
河南宝天曼珍稀园林树种陕西紫茎的群落特征
2018-03-14 11:24朱学灵程蓓蕾任应党
河南农业科学 2018年1期
朱学灵,杨 艳,程蓓蕾,任应党
(1.河南省昊泽苑绿化工程有限公司,河南平顶山467300;2.河南省农业科学院,河南 郑州450002)
紫茎属(Sewartial L.)是一类东亚-北美间断分布的山茶科(Theaceae)植物。我国的南部和西南部,既是紫茎属的起源中心,又是紫茎属的高度分化中心。全世界紫茎属植物共23种5变种,中国有17种,且均为我国的特有属植物,这些植物分布于广西、云南、广东、湖南等地[1]。陕西紫茎(Stewartia shensiensis Chang)分布于北纬33°附近,是紫茎属十几种植物中在中国大陆最北分布的一个种类[2]。河南宝天曼自然保护区的平坊、猴沟和七里沟林区的沟谷地带,分布有片状的陕西紫茎群落,且生长良好,是河南省陕西紫茎群落分布最为集中的地区之一。
陕西紫茎因树皮光滑,伏牛山区俗称其名为“猴不上”。其外皮淡黄灰色,呈纸层剥落,内皮黄绿色,花白色,于6月中旬开花,弥补了河南园林植物中晚春春花不足的缺憾,是一个优良的园林绿化树种。前人进行了紫茎属植物的分类学和地理分布研究[1],而对于陕西紫茎群落结构、物种分布机制、园林引种设计等方面研究得较少。目前,河南省尚未进行陕西紫茎的园林引种驯化工作,更没有在园林造景上使用过这种珍稀野生物种。为了揭示陕西紫茎群落的稳定性和物种共存机制,对宝天曼自然保护区陕西紫茎群落特征和景观应用方面开展研究,为我国中部地区进行陕西紫茎的引种和园林景观绿化提供科学依据。
1 材料和方法
1.1 研究地点概况
宝天曼自然保护区位于河南省内乡县,地处秦岭东段伏牛山南坡,地理位置为东经111°53'~112°00',北纬 33°25'~33°33',是陕西紫茎群落在我国的最北分布区。平均海拔800~1 840 m;气候为暖温带大陆性季风气候,年平均气温15.1℃,1月平均气温1.5 ℃,7月平均气温27.8 ℃,无霜期210~220 d;年平均降水量885.6 mm,年平均相对湿度68%;主要土壤类型为山地黄棕壤、棕壤等。保护区内植被类型多样,植物区系组成丰富[3-4],珍稀濒危植物种类较多[5],但植被类型除栎类等为大面积分布外,多数植物种类均为零星或小片状分布。
1.2 调查方法
调查地点设在宝天曼保护区的平坊林区。群落调查采用样地法,面积20 m×20 m,把样地均匀划分成16个5 m×5 m的相邻样方,样地调查以小样方为单位进行。乔木和灌木共调查16个小样方,采用逐株调查的方式;在每个小样方内,随机布设1个面积为2 m×2 m草本样方,和乔灌木一并进行调查。乔木小样方主要调查记录该样方内的植物种类、树高、冠幅和生活型;灌木小样方主要调查该样方内的植物种类、株数、多度、平均高度、盖度和生活型,乔木幼苗记入灌木;草本小样方主要调查该样方内的植物种类、株数(无性系植株每一克隆株为1个个体)、多度、平均高度、盖度和生活型[6]等指标。植物种类依据《河南植物志》[7]等有关物种的形态描述进行鉴定。
1.3 数据分析和处理
1.3.1 植物区系成分 根据已经鉴定的植物种类,确定样地植物在物种系统分类中的科属种。依据群落样地内物种所属的植物科属类别,对照《中国植物区系属的分布区类型》[8],确定陕西紫茎群落的植物区系地理成分。
1.3.2 陕西紫茎群落的叶级谱 对样地内的每个物种采集生长健康的大、中、小新鲜叶片各10片,带回实验室在105℃烘箱中杀青2 h左右。杀青后用扫描仪将每一个叶片进行扫描处理,输入电脑,然后用Photoshop 7.0软件中的面积求算功能,计算出样地中研究对象的平均叶片面积,根据Paijmans的分类标准[9],划分出样地植物的叶级谱。
