
蝙蝠飞行的空气动力学研究进展
2018-03-09 07:19余永亮中国科学院大学生物运动力学实验室北京100049
空气动力学学报 2018年1期
余永亮(中国科学院大学 生物运动力学实验室, 北京 100049)
0 引 言
蝙蝠是唯一可以飞行的哺乳动物,其种类繁多,几乎分布在世界上除南、北极以外的各个角落,其中又以热带地区的种类和数量最为丰富。在已发现的大约1000多种蝙蝠中,有3/4属于小型蝙蝠,重量在2 g到250 g之间;而大型蝙蝠身形较大,体重可达1.5 kg,双翼张开甚至可达2 m[1]。蝙蝠的生活习性比较独特且多样化,栖息时身体总是倒挂着,有的在山洞、地洞或建筑物的缝隙内,有的在树上或岩石上。
约70%的蝙蝠以昆虫为食,适应夜间飞行;还有以植物的果实为食的,它们大多白天出没。不同种类的蝙蝠飞行特性也不一样,比如犬吻蝠(Tadarida)善于持久的快速直线飞行,白长舌蝠(Leptonycteri)擅长中速机动飞行,而鼠耳蝠(Myotis)喜欢低速机动飞行[2]。
蝙蝠的广泛分布和多样性与其生活习性和环境有关。考古学家发现,五千多万年前的蝙蝠化石构造与现代蝙蝠并没有太大不同[3],这说明蝙蝠的生活习性和形态早已形成,并且一直对环境具有良好的适应性。其中,高超的飞行技能是它们适应环境的关键,使得它们具有很大的灵活性和机动性[4]。比如,大多捕食昆虫的蝙蝠能够在茂密的植物间自由飞行,可以实现急速起飞和降落、悬停、爬升、转弯、滚转、侧滑等机动动作[3,5]。因此,对蝙蝠飞行进行研究不仅可以增强人们对蝙蝠生理结构、形态适应及进化等生物学方面的认识,还有助于新概念可变形微型飞行器的设计[6]。
从空气动力学的角度来看,蝙蝠飞行有哪些特点呢?不妨从蝙蝠与昆虫、鸟类的飞行特点谈起。(A) 昆虫个体相对较小,飞行速度较低,雷诺数小,但拍翼频率高(可达100 Hz),翼周围的流动非定常性强。近二十年来,人们对昆虫拍翼飞行的研究成果十分丰富,逐步揭示了高升力产生的机制,包括“clap-and-fling”的Weis-Fogh机制,以及翼挥拍的前缘涡“不失速”机制[7]、拍动行程初期的“快速加速”机制和行程末期的“快速上仰”机制等等[8];从涡动力学角度分析,高升力主要依赖翼挥拍的附加惯性效应、抑制前缘涡脱落和增强后缘涡脱落三大机制的综合作用[9-10]。(B) 鸟类的体型相对较大,飞行速度快,从而飞行雷诺数大;其翅膀拍动频率低(1~2 Hz),流动非定常性较小。正因其雷诺数大、非定常性小的特点,一直以来人们用准定常空气动力学的理论就解答了鸟类产生气动力的机理,比如,用沉浮运动的翼型来阐述鸟类产生推力的机制[11]。(C) 中小型蝙蝠与昆虫相比,体格大,拍动频率低得多(10 Hz的量级),非定常性不如昆虫强;与鸟类相比,它的飞行速度小(1~10 m/s),因此飞行雷诺数不如鸟类大[12]。以某种小型蝙蝠(Leptonycteris yerbabuenae)为例[13],其特征飞行速率为3~5 m/s,身体倾斜角为7°,拍动频率为10 Hz,挥拍角为80°,飞行雷诺数为104~105,这些飞行参数介于昆虫与鸟之间。通过计算,若同条件下采用固定翼飞行,升力和阻力系数分别为0.62和0.12,若使刚性翼挥拍起来,升、阻力系数分别为0.80和0.19[14-15]。而根据实验数据[13],计算得到飞行所需的升力系数高达2.6。因此,除了挥拍的机制外,蝙蝠一定有着独有的产生高气动力机制。
