
运动员专项动作识别的神经效率:来自EEG节律去同步化和相干性证据
2018-03-11 06:54李安民
天津体育学院学报 2018年4期
魏 瑶,李安民
神经效率假说认为,在认知任务加工期间,与低绩效者相比,高绩效被试任务皮层激活降低,表现大脑神经功能的高效性[1-2]。研究者采用正电子断层扫描(PET)、功能性磁共振成像信号(fMRI)和脑电图(EEG)技术,进行智力测验、词语流畅、空间认知、工作记忆等任务试验,结果显示,高绩效者任务皮层的激活水平低于对照组[3-6],与神经效率假说一致。练习对大脑皮层激活的影响研究表明,后天学习诱发大脑皮层功能适应性变化,表现为皮层激活程度的改变[1,5-10]。这些结果表明,个体不同认知能力,可以通过大脑皮层激活差异性呈现,并且,练习可能降低大脑任务皮层激活水平,提高大脑皮层的神经效率。一些视知觉学习、音乐家和运动员数据也支持上述推论[11-14]。但是,另一些工作记忆、逻辑推理、数字转换等任务研究显示,高绩效者的任务皮层激活更强[15-19]。练习对脑区任务皮层激活程度的改变趋势,有皮层下降,也有皮层增强,还有下降和增强的皮层同时呈现[20-21]。因此,大脑认知皮层神经活动机制十分复杂,将神经效率解释为皮层激活降低,可能过于简单化,需要进一步深入探讨[22]。
依据大脑皮层功能的特异性观点,不同认知能力的皮层功能区域存在差异。但是,大脑皮层功能也体现系统性,大脑认知是不同脑区、不同层次的协同过程[23],即使是简单任务,也诱发大脑皮层多层次、多区域激活[21,24]。大脑神经图论研究显示,脑功能网络是全域性小世界拓扑和局域性心理模块化的统一[25-26],是大脑主干网络、局域网络和交互网络的复杂连接[27-29]。因此,探讨个体大脑皮层神经效率,既要分析脑区皮层功能激活,又要评价皮层间功能连接[6,30-32]。目前,大脑认知神经活动的测量方法,主要包括神经电生理和功能影像学技术。其中,神经电生理技术,采用EEG低频节律去同步化(如alpha-ERD)评价激活水平;功能影像学技术,采用fMRI血氧信号推测皮层激活程度。对大脑皮层间功能连接的评价,神经电生理通过计算电极对EEG节律相干性系数;功能影像学计算兴趣区的fMRI信号因果连接系数[33-35]。由于EEG节律震荡活动直接反映神经元活动,具有毫秒级的时间分辨率,在一些大尺度脑功能研究中,研究者常常采用EEG节律震荡信号,揭示大脑认知加工的神经机制[36-38],并将短距离(脑区内)神经元震荡视为皮层激活程度,用长距离(脑区间)神经元震荡评价功能连接程度[39-41]。因此,脑区皮层激活与功能连接,具有相同生理机制。
运动领域常常采用横向对比研究,通过检验运动员和对照组的差异性,验证长期训练对大脑皮层可塑性影响。运动员大脑神经效率研究,可以采用专家-新手范式,揭示长期训练对运动员大脑皮层神经效率的影响。由于运动项目的特异性,不同项目训练的大脑皮层功能变化,可能存在差异性。但相关研究显示,运动认知心理机制是感知-动作循环,主要涉及大脑额、顶、枕、颞区等相关皮层[-46]请核对。一些视觉-动作控制任务EEG研究显示,与对照组相比,运动员任务皮层激活下降。如在简单专项动作控制任务中,射击选手中央区和枕叶皮层alpha-ERD水平更低[47];艺术体操和空手道选手,在专项动作评价任务期间,其枕-颞、枕-顶和镜像系统alpha-ERD更低[48]。一些运动认知fMRI数据也显示,与对照组相比,运动员任务皮层激活更低。如在简单动作反应和视-空任务中,职业赛车手的辅助运动区、顶区和额下区激活低于对照组[49];在简单脚部动作控制任务期间,职业足球精英队员的动作皮层的激活低于对照组[50]。上述结果与神经效率假说一致。但是,另一些fMRI研究则显示,专项动作识别和表象任务期间,与对照组相比,运动员镜像系统(MNS)、动作观察网络(AON)、默认网络(DON)和前运动区(PMC)等皮层诱发激活更高[51-53],结果与神经效率假说不一致。可见,运动领域大脑皮层神经效率,可能存在条件性限制,不仅关注皮层激活变化趋势,还要分析大脑功能网络特征。
前人研究显示,练习诱发大脑皮层的可塑性[54],并且,大脑认知加工熟练化,表现心理模块化自动化趋势。其中,心理模块内的连接增强,模块间的连接削弱[55]。从运动认知研究的心理行为数据来看,练习导致个体认知加工方式,从控制性向自动化转变[10],表现为高级认知控制皮层的能量消耗降低,节约中枢认知资源,提高了练习者行为绩效[56]。神经生理学研究显示,练习对大脑皮层功能可塑性,可能表现为神经元功能的适应性变化,神经元连接突触的数量优化,以及功能有效连接的强化[2,12,20]。运动领域相关研究也显示,长期训练可能诱发脑区功能连接下降。如在专项任务期间,与对照组相比,射击选手的左侧颞-额(alpha节律)相干性系数下降[30,57];高尔夫有效推杆任务中,高尔夫选手的左侧颞-额(alpha节律)相干性更低[58];在专项动作识别任务中,乒乓球选手的左侧颞-额(theta)节律更低相干性[59],动作表象任务诱发乒乓球运动员左侧顶-颞(alpha)节律更低相干性[60],这些结果表明功能连接程度下降了。