1.3.3 物种生活型谱 根据Raunkaier的生活型系统[9],统计不同植物物种的生活型,将不同群落的植物分为高位芽植物、地上芽植物、地面芽植物、地下芽植物和1年生植物。以生活型谱表示群落的种群结构,生活型谱计算如下:某一生活型的百分率=该群落中生活型的种类/该群落中的全部种类×100%。
1.3.4 植物重要值(IV)的计算 乔木层IV=(相对密度+相对频度+相对显著度)/3;灌木和草本层IV=(相对密度+相对频度+相对盖度)/3。密度用样地内每一个物种每平方米内的株数来表示,盖度用每一个物种在样地内的投影面积与样地总面积的比值来表示,频度用每一个物种在所有样方内出现的概率来表示。相对显著度以某植物的高度与整个群落植物高度和的比值来表示。相对密度、相对盖度和相对频度用每一个物种的密度、盖度和频度与所有物种的密度和、盖度和及频度和比值来表示。
1.3.5 群落物种多样性的测定 物种多样性主要是反映一个地区(或地段)内物种数的多寡和种间个体数的平均分配程度。物种丰富度是指群落中物种的丰富程度。为了避免不同物种因个体大小和个体数差异悬殊而导致多样性测度结果的差异,在此采用群落物种的IV作为多样性的测度指标。在进行群落特征分析的基础上,采用以下4种多样性指数来测定陕西紫茎群落的物种多样性。
(1)丰富度指数Rr:

式中,S为样地内的物种数。
(2)Shannon-Wiener多样性指数(H')[10]:
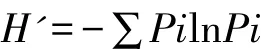
(3)Simpson 优势度指数(D)[11]

(4)Pielou 均匀度指数(J)[12]

上述(2)、(3)、(4)式中,Pi=Ni/N,Pi为该层次第i个物种相对重要值;Ni为样地中某一层次第i个物种的重要值,N为该层次所有物种重要值之和。1.3.6 陕西紫茎的园林群落设计 根据陕西紫茎的生物学特点、伴生树种和观赏特性,进行景观营造设计,确定该珍稀树种在园林上的应用途径。
2 结果与分析
2.1 陕西紫茎群落植物区系结构分析
2.1.1 区系组成 根据样地调查统计,共发现维管束植物37科59属77种。其中蕨类植物2科2属2种;种子植物35科57属75种。属种数量占优势的科为槭树科(Aceraceae,1属7种)、卫矛科(Celastraceae,2属5种)、蔷薇科(Rosaceae,5 属 5 种)、虎耳草科(Saxifragaceae,5属5种)、毛茛科(Ranunculaceae,4属 5种)、樟科(Lauraceae,1 属 4种);其他如山茶科、壳斗科(Fagaceae)、昆栏树科(Trochodendraceae)等18种植物均为1属1种,这些单属种植物占整个样地植物种类的48.6%。说明陕西紫茎群落物种构成复杂,陕西紫茎与槭树科、樟科、卫矛科等植物类群有较强的联结性。在群落水平上,宝天曼保护区陕西紫茎林物种多样性远大于浙江古田山自然保护区的紫茎群落,在群落植物的科属构成方面,与古田山自然保护区紫茎群落的构成类似[13]。2.1.2 地理成分 根据吴征镒的《中国植物区系属的分布区类型》[8],宝天曼自然保护区陕西紫茎群落内共有11个分布区类型及变型(表1),其中北温带分布的类型20属,占总属数的33.9%;东亚-北美间断分布的属为8属,占13.5%;泛热带和东亚分布的属均为7属,占11.9%;世界分布的属为6属,占10.1%。世界分布属的植物不参与热带、温带植物区系大类性质的统计,就属的地理成分性质来看,样地内属于热带属性质分布的类型10属,占17.0%,温带属性质分布的类型43属,占72.9%。这与宝天曼保护区处于我国北亚热带暖温带过渡区的地理位置有很大关系,也与保护区内杂木林、锐齿栎林等群落的植物区系特征相吻合[14],说明了宝天曼保护区植物与温带植物区系的亲缘关系。
