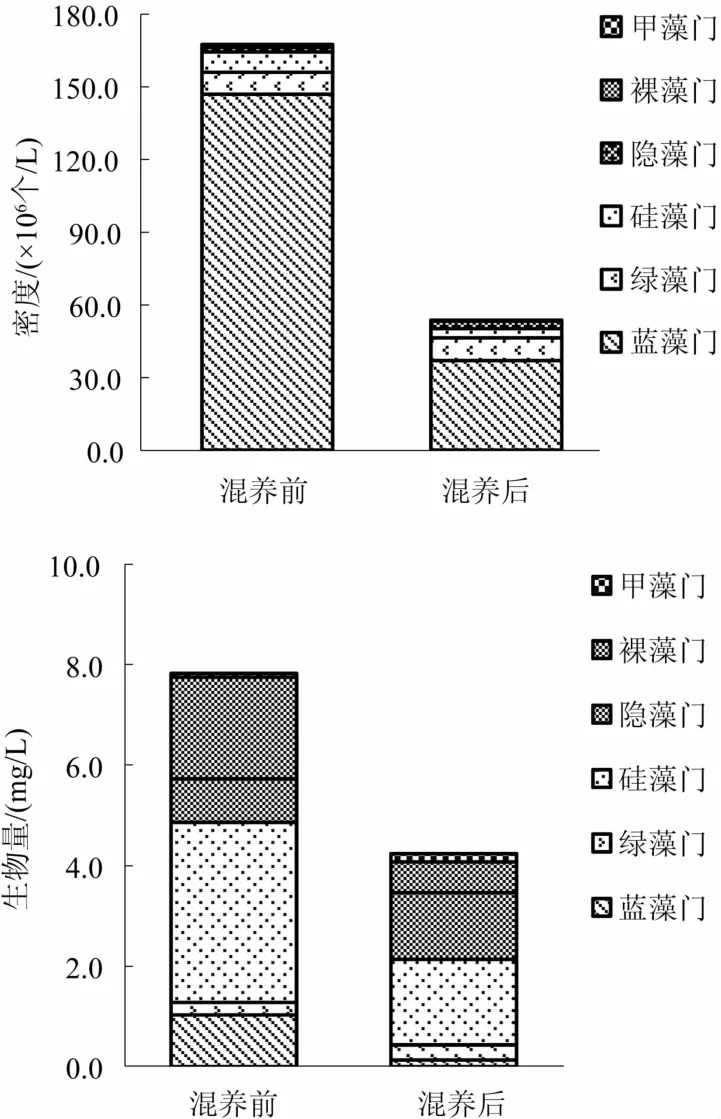
图1 混养前、后养殖水体浮游植物密度和生物量变化
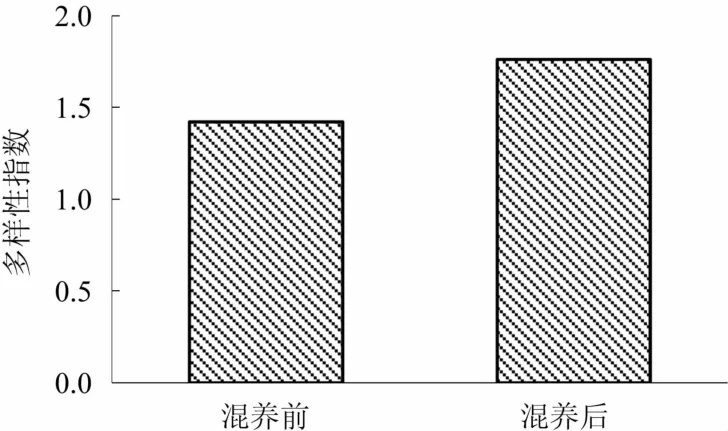
图2 混养前、后养殖水体浮游植物群落多样性指数变化
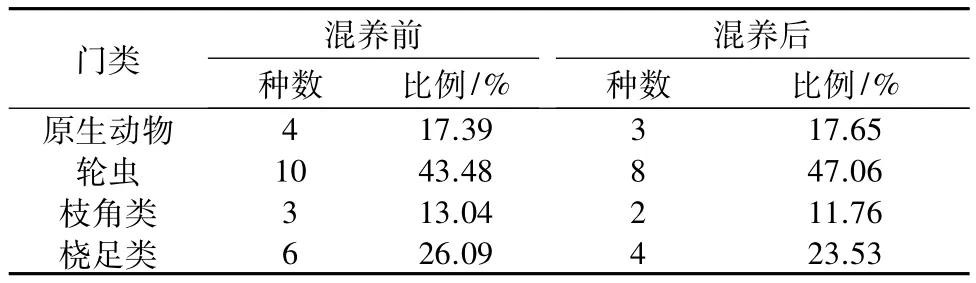
表4 混养前、后养殖水体浮游动物各门种数及其比例
2.4.2 浮游动物优势种 表5显示了混养前、后浮游动物种类组成变化,对虾池塘中的优势类群为轮虫,亚优势类群为桡足类;其中2口池塘混养前、后共有的优势种有纤毛虫(Ciliophora sp.)、暗小异尾轮虫(Trichocerca pusilla)和剑水蚤桡足幼体。对虾池塘混养后增加的优势种有单环栉毛虫(Didinium balbianii)、螺形龟甲轮虫(Keratella cochlearis)、长三肢轮虫(Filinia longiseta)、方形网纹溞(Ceriodaphnia quadrangula)和广布中剑水蚤(Mesocyclops leuckarti)。

表5 混养前、后养殖水体浮游动物种类组成变化
2.4.3 浮游动物的密度和生物量 图3显示了对虾池塘混养前、后浮游动物生物量和密度变化。混养前池塘浮游动物生物量和密度分别是3.981 mg/L和3 014.6 ind./L,混养后的浮游动物生物量和密度分别是 0.968 mg/L 和 1 201.65 ind./L,混养后浮游动物密度和生物量均低于混养前。对虾池塘混养前、后密度和生物量最高的浮游动物均为轮虫。
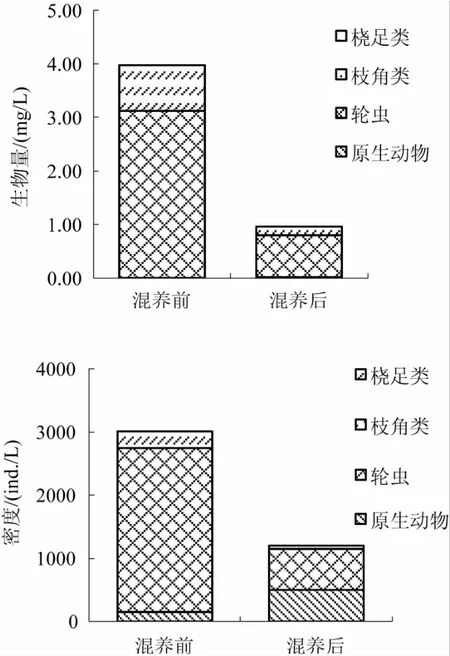
图3 混养前、后养殖水体浮游动物生物量和密度变化
2.4.4 浮游动物的生物多样性 图4显示了对虾池塘混养前、后浮游动物群落多样性指数变化。对虾池塘混养前的 Shannon-Weiner多样性指数为1.36,混养后的 Shannon-Weiner多样性指数为1.26,混养后的Shannon-Weiner多样性指数降低。
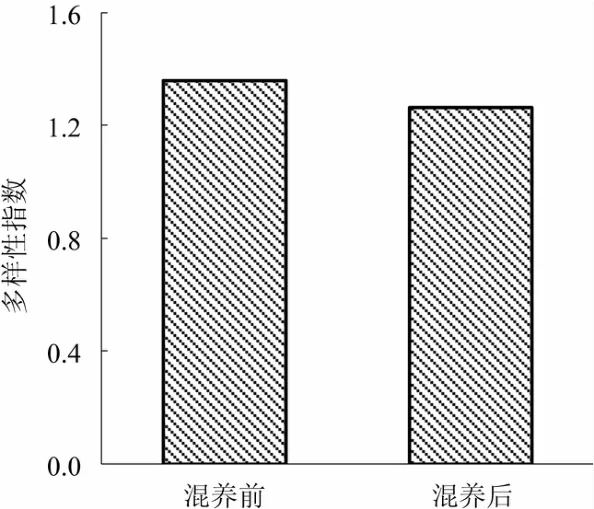
图4 混养前、后养殖水体浮游动物群落多样性指数变化
3 结论与讨论
3.1 混养模式下养殖池塘中水体及对虾生长的变化