然而,另一些研究结果显示,长期训练诱发脑区间功能连接增强。如在乒乓球接发球动作表象任务中,运动员右侧颞-额(theta)节律相干性更高[53,61];在一般视觉-空间任务中,乒乓球运动员的顶-枕、顶-额(alpha)节律相干性更高[62];大脑fMRI数据显示,在简单视-动任务期间,职业赛车手左侧顶-颞、颞中回-前运动区连接系数更高[49]。大脑功能连接变化趋势的不一致,可能与任务属性、加工方式、皮层环路等因素相关[6,21,32]。心理模块化是认知熟练化自动化的重要特征,运动训练对大脑功能连接的影响,可能表现为脑功能网络的优化,即强化有效功能连接,削弱低效或无效连接[63-65]。可能受调节变量的影响,脑功能连接变化趋势并不一致。因此,从脑区功能连接视角,分析大脑皮层的神经效率,需要明确所分析的功能连接通路,是否属于熟练化运动认知加工的有效连接。
专项动作的识别,是运动认知加工的重要环节,特别是开式技能运动项目,运动情境复杂多变,对运动员的动作识别能力提出更高要求。视知觉学习研究显示,练习诱发皮层功能可塑性,不仅涉及视觉皮层,还诱发额、颞等高级中枢功能性改变[66-67],并且,在不同学习阶段和任务复杂程度条件下,表现皮层激活变化与激活皮层分布区域的差异性[68-70]。动作识别的神经机制主要涉及顶-枕、颞-枕视觉通路,以及额-顶镜像神经元[71-72],其中,顶内沟/下顶叶(IPS/IPL)-额下弓形沟(IAS)-颞上沟(STS)之间交互作用,是动作识别加工的主要神经环路[73]。根据大脑皮层的可塑性,长期专项运动训练,很可能诱发动作识别皮层功能可塑性。不仅提高动作识别的行为绩效,还可能诱发任务皮层的可塑性,优化动作认知神经网络,降低皮层激活程度,提高运动员大脑神经效率。但是,目前专项动作识别的研究显示,运动员的任务皮层激活和功能连接的变化趋势并不一致[53,60-61,74]。由于运动情境快速、复杂、多变,与一般物体和空间识别相比,专项动作识别加工,特别是快速对抗性项目动作识别任务,可能诱发更广泛的皮层功能激活范围,对神经网络连接的可塑性变化更复杂。大脑功能偏侧化,也可能进一步影响皮层激活和功能连接的变化趋势[75-76]。此外,在专项动作识别的神经效率研究中,特别是采用专家新手范式,诱发任务的选择尤其重要,既要考虑生态学效度,又要保证专家新手加工方式和诱发皮层环路一致性。而受被试的运动经验的影响,诱发任务容易导致专家和新手的加工方式和皮层环路不同,那么这种条件下皮层激活变化,无法验证神经效率假说,激活组间差异可能是经验策略,而不是神经元功能的差异性[22]。但是,目前运动领域专项任务加工神经机制研究,很少考虑运动经验的调节效应,这在神经效率研究中应特别注意。
为了从大脑皮层的激活和功能连接视角,分析运动员的专项动作识别的神经活动特征,揭示长期训练对运动员皮层功能的可塑性,验证运动员大脑皮层的神经效率。根据目前相关研究结果,本研究应用脑电技术,选择典型快速对抗性乒乓球项目,采用专家新手范式,通过发球动作识别任务试验,比较专家和新手的行为和脑电数据组间差异性,分析长期训练对专项动作识别皮层功能的影响。依据大脑可塑性理论和大脑神经效率假说,提出如下假设:(1)乒乓球运动员专项动作识别行为绩效优于对照组;(2)乒乓球运动员动作识别相关皮层诱发的alpha节律ERD水平低于对照组;(3)乒乓球运动员的右侧枕-颞、枕-额、颞-额等脑区电极对theta、alpha等节律相干性系数更高,左侧枕-颞、颞-额、顶-颞等电极对theta、alpha节律相干性系数更低。为验证上述假设,随机抽取乒乓球二级运动员为专家组,乒乓球普修班大学生为新手组,完成乒乓球发旋转球动作识别的试验任务,同步记录行为数据和脑电信号,通过比较和分析被试的反应时、正确率、ERD/S和相干性系数组间差异,验证运动员专项动作识别的神经效率。
1 研究对象与方法
1.1被试
随机抽取乒乓球国家二级大学生运动员为专家组(n=19,Age=20.2±1.2),训练年限都是10年以上,并随机抽取乒乓球普修水平的大学生为新手组(n=20,Age=20.8±0.9),研究任务为发不同旋转球的动作识别。被试智力正常,身心健康,无精神病史,都是全日制在校大学生,且均为右利手,平均年龄组间差异不显著(t=0.34,P>0.05)。试验前阅读任务保证书,均是自愿参与试验,通过主试的讲解,较快熟悉试验流程与要求。试验结束后支付一定报酬。
1.2 试验任务
试验前,被试填写基本信息,并佩戴好电极帽,调整座椅位置等预备工作,让被试舒适地坐在计算机屏幕前,双眼平视屏幕中心位置,相距90 cm。试验开始显示指导语,告知试验任务与要求,试验采用E-Prime2.0程序编写任务呈现序列,每个刺激任务呈现之前,屏幕显示“+”提示,然后呈现刺激任务,时间设置3 s,按键反应后任务消失,缓冲2 s,然后呈现下一个trail(见图1)。全部任务是乒乓球不同旋转发球的动作视频。由于乒乓球发球动作复杂多变,识别难度大,新手因难度大可能采取随机猜测方式,导致2组被试的加工方式和神经通路都不同,影响行为和脑电数据的有效性。根据一般动作程序理论,动作速度属于动作模型的易变部分,调整速度而并不改变动作的基本属性[77]。因此,本研究将刺激呈现速度定格为16帧/s,既不改变动作属性,保证生态学效度,又降低动作识别难度,确保2组被试采用的加工方式和诱发皮层通路相同。本研究将能够发侧下旋球的动作作为靶刺激,其余为非靶刺激,两者比例为1:1,全部刺激任务共140个,试验过程随机呈现刺激,要求被试辨认是否属于发侧下旋球的动作,是按“1”键,不是不按键。试验过程设休息键,便于被试消除中途疲劳,按任意键继续,全部试验时间大约30 min。