本文将对蝙蝠飞行的研究进行评述,特别集中在空气动力学的研究方面。根据研究手段的不同,首先简述前人在蝙蝠形态学、生理学和运动学等方面的研究,然后从气动力的角度来分析蝙蝠飞行的空气动力学,最后提出未来的研究方向。
1 在形态学、生理学和运动学等方面的实验观测
事实上,人们对蝙蝠飞行的研究由来已久。在不同的时期,人们利用当时的研究条件,分别从形态学、生理学、运动学等多方面来研究蝙蝠飞行。按照时间(或者科技发展水平)顺序,大致可以把相关研究分为三个阶段。
第一阶段,人们通过静态的形态学数据间接研究蝙蝠的飞行能力。在20世纪六、七十年代,限于活体观测蝙蝠飞行的困难,人们常通过测量蝙蝠形态学(质量、翼外形、翼面积)和生理学(胸肌、心脏)参量,并结合蝙蝠的取食行为和生活环境,间接地研究了蝙蝠的飞行能力与环境适应的关系[16-18]。在这一阶段,人们仅能用准定常空气动力学分析方法探索蝙蝠飞行[19],但这只适用于快速前飞的蝙蝠。
第二阶段,随着摄影技术的引入,人们开始测量蝙蝠拍翼的运动学量,研究蝙蝠飞行的运动学规律。比如,飞行轨迹、飞行速度、转弯能力、拍动频率、拍动周期、拍动幅度等[5,20-25]。进一步地,根据数据得出了拍动频率f与飞行速率V的关系f=14.8V-0.277,拍动平面角α与速率之间的关系α=44.8V0.297,等[20]。这时,人们也意识到,对于慢速和悬停飞行的空气动力学研究就必须考虑蝙蝠翼拍动的非定常效应[26]。
第三阶段,随着新世纪里的技术革新,高速摄影以及粒子成像测速仪(PIV)的应用,这才使得对蝙蝠扑翼过程中翼的变形规律和尾迹结构的实验测量成为可能。大部分测量工作由三个大学的研究团队完成:美国布朗大学的Swartz和Breuer团队,瑞典隆德大学的Hedenström和德国比勒费尔德大学的Winter联合团队。Swartz和Breuer团队在短鼻果蝠的翅膀上布置了20 多个标记点,采用高速摄影追踪这些点的轨迹,从而分析蝙蝠翼拍动过程中的动态变形[27]。而对变形描述更详尽的是Hedenström和Winter团队,他们[13,28]不仅给出了运动学参数(如拍动频率、拍动平面角等与蝙蝠飞行速度的关系),还给出了不同速度下,第五手指所在截面的弓形变形规律,给出了蝙蝠内翼面积随时间变化规律(外翼改变较小),还给出了内翼和外翼中心点处的迎角随时间变化规律,等等。进一步地,上述两个研究团队还通过PIV技术开展了蝙蝠飞行的流场结构研究。比如,重构的尾涡结构[13,29-33],悬停蝙蝠下方形成的一系列的涡环堆积结构[33],甚至前缘涡结构(LEV)[34]。这为揭示蝙蝠飞行的高升力和推力机制提供了可能。
在第一阶段和第二阶段,研究者们只能通过部分“静态”的数据来研究蝙蝠飞行,在第三阶段,人们开始重视蝙蝠飞行中翅膀的动态变形了。但当前调研的结果看,由于蝙蝠翼构造和变形的复杂性,目前对蝙蝠翼变形的研究处于起步阶段[35]。