前人研究表明,浮游植物的密度、生物量与水体中氮、磷的含量呈正相关[21]。其原因可能是浮游植物容易受到水体中营养盐的影响。在其他影响因子相同的情况下,一定范围内,氮和磷的含量越高,养殖水体中浮游植物的种类和丰度就越高[22]。本试验中,混养后,养殖池塘水体中总氮的含量降低,浮游植物的多样性增加,物种数减少,在一定程度上控制了水体富营养化[23-24]。
Burford等[25]认为,从饵料和对虾粪便中释放到水体中的氮主要以有机氮的形式存在,这些有机氮的生物利用率低,在养殖池塘中逐渐积累,可能影响对虾养殖池塘的水质情况,促进有害物质的生长。本试验中单养模式下养殖水体亦是如此。在混养鲮之后,鲮对残饵和粪便的摄取有效地降低了养殖水体中氮的含量,从而改善对虾生长环境,为对虾产量的提高创造了极有利的条件。
水产动物的生长受到生长环境的影响,高产池塘养殖环境下鱼虾的生长受到池塘水质的影响,本试验中混养鲮后的凡纳滨对虾池塘水质得到明显改善,同时生长数据表明,凡纳滨对虾产量有了较大提升,表明池塘混养生态型模式的构建有利于鱼虾的生长,更有助于养殖池塘产量的提升。
3.2 混养模式下养殖池塘中浮游植物的变化
通常情况下,浮游植物中的硅藻门量多被认定为水质污染较轻的典型特征,而蓝藻门量多则意味着水体富营养化程度较高[26]。本试验结果表明,浮游植物的优势种为绿藻门,但蓝藻门的密度较大,硅藻门的生物量较高。试验期间,对虾池塘混养前与混养后的浮游植物的密度和生物量相差较大,其原因可能是2口池塘中的浮游植物群落组成及结构易受外界环境因子的影响,如水温、营养盐、动物(主要包括养殖经济动物及浮游动物)的摄食和降雨等[27]。浮游植物多样性研究具有独特的生物学特征,在浮游生物群落生态学研究中发挥越来越重要的作用。浮游植物多样性也是浮游生物群落结构和功能完整性评价的重要指标之一[28]。本试验中,混养后的Shannon-Weiner多样性指数高于混养前的Shannon-Weiner多样性指数,表明在凡纳滨对虾池塘中混养鲮的方式,可以有效提高池塘中浮游植物的多样性,有利于凡纳滨对虾的健康和快速生长。
3.3 混养模式下养殖池塘中浮游动物的变化
由优势种可以看出,小单环栉毛虫、长三肢轮虫、广布多肢轮虫和角突臂尾轮虫均为富营养水体的常见指示物种,均具有较强的耐污能力[29-30]。此外,随着水体富营养化程度的增加,浮游动物的密度和生物量均逐渐增加,其中小型浮游动物原生动物和轮虫的增长尤为迅速,浮游动物往往还表现出小型化趋势[31-32]。本试验中,上述的浮游动物的种类和数量在混养前后没有明显变化,可能和虾苗前期的摄食以及混养后鲮水花对浮游动物的摄食有关。