图1 试验刺激的呈现程序Figure1 Rendering Procedure of Experimental Stimuli
1.3 仪器与数据采集
使用2台DELL计算机记录试验数据,其中,一台采用EPrime 2.0程序,编制试验程序和播放刺激任务,并收集被试在线行为数据,用于计算正确率和反应时指标值。另一台同步连接德国Brain Products公司64导脑电记录电极帽和放大器,通过BP Recorder2.0软件,同步采集任务诱发被试脑电数据,便于后期离线处理。电极帽为标准10-20系统,AFz为参考电极点,FCz为接地电极点(Ground)。全部电极电阻小于5 KΩ,信号带宽0.1~100 Hz,采样率为256 Hz。
1.4 脑电数据处理
(1)首先,对全部被试的离线脑电数据进行预处理。主要包括以双耳乳突的平均电位为参考值,更改原参考电极点;采用独立成分法(ICA)去除眨眼伪迹,并排除噪音干扰和其他伪迹;采用40 Hz低通滤波,去除50 Hz市电干扰,并将波幅大于±100 μV伪迹自动剔除。其次,对预处理后的脑电数据,按照刺激锁时方法处理。由于绝大部分被试反应时均小于2 s,本试验EEG分段采用刺激锁时方式,依据同步记录的mark点,将被试回答正确的脑电数据分段,包括基线期(-2 000 ms,0)和事件期(0,2 000 ms)2段。预处理数据用于alpha-ERD/S和相干性系数的计算。
(2)根据前人有关视觉动作识别加工诱发脑区特征[30,50],大脑功能的偏侧化特征[53,59,74,76],以及有关 alpha节律去同步化/同步化反映皮层激活的生理意义[78]。本研究通过计算皮层诱发EEG的低、高频alpha-ERD/S水平,评价视觉动作识别相关兴趣区皮层激活变化。计算公式为:ERD/S=(E-P)/P×100%,E表示事件期节律功率值,P表示基线期节律功率值。首先,将预处理脑电数据,通过BP分析软件包,进行傅里叶变换,并提取事件期和基线期的不同节律(如alpha)功率值;其次,采用SPSS编程计算全部被试的alpha-ERD/S。负值表示节律波去同步化(ERD),正值表示节律同步化(ERS),其中,alpha-ERD和ERS分别反映皮层的激活或抑制水平。本研究目的是探讨任务正相关皮层网络(TPN),主要包括视觉背侧通路(BAs7/5/6d)、腹侧通路(BAs19/37/20)和镜像系统(BAs40/6v/44/45)等兴趣区(ROI),主要计算O1、O2、P3、P4、P5、P6、F5、F6、T7和T8等电极点的alpha-ERD/S值。
(3)根据大脑网络图论研究,大脑认知网络呈现全域性小世界拓扑和局域性模块化特征,并且,熟练化加工促进大脑网络功能模块化,熟练化有效功能连接加强,与熟练化无关的连接减弱[55],能够最大限度提高大脑皮层认知加工效率[79],进而降低局部皮层激活水平。神经科学研究显示,练习诱发神经皮层功能重组,神经元现有突触连接强化或新突触的形成,因此,神经效率可能与大脑功能连接的改变相关[2,21]。因此,本研究通过计算诱发脑区EEG节律的相干性系数,反映皮层间的神经功能耦合程度。相干性系数计算公式为:Coh(x,y)(f)=Sxy(f)/[Sxx(f)×Syy(f)]1/2。其中,Coh(x,y)(f)表示x和y2个信号f节律的相干系数,反映不同脑区皮层间功能耦合程度,取值范围[0,1],“1”说明信号y是x的在某一频频段上完全相关,“0”表明信号y是x的在某一频段完全独立;Sxy表示x(t)和y(t)信号的交叉能量谱(Cross-power spectrum),Sxx和Syy分别是x(t)和y(t)信号的自频谱(Auto-spectra)。运行BP分析软件包的相干性计算工具,计算任务期诱发电极对的 theta(4~7 Hz),low alpha(8~10 Hz),high alpha(11~13 Hz)节律波相干性系数。并且,为了避免其他潜在环路(如抑制加工环路)对任务皮层连接分析的干扰,本研究只探讨皮层激活增强脑区的相干系数,主要计算左半球内(O1P3,O1T7,O1F3,T7F3,P3F3,T7P3)、右半球内(O2P4,O2T8,O2F4,T8F4,P4F4,T8P4)和半球间(O1O2,P3P4,T7T8,F3F4)脑区电极对相干性系数(见图2)。
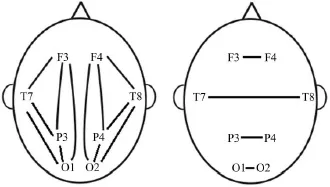
图2 半球内或半球间电极对的相干性Figure2 Coherence of Inter/Intra-hemispheric Electrode Pairings
1.5 统计分析