2 蝙蝠飞行的空气动力学研究
对飞行中蝙蝠翼如何产生气动力的研究一直没有停歇过。正如上小节中提到的第一阶段和第二阶段,人们发现用传统的准定常空气动力学只能分析大型蝙蝠的飞行,而对于中小型蝙蝠的飞行却需要考虑非定常效应。随着第三阶段的实验数据逐渐丰富起来,对于蝙蝠飞行已有来部分结论,这些结论有共性也有差异。共同的结论有三个:中小型蝙蝠拍翼产生的涡结构比鸟类复杂;蝙蝠翼下拍过程能产生正升力;蝙蝠中低速飞行时,必须考虑非定常性的影响。然而,在高升力和推力机制上达不成共识。有的认为与下拍过程的起动涡和终止涡产生高升力[12],有的认为是由蝙蝠翼下拍时翼尖涡和反向的根部涡导致的高升力[30],有的认为是前缘涡导致的[34];有的认为上挥行程可以产生推力[32],有的认为是阻力[36]。为了厘清蝙蝠飞行高气动力的机制,需要考虑特殊构造的蝙蝠翼在挥拍中的大变形效应[27,37]。
那么,蝙蝠翼在飞行中到底有哪些变形?这些变形对气动性能有什么样的影响?影响的机理是什么?当机动飞行需要增升或增推时,蝙蝠应该如何控制翼的变形?柔软翼膜的被动变形在其间又扮演什么样的角色呢?上述问题,部分已经有了初步答案,更多的还尚待解答。
2.1 蝙蝠翼的特征
在研究蝙蝠飞行的空气动力之前,得先了解蝙蝠翼的主要特征,然后总结变形规律。在此,我们根据研究需要,总结出蝙蝠翼的三个主要特征。
1) 蝙蝠翼与身体侧边相连,从脖子一直连到脚踝处,是“翼身一体化”构造。这种构造限制了蝙蝠翼的某些自由度。蝙蝠翼拍动时,不能像鸟类和昆虫一样自由旋转。如果想实现类似昆虫和鸟类的仰旋/俯旋运动,蝙蝠就必须扭转其翅膀,产生扭转变形(翼展向不同截面的迎角不同)。在飞行时,蝙蝠脚上下摆动,还能够影响翼的有效迎角,这是昆虫和鸟类不能做到的。
2) 多骨骼和关节结构。蝙蝠的骨骼结构跟其他哺乳动物没有本质上的不同,但前肢发生了适应性的变化。蝙蝠的前肢如同人类的前肢,骨骼包括非常强壮的肱骨,伸长的尺骨和桡骨,六枚腕骨、五枚掌骨和相应的五套指骨。其中,除了拇指还保留独立的活动性外(蝙蝠栖息倒钩就依靠拇指的抓握能力),其他四枚掌骨和指骨都极度伸长,形成翼膜的主体支架结构[27]。在肱骨、桡骨以及手指骨上,都附有健壮的肌肉,包括肱二头肌、肱三头肌和腕尺屈肌等[18]。多骨骼就意味着多关节,蝙蝠翼上有20多个关节[18,27,38],如肩关节、肘关节、腕关节、指关节等。
3) 翼膜结构。骨骼结构是蝙蝠翼的支撑框架,膜结构则构成了蝙蝠翼的受力主体。翼膜包括:位于腰部和第五手指间的体侧翼膜(Plagiopatagium),肩部和腕骨间的前翼膜(Propatagium),第五手指和翼尖支撑的手翼膜(Dactylopatagium),以及腿部和尾巴支撑的尾翼膜(Uropatagium)[39],如图1所示。翼膜又具有几个显著的特点。第一,非常薄,非常软。根据文献显示,翼膜厚度与弦长比在0.2%左右,杨氏模量在1 MPa的量级[39]。第二,表现为很明显的各向异性。蝙蝠翼膜由横纵纤维构成,不同部位不同方向的纤维分布不一,这就导致了翼膜材料各向异性的特点[39-41]。有些部位,弦向刚度是展向刚度的250倍;某些部位展向与弦向的刚度比达到1000∶1[41-42]。第三,作为生物材料,翼膜是粘弹性材料。