试验期间,对虾养殖池塘混养前、后浮游动物的密度和生物量相差较大,混养后浮游动物的生物量和密度均低于混养前,这可能与混养鲮后池塘氮水平降低,富营养化程度缓解,以及鲮大量捕食浮游动物有关。
魏小岚等[33]分析得出,凡纳滨对虾循环水养殖水体中浮游动物的多样性指数为0.76~2.08;本试验中,对虾池塘混养前、后的浮游动物 Shannon-Weiner多样性指数分别为1.36和1.26。浮游动物物种多样性易受到多种因素(水温、空间、时间、竞争和捕食、水体条件、初级生产力等)的影响[29],对虾池塘浮游动物Shannon-Weiner多样性指数混养后低于混养前,可能与水体水质条件以及鱼类捕食有关。
综上,混养后养殖水体总氮、氨氮含量等水质指标显著降低,磷酸盐含量升高,一方面减少了氨氮等有毒含氮化合物对凡纳滨对虾产生的直接毒害作用,另一方面,降低了水体氮磷比,对平衡池塘营养物质起到一定促进作用;其次,混养后养殖水体浮游植物和浮游动物密度及生物量均降低,且混养初级生产力浮游植物多样性提高,次级生产力浮游动物多样性降低,说明混养鲮可充分利用水体中的各种天然饵料,有利于养殖水体的藻相平衡。凡纳滨对虾与鲮混养可作为一种生态养殖模式。
[1] 农业部渔业渔政管理局.中国渔业年鉴[M].北京:中国农业出版社,2016.
[2] 尤宏争,武宝岭,郑艳坤,等.海水池塘设施化养虾技术初步研究[J].天津农业科学,2013,19(10):7-9.
[3] 杨世平,邱德全.对虾高密度养殖过程中水质的周期变化与分析[J].水产科学,2006,25(9):459-462.
[4] 陈东兴,杨超,华雪铭,等.3种虾类养殖池塘污染强度及氮磷营养物质收支研究[J].河南农业科学,2013,42(8):132-136.
[5] Martínezporchas M,Martínezcórdova L R,Porchascornejo M A,et al.Shrimp polyculture:A potentially profitable,sustainable,but uncommon aquacultural practice[J].Reviews in Aquaculture,2010,2:73-85.