(1)本研究运用KS方法,对行为和脑电指标数据进行正态性检验。为了验证假设一,研究采用独立样本T检验法,检验反应时的组间差异显著性(α=0.05,下同);运用Z检验法,检验被试正确率的组间差异显著性。(2)为了分析专项动作识别诱发皮层脑电特征,本研究依据脑区电极点分布与前期研究结果[80-81],计算被试O1、O2、T7、T8 、P3、P4、P5、P6、F5和F6等电极点EEG信号指标。采用Mauchly球型度,检验不同被试组方差齐性,并对方差不齐性的数据,采用Greenhouse-Geisser法进行自由度矫正。(3)为了验证假设二,采用三因素(2组别×2半球×2脑区电极点)重复测量方差分析法,分别对背、腹侧视觉通路,以及镜像系统的主要电极点,进行alpha-ERD/S水平的组间差异性检验。(4)为了验证假设三,采用三因素(2组别×2半球×6电极对)重复测量方差分析方法,分别对O1P3、O1T7、O1F3、F3T7、F3P3、T7P3电极对,以及O2P4、O2T8、O2F4、F4T8、F4P4、T8P4电极对,进行theta、低频和高频alpha节律相干性系数的组间差异性检验;采用两因素(2组别×4脑区)重复测量方差分析方法,对O1O2、T7T8、P3P4、F3F4电极对进行theta、低频和高频alpha节律相干性系数的组间差异性检验。(5)对方差分析交互效应显著的结果,进行简单效应分析与事后比较,进一步明确脑电指标的组间差异显著性。
2结果
2.1 行为数据
本研究分别对被试反应时和正确率进行组间差异显著性检验,反应时T检验结果显示,专家组[n=19,(1 520±173)ms]与新手组[n=18,(1 691±167)ms]差异不显著(t=1.34,P>0.05);正确率Z检验结果显示,专家组(n=19,84.85%±4.22%)和新手组(n=18,74.01%±8.59%)差异显著(Z=2.71,P<0.05),专家组的动作识别正确率高于新手组。
2.2 Alpha-ERD/S水平的比较
低频alpha-ERD/S的组间差异性检验结果显示:背侧通路(O1、O2、P3、P4)的组间主效应显著[F(1,35)=8.398,P<0.01,ŋ2=0.194],半球主效应显著[F(1,35)=13.218,P<0.01,ŋ2=0.274],组别、半球和脑区电极点交互效应不显著[F(1,35)=1.141,P>0.05,ŋ2=0.032];腹侧通路(O1、O2、T7、T8)的组别主效应显著[F(1,35)=5.336,P<0.05,ŋ2=0.132],半球主效应显著[F(1,35)=45.266,P<0.01,ŋ2=0.564];镜像系统(F5、F6 、P5、P6)的组别主效应不显著[F(1,35)=2.591,P>0.05,ŋ2=0.132],组别、半球、脑区电极点交互效应也不显著[F(1,35)=2.995,P>0.05,ŋ2=0.079]。事后比较显示,专家组枕-顶视觉通路O1、O2、P3、P4电极点的低频alpha-ERD低于新手组(见图3)。
高频alpha-ERD/S的组间差异性检验结果显示:背侧通路(O1、O2、P3、P4)的半球主效应显著[F(1,35)=6.520,P<0.01,ŋ2=0.220],组别主效应显著[F(1,35)=17.767,P<0.01,ŋ2=0.337],半球、脑区电极点和组间交互效应显著[F(1,35)=6.392,P<0.05,ŋ2=0.154];腹侧通路(O1、O2、T7、T8)的组别主效应显著[F(1,35)=17.120,P<0.01,ŋ2=0.328],半球组别交互效应显著[F(1,35)=4.867,P<0.05,ŋ2=0.122],组别和脑区交互效应显著[F(1,35)=13.059,P<0.01,ŋ2=0.272],半球、电极点和组间交互效应显著[F(1,35)=15.504,P<0.05,ŋ2=0.307];镜像系统组别主效应显著[F(1,35)=30.025,P<0.01,ŋ2=0.462],脑区电极点主效应显著[F(1,35)=9.876,P<0.01,ŋ2=0.220],半 球 主 效应 显 著 [F(1,35)=11.924,P<0.01,ŋ2=0.254],脑区与组别交互效应显著[F(1,35)=19.027,P<0.01,ŋ2=0.352]。简单效应分析显示:左侧顶区[F(1,35)=13.574,P<0.01,ŋ2=0.279]组间差异显著,左侧枕区[F(1,35)=12.351,P<0.01,ŋ2=0.262]和右侧枕区[F(1,35)=13.959,P<0.01,ŋ2=0.285]组间差异显著,左侧颞区(T7)组间差异显著[F(1,35)=13.744,P<0.01,ŋ2=0.282],镜像系统双侧(F5、F6)额中回[F(1,35)=41.639,P<0.01,ŋ2=0.543]和双侧(P5、P6)下顶叶[F(1,35)=17.883,P<0.01,ŋ2=0.338]组间差异显著。事后比较显示:专家组左侧上顶叶(P3)、左侧颞区(T7)、双侧下顶叶(P5、P6),双侧额中叶(F5、F6)和双侧枕叶(O1、O2)高频alpha-ERD低于新手组(P<0.01)(见图3)。