图1 蝙蝠翼多骨骼、多关节、柔性膜结构。图片来自Taylor et al. (2012)[35]Fig.1 The structure of the bat wing, including bones, joints and flexible membrane[35]
总之,从蝙蝠翅膀结构来看,蝙蝠翼既不同于大多数昆虫的轻质“膜+脉”结构,也不同于鸟类的前肢“羽翼”结构,而是由上肢骨骼和翼膜构成的“翼身一体化”构造(翼膜从脖子一直连接到脚踝)。由于蝙蝠的手指长、多关节,自由度可多达30个[38],且各向异性的翼膜薄且软,所以蝙蝠在拍动飞行中(特别是小型蝙蝠的中低速飞行),翼的变形极大,包含了骨骼控制的主动大变形和翼(膜)的被动变形。
2.2 蝙蝠翼的运动和变形规律
为了研究蝙蝠翼拍动时的空气动力特性,首先应该找出翼有哪些变形,然后建立变形的模型。
Hedenström和Winter团队通过实验测量了名为Leptonycteris yerbabuenae一种长鼻蝙蝠的翼在飞行的运动学和变形数据[13,28]。蝙蝠被控制在风洞中飞行,飞行速度在0~7 m/s,研究人员详细记录了蝙蝠翼挥拍过程中的三维运动学数据,以及翼的面积和升力系数。结果发现,蝙蝠翼的面积、迎角和弦向弓形变形都是提高升力的决定因素,而且它们随着飞行速度的提高而减小。弓形变形沿着展向变化,其主要受到身体和腿的弯曲,第5根手指(图2中的第4点到第7点的连线)的弯曲,翼前缘的挠曲和翼尖的弯曲。这些测量结果还表明蝙蝠翼在拍动中受到主动变形或者气动弹性的控制。下拍过程中,用来描述非定常性的Strouhal数保持相对恒定,这表明在研究的速度范围内,在下拍行程中保持良好的流动控制能力。鉴于此详细的三维运动学数据,以及文中提及的特征点的运动规律,Guan和Yu[14-15, 43]提出了四种变形方式,并给出了相应的数学模型,如图3所示。
1) 扭转变形,即迎角从翼根到翼尖线性分布,翼根迎角与身体倾斜角相同,翼尖迎角周期性变化,
αtip(t)=αtip0+αAcos(ωt+φα)
(1)
其中,αtip0表示翼尖的平均迎角,与翼根部的迎角(身体的倾斜角)是不等的,αA是翼尖迎角变化的幅度,φα是初始相位,ω为挥拍频率。

图2 蝙蝠翼的几何示意图。对三角形(Tr)用来测量迎角,臂部(armwing)和手部(handwing)用来定义展向变形。图片来自Wolf et al. (2010)[28]Fig.2 Schematic drawing illustrating the definition of wingbeat kinematicsparameters. Triangles (Tr) used forangle of attack analysis and the definition of armwing and handwing used for spanratio calculations[28]
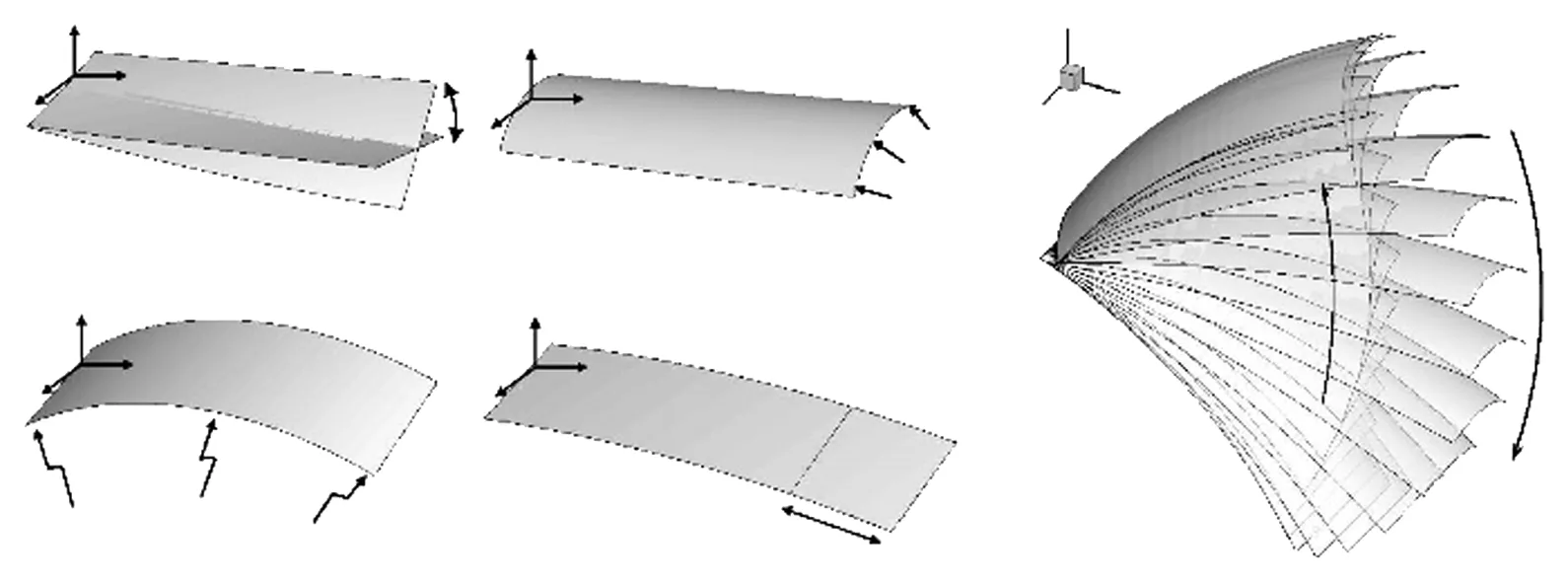