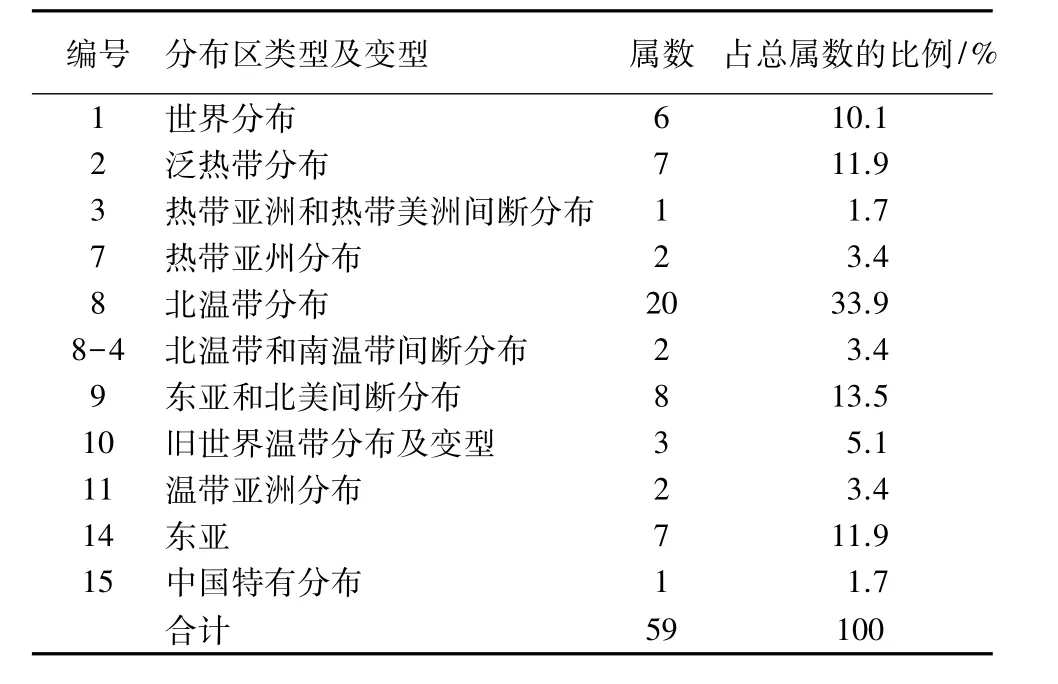
表1 宝天曼自然保护区陕西紫茎群落植物区系属的分布区类型
2.2 陕西紫茎群落外貌
2.2.1 季相 宝天曼自然保护区陕西紫茎林具有鲜明的季相。3月下旬冬芽萌动,4月中旬开始展叶;6月中旬陕西紫茎开花;10月中旬进入果熟期;10月下旬进入变叶期;11月上旬开始落叶。春季群落外貌呈淡绿色;夏季群落则变为浓绿色;深秋树叶凋零,陕西紫茎树干颜色特别醒目;入冬树叶落尽,整个群落季相呈黄褐色,林地枯枝落叶层较厚,林冠层透光度变大。
2.2.2 叶的性质 根据 Paijmans的分类标准[9],陕西紫茎群落的叶级谱中,以中型叶为主,共计68种,占总数的 87.17%;小型叶 4 种,占 5.13%;微型叶和大型叶各3种,占总数的3.85%。叶型以单型叶为主,叶质中草质叶占绝对优势,为88.31%(表2),基本上反映了暖温带落叶阔叶林的叶级谱特征。

表2 陕西紫茎林叶的性质
2.2.3 生活型 根据 Raunkaier[9]的生活型分类系统,确定陕西紫茎林的生活型谱。从表3可以看出,群落中,高位芽植物种类最多,有47种,占整个植物生活型谱的61.0%;地面芽植物次之,有14种,占整个植物生活型谱的18.2%;地上芽和1年生植物均为7种,占生活型谱的9.1%;地下芽植物种类最少,有2种,占生活型谱的2.6%。陕西紫茎群落以高位芽植物占优势,地面芽植物次之,与我国温带地区的落叶阔叶林相近,远低于亚热带常绿阔叶林和亚热带次生常绿落叶阔叶混交林,反映出陕西紫茎植物群落与气候之间的关系。
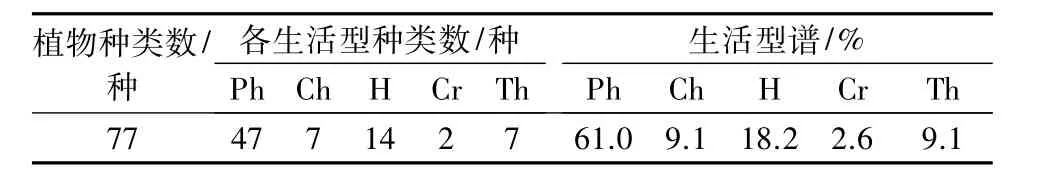
表3 宝天曼自然保护区陕西紫茎群落生活型谱
2.2.4 群落的结构特征 乔木层共出现种子植物15科15属19种,其中种属数量上处于优势的科为槭树科(1属4种),其余的均为1属1种。乔木层盖度约为80%,可分为2个亚层。其中第1亚层高13.0~18.0 m,主要组成种类为锐齿栎、白玉兰、山樱桃;第 2 亚层高 3.5~8.0 m,优势种类主要有陕西紫茎、千金榆、三亚乌药。从乔木层物种重要值(表4)来看,乔木层优势种为陕西紫茎、千金榆、青楷槭。重要值小于4的有11种,占总种数的57.9%,表明该群落的优势种较为显著。除华山松外,宝天曼保护区陕西紫茎的乔木层树种全部为落叶阔叶林,这与浙江古田山保护区紫茎林乔木层树种以常绿成分的种类占优势是明显不同的[12]。

表4 陕西紫茎林乔木层物种重要值
灌木层盖度约为 40%,高度为 0.3~2.2 m,共出现种子植物(含幼苗)20科28属43种。根据其物种重要值(表5),优势种为伏牛箭竹、长柄绣球、南蛇藤、荚蒾等;重要值小于2的有31种,占总种数的72.09%,如钝叶蔷薇(Rosa sertata)等。说明灌木优势种类也是比较明显的。该林分中,除伏牛箭竹为常绿种类外,其他灌木全部为落叶种类。
层间植物可归于灌木类。样地内有南蛇藤、粉背南蛇藤、清风藤(Sabia japonica)、五味子(Schisandra chinensis)、绣球藤(Clematis montana Buch-Ham.ex DC),层间植物的多样化,使陕西紫茎的群落结构更为复杂。

表5 陕西紫茎林灌木层物种重要值