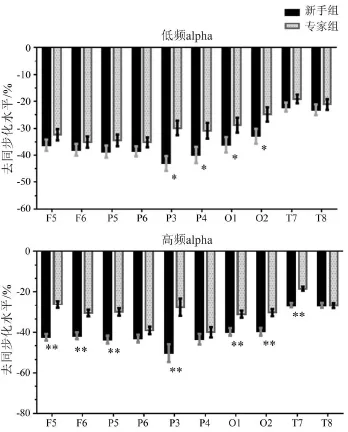
图3 兴趣区电极点低频和高频Alpha节律ERD/S均值Figure3 Low/High-alpha Average ERD/S of the ROIS Electrode
2.3 EEG节律相干性的比较
(1)进行theta节律相干性系数的差异性检验。半球内重复测量(2组别×2半球×6电极对)方差分析显示:组别主效应不显著显著[F(1,35)=1.417,P>0.05,ŋ2=0.039],半球主效应显著[F(1,35)=6.292,P<0.05,ŋ2=0.152];脑区电极对主效应显著[F(5,175)=585.491,P<0.01,ŋ2=0.944],半球和组别交互效应显著[F(1,35)=39.257,P<0.01,ŋ2=0.529],脑区电极对与组别交互效应不显著[F(5,175)=1.133,P>0.05,ŋ2=0.031],半球、脑区电极对和组别交互效应显著[F(5,175)=3.888,P<0.01,ŋ2=0.101]。事后比较显示:专家组左侧额-颞(F3T7)相干性系数均值低于新手 组 [F(1,35)=4.023,P<0.05,ŋ2=0.121];专 家 组 右 侧 枕 -颞(O2T8)相干性高于新手组[F(1,35)=4.884,P<0.05,ŋ2=0.122],专家组的右侧枕-颞(O2F4)相干性高于新手组[F(1,35)=5.241,P<0.05,ŋ2=0.151],右侧额-颞(F4T8)相干性高于业余组[F(1,35)=19.721,P<0.01,ŋ2=0.361]。半球间重复测量(2组别×4 脑区电极对)方差分析显示:脑区主效应显著[F(3,105)=216.345,P<0.01,ŋ2=0.704],组别主效应显著[F(1,35)=25.167,P<0.01,ŋ2=0.418],脑区和组别交互效应显著[F(3,105)=4.082,P<0.05,ŋ2=0.104]。事后比较显示:专家组额区[F(1,35)=8.916,P<0.01,ŋ2=0.203]、顶区[F(1,35)=6.762,P<0.05,ŋ2=0.133]和颞叶[F(1,35)=5.447,P<0.05,ŋ2=0.121]半球间相干性系数低于新手组(见图4)。
(2)进行低频alpha相干性系数的差异性检验。半球内重复测量(2组别×2半球×6电极对)方差分析显示:组别主效应不显著[F(1,35)=0.852,P>0.05,ŋ2=0.024],半球主效应不显著[F(1,35)=0.625,P<0.05,ŋ2=0.018];脑区电极对主效应显著[F(5,175)=248.491,P<0.01,ŋ2=0.876],半球和组别交互效应显著[F(1,35)=17.541,P<0.01,ŋ2=0.334],电极对与组别交互效应不显著[F(1,35)=1.651,P<0.05,ŋ2=0.045],半球、电极对和组别的交互效应显著[F(5,175)=6.235,P<0.01,ŋ2=0.151]。事后比较显示:专家组左半球颞-额(T7F3)相干性低于新手组[F(1,35)=12.615,P<0.01,ŋ2=0.265],而右半球颞-额(T8-F4)相干性高于新手组[F(1,35)=2.105,P<0.05,ŋ2=0.057]。半球间重复测量(2组别×4脑区电极对)方差分析显示:组别主效应显著[F(1,3)=5.520,P<0.05,ŋ2=0.136],脑区主效应显著[F(3,105)=383.781,P<0.01,ŋ2=0.916],组别与脑区交互效应显著[F(3,105)=4.646,P<0.05,ŋ2=0.117]。事后比较显示:专家组额区半球间[F(1,35)=6.243,P<0.05,ŋ2=0.151]、顶区半球间[F(1,35)=6.958,P<0.05,ŋ2=0.165]和颞区半球间[F(1,35)=4.174,P<0.05,ŋ2=0.093]电极对相干性系数低于新手组(见图4)。
(3)进行高频alpha节律相干性的差异性检验。半球内(2组别×2半球×6电极对)重复测量方差分析显示:组别主效应不显著 [F(1,35)=0.138,P>0.05,ŋ2=0.004],半球 主效应 不显 著[F(1,35)=0.483,P>0.05,ŋ2=0.014];电极对主效应显著[F(5,175)=929.502,P<0.01,ŋ2=0.964],半球和组别交互效应显著[F(1,35)=4.991,P<0.05,ŋ2=0.125],电极对与组别交互效应显著[F(5,175)=7.511,P<0.01,ŋ2=0.177],半球、电极对和组别的交互效应显著[F(5,175)=5.741,P<0.05,ŋ2=0.145]。