图3 模型翼的变形示意图[14]Fig.3 The morphing-wing model given by Yu and Guan[14]
2) 弓形变形,即由手指控制的弦向弓形弯曲。为了描述方便,把弓形变形近似成圆弧,弓形高度是实验测量的物理量。弓形高度与弦长之比的变化规律可以近似为
fc(t)=Ac+Ac1sin(ωt+φc1)+Ac2sin(2ωt+φc2)
(2)
其中的参数由实验进行标定[43]。
3) 展向弯曲变形,下拍的变形量小于上挥。同样地,展向弯曲变形用圆弧近似,其圆心角的变化规律为

(3)
4) 由于手指张合导致的翼膜面积变化。可以简化为周期性的简谐变化,即
S(t)=S0[1+Assin(ωt+φs)]
(4)
同样,各参数可以由实验数据标定。
虽然Hedenström等测量了不同速度下的挥拍模式,但为了简化,Guan和Yu[43]选择了中等速度 (3 m/s)下的数据进行挥拍建模,把挥拍运动看作简谐运动,即挥拍角满足
θ(t)=θ0+θAsin(ωt)
(5)
其中蝙蝠翼的挥拍参数也同样可以通过观测得到。如果需要非对称的拍动,可以借鉴关于昆虫拍动的模型[9],相应的变形模型也需要根据实验数据重新修订。
2.3 蝙蝠翼主动变形下的气动响应
2015年,Hedenström& Johansson在综述文章中谈到[44],虽然蝙蝠飞行的运动学和气动性能间有直接的关系,但仍缺少对蝙蝠翼形态结构和气动控制的认识。在von Busse等[13]给出的结果来看,升力的确受到变形的影响,为了确定到底哪类变形对升力和推力的影响大小,需要对每一类变形进行气动分析。Guan和Yu[15,43]用面元法研究了展弦比为3的矩形平板在四种变形方式下的气动性能响应,结果表明,升力主要依赖于弓形变形的大小和扭转变形的平均值αtip0,前者是上挥和下拍中有效迎角的非对称效应,后者的增大相当于蝙蝠在翼展方向的飞行迎角提高;推力主要依赖周期性的扭转变形,这类似于昆虫挥拍的俯仰机制,即从尾缘脱出的尾涡控制;而展向弯曲变形和翼膜面积变化对升力和推力起着放大的作用。综合变形得到的结果表明,升力和推力主要由下拍过程产生,上挥过程的气动力几乎可以忽略,如图4所示。

图4 一个周期内的升、阻力系数变化规律[14]Fig.4 Lift and drag generated by the integrated morphing wing during a whole stroke cycle[14]
与此同时,中国科学院力学研究所何国威课题组[45-46]和弗吉利亚理工大学的Tafti课题组[47]也对蝙蝠翼展向伸缩(面积变化)的气动性能也做了细致的研究。
2.4 模型翼主动变形的气动性能调控
在不同的环境、不同的状况下,蝙蝠需要产生不同的气动性能,以满足不同的飞行需求。比如蝙蝠要长距离匀速飞行,就需要产生恒定的升力(与重力平衡),阻力与推力相等(匀速)和能耗最少(最节省能量);蝙蝠遇到天敌或者需要捕食,需要在满足升力平衡体重的前提下,产生很大的推力,以获得大的加速度;蝙蝠进食或者怀孕,体重增加,就需要产生更大的升力,等等。那么蝙蝠是如何通过控制变形来满足这些需求呢?