[6] 郑辉,许文超.4种海藻在南美白对虾养殖水体中的生态作用[J].河南农业科学,2016,45(5):144-147.
[7] 王仁杰,姜令绪,李玉全.内外因素对凡纳滨对虾仔虾同类相残的影响[J].河南农业科学,2015,44(2):142-145.
[8] 朱林,车轩.有益微生物在南美白对虾养殖中的应用研究[J].山西农业科学,2013,41(11):1291-1293.
[9] Henry-Silva G G,Maia C S P,Moura R S T,et al.Integrated multi-trophic culture of Nile tilapia(Oreochromis niloticus)and Amazon river prawn(Macrobrachium amazonicum) in brackish water[J].Arquivo Brasileiro de Medicina Veterináriae Zootecnia,2015,67(1):265-273.
[10] Apún-Molina J P,Santamaría-Miranda A,Luna-González A,et al.Growth and metabolic responses of white leg shrimp Litopenaeus vannamei and Nile tilapia Oreochromis niloticus in polyculture fed with potential probiotic microorganisms on different schedules[J].Latin American Journal of Aquatic Research,2015,43(3):435-445.
[11] Yuan D,Yi Y,Yakupitiyage A,et al.Effects of addition of red tilapia(Oreochromis spp.)at different densities and sizes on production,water quality and nutrient recovery of intensive culture of white shrimp(Litopenaeus vannamei)in cement tanks[J].Aquaculture,2010,298(3):226-238.