事后比较显示:左半球内的电极对中,专家组的左侧枕-顶(O1P3)电极对[F(1,35)=4.768,P<0.05,ŋ2=0.112]、枕-额(O1F3)电极对[F(1,35)=5.541,P<0.05,ŋ2=0.137]、颞-额(T7F3)电极对[F(1,35)=4.987,P<0.05,ŋ2=0.124]和额-顶(F3P3)电极对[F(1,35)=5.619,P<0.05,ŋ2=0.138]相干性系数低于新手组;右半球内的电极对中,专家组的右侧枕-颞(O2T8)电极对[F(1,35)=4.775,P<0.05,ŋ2=0.182]、枕-额(O2F4)电极对[F(1,35)=6.763,P<0.05,ŋ2=0.174]、额-颞(F4T8)电极对[F(1,35)=7.945,P<0.01,ŋ2=0.219]相干性高于新手组。半球间电极对(2组别×4脑区电极对)重复测量方差分析显示:组别主效应显 著[F(1,35)=5.551,P<0.05,ŋ2=0.137],脑区 主效 应显 著[F(1,35)=671.981,P<0.01,ŋ2=0.950],脑区与组别交互效应显著[F(3,105)=7.659,P<0.01,ŋ2=0.180]。事后比较显示:专家组的左右额区[F(1,35)=8.635,P<0.01,ŋ2=0.198]、左右顶区[F(1,35)=7.961,P<0.01,ŋ2=0.185]电极对的相干性低于新手组(见图4)。
3讨论
为了通过乒乓球专项任务试验的行为绩效和脑电数据分析,揭示专项运动训练对动作识别皮层神经效率影响,本研究通过比较和分析专家和新手2组被试的行为和脑电数据差异性,验证研究假设。

图4 兴趣区电极对theta、低频alpha和高频alpha节律相干性系数图片重新修订请核对Figure4 Coherence of the ROIS Electrode Pairings for Theta,Lowalpha,High-alpha
3.1 行为绩效分析
为了验证假设一,本研究分析2组被试的反应时、准确率等行为数据。总体来看,2组被试都能正常识别发球动作任务,正确率都较高,这说明新手并不是采取随机猜测方式,而是努力而认真地完成试验任务。本研究采用较慢呈现速度,2组被试(特别是新手)都能感知提取识别动作所需要的信息,完成发球动作识别加工。同时,全部被试动作识别正确率较高,表明刺激任务呈现速度较慢,并不改变发球动作属性[77],保证了研究生态效度,也尽量避免2组被试加工方式与皮层环路的差异性。行为数据的组间差异性检验显示,专家组动作识别正确率高于对照组,反应时组间差异不显著,这表明在动作识别任务过程,专家识别加工的高绩效加工,可能与识别过程采用准确性为主的策略,而不是追求识别的快速度,结果与前期动作识别相关研究一致[60,75]。专家组呈现的动作识别高正确率,验证了假设一。
专家表现更高动作识别准确性,可能与运动员丰富专项动作经验有关[80]。运动认知是多成分系统、多加工方式协同过程,感知-动作循环是动作认知是心理机制,观察他人和执行相同动作共享编码,具有相同心理表征[42]。专家动作自动化加工模式,不仅节约动作执行加工资源,也能通过感知-动作循环,降低动作识别和理解的资源消耗,提高专家动作觉察、分辨和确认的有效性[81]。物体识别的特征匹配模型认为,知觉是外部信息与记忆特征模型匹配(自下而上),以及记忆特征模型和外部信息反馈(自上而下)的双向信息流交互作用过程[82]。乒乓球发球动作识别加工,也是自上而下与自下而上的交互过程。个体记忆网络的动作经验,影响识别加工过程。其中,专家发球动作特征记忆模型,具有高度概念化、自动化特征,可能通过自上而下反馈信息流,影响动作特征的信息提取与匹配。但是,新手记忆的动作特征模型,概括化水平低,以控制性加工为主,这需要提取更多动作细节信息,消耗更多控制性认知资源,才能完成发球动作特征识别。这种高认知负荷、低效率资源消耗的特征,不利于中枢资源合理分配,降低动作识别的行为绩效。一些篮球[83]、乒乓球[84]相关的眼动研究也证实,专家眼动的眼跳距离短、注视点更集中。专家这种视觉搜索策略,既经济又高效,能降低视觉注意负荷,提高动作识别准确性。总之,专家的长期训练经验,通过感知-动作循环[42],提高动作特征提取与匹配速度,节约中枢神经资源,专家获得更高行为绩效。
3.2 脑区的alpha-ERD/S水平分析
为了验证假设二,本研究计算和比较大脑皮层alpha-ERD,评价相关皮层激活的组间差异性。根据前期视觉信息加工[66-69,85]和动作识别神经机制研究[86],本研究重点考察视觉通路和镜像系统,并采用alpha节律ERD/ERS作为皮层激活评价指标。由于低频alpha-ERD/S与非特异性唤醒有关,高频alpha-ERD/S反映特定任务加工,本研究计算O1、P3、O2、P4、T7、T8、F5、F6、P5、P6等电极点的低频和高频alpha-ERD/S,主要分布视觉通路和镜像系统等皮层区域。其中,视觉腹侧和背侧通路,涉及物体识别和视觉空间加工[87],镜像系统激活与动作理解和计划执行相关[88],顶内沟/下顶叶(IPS/IPL),额下弓形沟(IAS)颞上沟(STS)3个脑区通路的循环信息流,是动作识别理解神经机制[87]。依据动作理解层次模型,动作理解包括视觉运动、动作方式、动作目标和动作意图等不同认知层次,腹侧通路、镜像系统和动作观察网络交互影响,反映动作认知的神经机制[89]。本研究任务诱发2组被试视觉通路和镜像系统激活,进一步验证上述研究结论,同时,被试激活皮层神经环路相同,避免了任务难度或认知策略等因素,影响皮层激活趋势的反转[6,90]。