陕西紫茎林草本层平均盖度约40%,分布均匀,种类丰富。调查显示,群落内草本植物共计15科24属29种,主要由地面芽植物和地下芽种类组成。优势种是糙苏、球果堇菜、犁头叶堇菜、东亚唐松草、斑叶堇菜、鬼灯擎、野菊、京大戟、宽叶苔草(Carex siderosticta)等。这些种类均可在局部形成优势群落(表6)。

表6 陕西紫茎林草本层物种重要值
2.2.5 群落物种多样性 生物群落是一个自然历史演变的过程,是在一定的地理区域内同一环境下不同物种的种群组合。群落物种多样性是群落在组成和结构上表现出的多样性,是认识群落的组织水平和功能状态的基础。经过调查计算陕西紫茎林乔木层、灌木层和草本层物种多样性指数(表7),可以发现,宝天曼自然保护区陕西紫茎群落生物多样性丰富,是我国暖温带地区落叶阔叶林生物多样性分布最高的地区之一。陕西紫茎林不同层次的物种多样性大小依次为灌木层>草本层>乔木层,且各层次物种的均匀度较高。这种生物多样性指数的分布格局与紫茎属植物中心分布区浙江古田山紫茎群落相比有很大不同[12]。总体上宝天曼自然保护区陕西紫茎林生物多样性大于浙江古田山。与宝天曼保护区内其他主要森林群落的物种多样性相比较,陕西紫茎林的Shannon-Wiener多样性指数也高于建群种锐齿栎群落的 1.43~2.18 及栓皮栎群落的 1.64~2.00[14-15]。

表7 陕西紫茎林群落的物种多样性
2.3 园林景观群落设计
园林植物的观赏特性是植物的美化功能和艺术效应的结合。陕西紫茎具有独特的形态、色彩、风韵和芳香等美的特色。根据陕西紫茎的生态习性、生物学特性、伴生树种和观赏特点,从保持园林紫茎群落结构的稳定性和色彩、质地等方面,进行陕西紫茎园林绿化应用的群落设计。
2.3.1 陕西紫茎风景林或行道树栽植 陕西紫茎在野外可以长成15 m高的大树,并且较耐寒冷,在伏牛山区-10℃的天气情况下能在野外安全越冬,陕西紫茎群落生长良好,植株相对较为高大,冠大荫浓,树形清秀。因此,可以在城市绿地、公园、庭院、草坪的角隅等地,营造陕西紫茎风景林(纯林),或对植于堂前和路口处,赏其幽雅的姿态。也可以作为小区道路、人行道绿带等行道树种,能够发挥出春季观花、夏季遮阳、秋冬季观姿、美化街景以及装饰建筑立面等功能。
2.3.2 落叶阔叶风景林 落叶阔叶林是河南省地带性的植被类型。无论是在原产区还是在伏牛山区,陕西紫茎与壳斗科、槭树科、樟科、榆科等暖温带的落叶植物种类具有较强的种间联结性,已与这些植物种类形成了稳定的落叶阔叶林群落。因此,在园林应用中,应根据其生物学、生态学特性和树种间的互利关系,通过乔灌草的合理配置,营造陕西紫茎风景林,在发挥其观赏特性前提下,保持陕西紫茎景观园林的稳定性。
2.3.3 常绿与落叶阔叶混交林风景林 陕西紫茎属中性偏阳且喜湿润肥沃的酸性土壤的树种,宜植于水、肥、热均佳处。一般丛植于林缘,背景衬以樟树、石楠、桂花、松类等常绿针阔叶树种,林下灌木层配置蔷薇类和黄杨、海桐等常绿灌木,更能显其清雅脱俗的清姿丽质。陕西紫茎具有较高的观赏价值,也可以和其他常绿的山茶科植物一起,在我国中部地区营造山茶科植物专类园,创造出春花秋叶冬姿的特色景观。
3 结论与讨论
宝天曼自然保护区陕西紫茎群落物种丰富,植物科属组成较为分散,从属的分布区类型来看,温带分布的属占据优势;群落的叶级谱以中型叶为主,叶型以单叶为主,叶质中草质叶占绝对优势;群落的生活型以高位芽植物为主,具有一定的层间植物;群落乔木层、灌木层和草本层的物种多样性大小依次为灌木层>草本层>乔木层。根据陕西紫茎的观赏特性和生态习性,提出河南省园林造景中陕西紫茎纯林、落叶阔叶林和常绿与落叶阔叶混交林的3种景观配置方式。