[12] 孔谦.凡纳滨对虾与鲻鱼混养中精养池的理化生物因子的研究[D].广州:广东海洋大学,2010.
[13] 张嘉晨,刘建勇,袁瑞鹏,等.凡纳滨对虾与点带石斑鱼的混养模式[J].广东海洋大学学报,2015,35(4):46-50.
[14] 王吉桥,罗鸣,马成学,等.低盐水体南美白对虾与鲢鳙鱼混养的试验[J].水产科学,2003,22(6):21-24.
[15] 虞为,李卓佳,朱长波,等.我国对虾生态养殖的发展现状、存在问题与对策[J].广东农业科学,2011,38(17):168-171.
[16] Avnimelech Y,Kochva M,Hargreaves J A.Sedimentation and resuspension in earthen fish ponds[J].Journal of the World Aquaculture Society,2000,30(4):401-409.
[17] Tendencia E A,Pea M R D,Choresca C H.Effect of shrimp biomass and feeding on the anti-Vibrio harveyi,activity of Tilapia sp.in a simulated shrimp-tilapia polyculture system[J].Aquaculture,2006,253(1/2/3/4):154-162.
[18] 韩茂森.淡水浮游生物图谱[M].北京:农业出版社,1980.
[19] 邓道贵,杨威,孟小丽,等.淮河中游浮游甲壳动物群落结构的季节动态[J].水生生物学报,2013,37(5):869-875.
[20] 刘健康.高级水生生物学[M].北京:科学出版社,1999:78.
[21] 郑宗林,郑曙明,王广军,等.河蚌对草鱼混养精养池塘中浮游植物群落结构的影响[J].水产科学,2015,34(12):757-761.
[22] Hanno T,Bernhard M,Drte B,et al.Substrate-controlled succession of marine backterioplankton populations induced by a phytoplankton bloom [J].Science,2012,336(6081):608-611.
[23] Domis L N,Van de Wall D B,Helmsing N R,et al.Community stoichiometry in a changing world:Combined effects of warming and eutrophication on phytoplankton dynamics[J].Ecologgy,2014,95(6):1485-1495.
[24] Stephanie H,Richard L,David J.Variability in phytoplankton community structure in response to North Atlantic Oscillation and implications for organic carbon flux[J].Limnology and Oceanography,2012,57(6):1591-1601.
[25] Burford M A,Williams K C.The fate of nitrogenous waste from shrimp feeding[J].Aquaculture,2001,198(12):79-83.
[26] 邓文丽,刘均平,王晓星,等.北京野鸭湖浮游植物群落结构与水质关系研究[J].湿地科学,2013,11(1):27-34.
[27] 胡梦红,武震,周作强,等.鱼蚌混养对池塘水质、藻相结构及三角帆蚌生长的影响[J].水产学报,2014,38(2):200-207.
[28] 孙儒泳.动物生态学原理[M].3版.北京:北京师范大学出版社,2002:177.
[29] 沈韫芬,顾曼如,龚循矩,等.微型生物监测新技术[M].北京:中国建筑工业出版社,1990:152-175.
[31] 钱方平,席贻龙,温新利,等.湖泊富营养化对轮虫群落结构及物种多样性的影响[J].生物多样性,2007,15(4):344-355.
[32] 杨宇峰,黄祥飞.武汉东湖浮游动物群落结构的研究[J].应用生态学报,1994,5(3):319-324.
[33] 魏小岚,李纯厚,颉晓勇,等.对虾高位池循环水养殖水体浮游植物生态特征研究[J].农业环境科学学报,2014,33(2):392-403.
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年5期)2022-06-05
当代水产(2021年8期)2021-11-04
海洋通报(2021年3期)2021-08-14
当代水产(2021年4期)2021-07-20
科学养鱼(2020年10期)2020-11-23
课外语文·中(2020年2期)2020-03-24
课外语文(2020年5期)2020-03-14
当代水产(2019年2期)2019-05-16
当代水产(2019年1期)2019-05-16