低频alpha-ERD的组间比较显示,专家背侧腹侧视觉通路O1、O2、P3和P4电极点均值低于新手组,而镜像系统电极点组间差异不显著。低频alpha-ERD被视为大脑非特异性唤醒程度,专家枕叶低频alpha-ERD更低,而镜像系统组间无差异,表明专家视觉皮层的唤醒水平低于对照组,这可能是长期训练诱发视觉神经元兴奋阈限增加了,镜像系统组间无差异,可能性是动作理解的一般唤醒水平与动作类型无关。高频alpha-ERD组间比较显示,专家枕叶(O1、O2)、额下回(F5、F6)、左侧顶叶(P3、P5)和颞叶(T7)电极点低于对照组。高频alpha-ERD被视为大脑皮层特异性认知相关,专家镜像系统和视觉通路(顶、颞)高频alpha-ERD的更低,表示专家大脑在特定动作任务认知过程,其视觉通路和镜像系统都表现更低激活。
其中,运动员枕叶更低激活,可能与运动员记忆系统的运动经验有关。运动员经过长期训练,其长时记忆系统存储有关发球动作特征模型,这些概括化的动作模型,只需少量的动作特征信息,就可完成识别任务。长时记忆存储的动作模式,通过自上而下和自下而上循环信息流,影响视觉皮层对动作特征信息提取效率,随着专家视觉加工注意负荷的降低,提高对动作特征信息检索、编码和匹配有效性,其视觉皮层激活也降低了。有关研究显示,运动员视觉皮层不仅激活下降,其MT+皮层的灰质也增厚了[91]。本研究显示专家视觉皮层激活降低,与前期复杂任务知觉学习显示的,视觉皮层激活下降的结果一致[70],在击剑、体操、空手道、排球、乒乓球等类似研究,也呈现相同的结果[50,89,91-92]。
除了枕叶皮层,专家镜像系统(左侧)和顶叶皮层的激活也低于新手,这表明长期训练,还可能促进高级认知皮层的神经效率。镜像系统是个体动作意图、动作目标和动作识别的重要神经元集群,主要分布在额下回、顶内沟和颞上回等皮层区域。运动员镜像系统激活低于新手的原因,可能是长期视-动信息系统联结学习,提高运动员接发球动作特征识别与理解能力。相关研究显示,运动认知涉及视觉和镜像系统,而且,动作特征信息感知与动作计划执行密切关联[42]。因此,长期训练对动作认知神经环路的影响,可能仅仅包括动作计划执行系统,还可能涉及动作感知系统(包括视觉感知)。专家动作认知自动化,不仅节约大脑控制性注意资源,提高动作认知能力,也可能诱发动作理解有关的镜像系统可塑性[88]。相关研究也显示,随着动作技能熟练化,其皮层激活从前向后、从皮层向皮下转移,额-控制皮层激活降低[93],与本研究结果一致。此外,跆拳道、艺术体操、吉他等动作评价研究也显示,运动员的镜像系统激活下降[13,50-51]。
专家的顶叶激活程度更低,可能是长期训练,提高专家动作识别与理解能力。由于后顶叶皮层与视觉-空间注意相关,专家动作识别加工,可能从初期视觉背侧通路为主,转变为腹侧皮层加工为主,降低专家的视觉空间认知负荷,其顶叶皮层激活下降了。前期镜像书写阅读学习的研究显示,随着练习熟练化,练习者背侧视觉通路的激活也下降了[94]。根据神经生理学研究,大脑皮层激活降低,可能是练习诱发神经元钝化现象[48]。一些大脑可塑性研究也显示,练习诱发大脑皮层激活变化,其原因可能是神经元激活的阈限提高,或者神经突触功能性重组[2,21]。
本研究还显示,专家皮层激活呈现偏侧化趋势,表现在左侧相关皮层的激活下降,专家左侧顶叶和颞叶激活较低,其原因可能与动作识别自动化加工相关。大脑功能偏侧化理论认为,左半球侧重言语逻辑分析,右半球优势是动作-空间加工。长期的专项运动训练,提高专家对发球动作的识别能力,减少左侧言语逻辑分析加工,而新手尽管能够识别发球动作,但由于动作记忆的概括化水平低,需要发球动作特征与理解的言语逻辑分析,左、右半球皮层功能更强整合,才能完成动作特征的识别理解。这种皮层激活偏侧化趋势,节约左侧大脑认知资源,也是运动员大脑皮层神经效率的体现。一些专项和一般认知任务研究中,也呈现这种偏侧化现象[59,75,76]。因此,专家左侧顶-枕皮层激活下降,进一步验证了神经效率假说。但是,舞蹈、篮球等项目的动作观察、表象任务研究显示,与新手相比,专家动作理解的镜像系统激活更强[51-53],这可能与任务难度[6]、默认网络[12]和自信心[49]等因素的调节效应有关。
3.3 脑区间节律相干性分析
为了验证假设三,本研究计算和分析枕-顶、枕-额、顶-额、颞-额电极对事件期theta、alpha节律的相干性系数(见图4)。前期研究认为,简单地将大脑皮层的神经效率,解释为皮层激活降低,并没有真正揭示其内在机制[22]。因此,在分析任务皮层激活差异性基础上,进一步考察诱发枕、顶、颞、额等激活脑区间theta(与一般注意和记忆加工相关)、低频alpha(与一般注意唤醒相关)、高频alpha(与特定任务注意加工相关)相干性是十分必要的。本研究结果显示,专家的右侧枕-额(theta、alpha)、颞-额(低频alpha)、枕-颞(theta、高频alpha)相干性系数高于新手组。但是,左侧颞-额(theta)、额-顶(高频alpha)连接系数低于新手,左右额、顶、颞区内(theta、alpha)相干性也低于对照组。