陕西紫茎在河南已经达到其最北的分布区边沿,本研究提出了陕西紫茎的群落结构特征和园林景观的营造模式。陕西紫茎在群落内树高处于第2层,与千金榆、青楷槭等树种共优共存,整个群落乔木层生物多样性指数低于灌木层和草本层。宝天曼保护区区域土壤肥沃,林内物种更新良好,幼苗种类较多,这也是陕西紫茎群落内灌木层和草本层物种多样性较高的原因之一。生物多样性能够反映群落的物种构成及功能规律,生物多样性指数的大小与群落的丰富度有关,但也与取样的面积和技术有一定的关联性。为了更全面地研究宝天曼自然保护区陕西紫茎林群落构成和生态功能,需要扩大样地的取样面积,排除调查样地取样的偶然性,为开展陕西紫茎的引种驯化和园林绿化提供更加详实科学的技术资料。
[1] 李捷.山茶科紫茎属和舟柄茶属的系统学研究[J].植物分类学报,1996 34(1):48-67.
[2] 石祥刚.中国山茶科属种地理学研究[D].广州:中山大学,2003.
[3] 史作民,刘世荣,河南宝天曼种子植物区系特征[J].西北植物学报,1996,16(3):329-335.
[4] 卢炯林,王磐基,王正用.河南宝天曼保护区珍稀濒危植物资源的初步研究[J].河南大学学报,1988(3):35-41.
[5] 王亚平,韦博良,殷卫抗,等.宝天曼杂木林样地的物种组成及空间分布格局[J].河南农业科学,2014,3(4):95-100.
[6] 朱学灵,吴明作.旅游活动对宝天曼保护区珍稀植物群落陕西紫茎林的影响[J].西部林业科学,2009,38(1):42-47.
[7] 丁宝章,王遂义.河南植物志[M].郑州:河南科学技术出版社:1981—1998.
[8] 吴征镒.中国植物区系属的分布区类型[J].云南植物研究,1991(13):1-139.
[9] Raunkaier C.The life form of plant and statistical plant geography[M].Oxford:Clarendon Press,1934:623.
[10] 马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究Ⅱ.丰富度、均匀度和物种多样性指数[J].生态学报,1995,15(3):268-277.
[11] 马克平.生物群落多样性的测度方法I.α多样性的测度方法(上) [J].生物多样性,1994,2(3):162-168.
[12] 马克平.生物群落多样性的测度方法I.α多样性的测度方法(下) [J].生物多样性,1994,2(4):231-239.
[13] 胡正华,于明坚,余建平,等.浙江古田山自然保护区紫茎林特征研究[J].中国计量学报,2002,13(3):199-204.
[14] 程瑞梅,刘玉萃,蒋有绪,等.河南宝天曼锐齿栎林群落学特征[J].生态学杂志,1999,18(4):25-30.
[15] 程瑞梅,肖文发.河南宝天曼栓皮栎林群落特征及物种多样性[J].植物资源与环境,1998,7(4):8-13.
猜你喜欢
当代水产(2022年8期)2022-09-20
贵州畜牧兽医(2022年3期)2022-06-28
昆明医科大学学报(2022年2期)2022-03-29
现代园艺(2021年23期)2021-12-01
食品安全导刊(2021年20期)2021-08-30
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
河南科学(2020年3期)2020-06-02
智富时代(2019年8期)2019-09-23
智富时代(2019年8期)2019-09-23