专家左侧颞-额、额-顶电极对,以及左右额、顶、颞区电极对的相干性系数下降,表明在动作识别过程中,专家这些脑区间功能连接降低。对专家而言,随着动作认知加工逐步自动化,对高级执行控制皮层的依赖性下降,这些相干性系数下降的神经连接,似乎与动作的言语逻辑描述或计划控制相关,并不是动作自动化心理模块连接。这些无效连接增强,反而破坏专家动作认知自动化。对新手而言,由于动作识别的概括化水平低,动作的意识控制要求多,需要高级控制性加工皮层参与。新手动作心理表征的低概括性和高控制性,导致在发球动作识别中,新手需要左侧动作言语描述性信息加工,以及动作空间(右侧)和言语描述(左侧)信息的更强协同,才能顺利完成动作识别加工任务,导致新手左侧半球内和半球间功能连接增强。
专家右侧枕-额、颞-额电极对的相干性系数增加,表明专家的这些脑区功能连接增强。其原因可能是,长期专项训练,优化专家接发球动作的感知-动作循环,提高动作识别与计划控制加工效率。与左侧和半球间脑区的功能连接不同,右侧枕-颞、颞-额脑区功能连接,可能专家动作认知心理模块的有效连接。右侧皮层功能连接增强,可能是专家动作识别加工高效性体现。并且,这种认知加工的高效性,主要依赖右侧皮层功能。动作认知心理模块有效连接的强化,有利于右侧脑区动作感知-行动循环,进而促进专家动作识别与计划执行。一些视知觉神经机制研究显示,大脑的枕、颞、额皮层与基底神经节,构成皮层-基底神经节-皮层结构环路[22,92],这些环路与视觉信息加工密切相关。长期乒乓球训练,对感知-动作循环的影响,可能更多是右侧枕区-基底神经节-额区环路。专家和新手动作认知,都涉及运动的位置知觉和特征识别,只是新手的2种信息加工都重要,其左、右脑区环路广泛激活,而专家主要依赖于右侧皮层环路。因此,专家动作心理模块有效连接增强和无效连接减弱,体现专家大脑网络功能的高效性[55],也验证练习的皮层可塑性[2,21]。专家脑区功能连接的选择性变化,体现了运动员大脑皮层神经效率。
此外,专家左侧脑区间功能连接降低,以及右侧脑区间功能连接的增强,体现大脑皮层功能偏侧化,并且,乒乓球单侧手的操作特征,进一步增强皮层功能偏侧化趋势[53,59,74-76]。新手组被试对发球动作识别能力较低,需要较多的左侧言语描述加工,以及右侧动作-空间认知加工,表现为左侧颞-顶-额网络的功能连接程度强。专家发球动作识别过程,主要依赖右侧感知-动作循环,这些功能连接的增强,提高专家动作识别的皮层神经效率。前期研究显示,职业赛车手右侧视觉运动区-前运动区、乒乓球运动员右侧颞-额区的功能连接更强,射击、高尔夫、乒乓球选手左侧前运动区-颞-枕通路的功能连接降低[10,63-65]。纵向学习结果也显示,练习诱发被试左侧额下回-扣带回-基底神经(主要是壳核)的功能连接降低[65],这与本研究结果一致。因此,对专家动作识别过程,这种左侧言语逻辑皮层连接削弱,也体现专家大脑皮层的神经效率。
3.4 研究的局限性
(1)受制于EEG技术空间局限性、任务属性等因素影响,本研运动员的脑区激活和功能连接分析,空间分辨率较低,属于大尺度皮层功能活动特征,今后可以通过EEG和fMRI技术的结合,解决上述问题。(2)本研究采用专家新手范式的横向比较分析,尽管注意被试年龄、学历、智力等变量的控制,但是,神经皮层活动的差异性,无法确认是先天还是后天,今后研究可结合纵向训练研究,解决上述问题。(3)尽管本研究采用被试都熟悉的简单发球动作识别任务,既保证研究生态效度,又诱发被试相同皮层通路,提高被试皮层神经效率的可比性。本研究选择的专项动作任务,无法避免动作识别策略等知识因素,对个体激活与功能连接的影响[22]。而采用简单的一般性任务,又无法保证专项任务加工的特异性。今后的神经效率研究,还需要优化试验任务设计,既保证诱发被试相同的皮层环路,又避免经验知识对神经皮层功能差异化的干扰效应。(4)运动员大脑皮层的神经效率,是大脑神经元突触和分子等微观功能机制的反映,这些是无创伤脑功能研究手段无法解决,今后考虑通过相关动物模型,从微观层次探讨运动员大脑神经效率。
4结 论
运动员在专项发球动作识别任务中,表现更优行为绩效,以及枕-顶视觉通路和镜像系统皮层更低激活,验证了神经效率假说。并且,运动员的右半球脑区功能相干性系数提高,以及左半球脑区和半球间脑区功能相干性系数降低,表明运动员大脑皮层的神经效率,不仅表现为任务脑区激活下降,还表现为脑功能认知网络的有效连接增强和无效连接减弱。
猜你喜欢
中华实验眼科杂志(2022年6期)2022-11-15
北京航空航天大学学报(2022年6期)2022-07-02
中国典型病例大全(2022年12期)2022-05-13
心理学报(2021年8期)2021-08-11
首都体育学院学报(2019年5期)2019-10-18
中学科技(2018年9期)2018-12-19
中国当代医药(2017年17期)2017-07-25
健康管理(2017年3期)2017-04-20
红领巾·成长(2009年10期)2009-12-02
中学生英语·中考指导版(2008年7期)2008-12-19