为了回答这个问题,首先得知道每种变形对气动功率的影响,然后探讨气动控制的可能性。管子武在其博士论文[48]中给出了初步的数值研究结果。他发现,扭转变形幅值的增加可以提高平均推力,平均升力略微降低,而平均气动功率降低较大,整体上气动性能(此处定义为平均升力与平均功率之比)增强。弓形变形的增加,平均升力增加,平均推力也增加,气动功率也增加,整体上气动性能也增强。增加面积收缩率不仅可以提高平均升力,也可以增加平均推力,同时能耗也增加,整体气动性能也减弱。
进一步地,管子武[48]假定蝙蝠身体的阻力系数(假设为0.1),用计算手段寻找平稳飞行的状态,即满足升力平衡体重,推力平衡阻力。然后,分别找到推力平衡阻力时的升力调节机制和升力平衡体重时的推力调节机制,通过参数相图找到了条件最优的变形组合,即满足气动力条件下的最低气动功率输出的状态。这为变形优化提供了一种研究途径。
在未来的研究中,可以用遗传算法或者伴随优化算法等对蝙蝠翼动态变形进行多参数优化。
2.5 研究蝙蝠飞行还存在的困难和挑战
从现有的文献来看,对于蝙蝠飞行的空气动力学研究还并不充分,这主要归结于两方面的原因。一方面,对蝙蝠飞行中的翼变形和飞行模态之间的关系缺少详细的数据,也无法区分主动变形和被动变形,这不利于建立合理的数学模型;另一方面,研究手段有待提高,在测量上,不仅需要蝙蝠飞行时的详细运动学数据,还需要测量生理能耗、翼膜和骨骼的材料性质等数据。数值分析方面也遇到了不少的困难,比如几何建模、“骨骼-膜”结构如何划分网格,指尖的网格奇性处理、流固耦合计算等。
从空气动力学的研究内容看,还存在很多需要解答的困惑,比如,对于蝙蝠这中“翼身一体”化的飞行动物,如何区分左右翼的干扰,翼身之间的干扰,以及如何区分蝙蝠膜翼结构的被动变形特征及其气动响应,结合骨骼材料特性的流固耦合、飞行稳定性、自由飞行和机动飞行的控制等等,这些都等着更多研究者们的投入。
3 结 论
当前对蝙蝠飞行的研究已经有了较大的进展,除了对蝙蝠翼的变形和非定常气动力的测量手段有大的进步外,对飞行时翼的主动大变形产生气动力的机制也已有了初步的认识,但仍有很长一段路要走。首先,需要进一步明确蝙蝠飞行的“翼身一体”的升力、推力及力矩特性,探讨蝙蝠“翼-身”“翼-翼”之间的动态干扰;其次,需要区分蝙蝠翼膜在飞行中的被动变形和主动变形,判定被动变形对气动性能影响的大小;最后,探索蝙蝠飞行的稳定性及自由飞行和机动飞行的控制机制等等。
[1]Wilson D E, Reeder D M. Mammal species of the world: a taxonomic and geographic reference[M]. Johns Hopkins Unversity Press, 2005
[2]Struhsaker T T. Morphological factors regulating flight in bats[J]. Journal of Mammalogy, 1961, 42(2): 152-159
[3]Altringham J D, et al. Bats: biology and behaviour[M]. Oxford University Press Oxford, 1998
[4]Swartz S M, Konow N. Advances in the study of bat flight: the wing and the wind[J]. Canadian Journal of Zoology, 2015, 93(October): 977-990
[5]Norberg U M. Aerodynamics, kinematics, and energetics of horizontal flapping flight in the long-eared bat Plecotus auritus[J]. Journal of Experimental Biology, 1976, 65(1): 179-212
[6]Valasek J. Morphing aerospace vehicles and structures[M]. John Wiley & Sons, 2012
[7]Dickinson M H, et al. Wing rotation and the aerodynamic basis of insect flight[J]. Science, 1999, 284(18 June): 1954-1960
[8]Sun M. Insect flight dynamics: stability and control[J]. Reviews of Modern Physics, 2014, 86(2): 615-646
[9]Yu Y, Tong B. A flow control mechanism in wing flapping with stroke asymmetry during insect forward flight[J]. Acta Mechanica Sinica, 2005, 21(3): 218-227
[10]Yu Y, et al. An analytic approach to theoretical modeling of highly unsteady viscous flow excited by wing flapping in small insects[J]. Acta Mechanica Sinica, 2003, 19(6): 508-516
[11]Wu T Y. A review on fish swimming and bird/insect flight[J]. Annual review of fluid mechanics, 2011, 43: 25-58
[12]Hedenström A, et al. Bird or bat: comparing airframe design and flight performance[J]. Bioinspiration & Biomimetics, 2009, 4(1): 15001
[13]Von Busse R, et al. Kinematics and wing shape across flight speed in the bat, Leptonycteris yerbabuenae[J]. Biology open, 2012, 1(12): 1226-38
[14]Yu Y, Guan Z. Learning from bat: Aerodynamics of actively morphing wing[J]. Theoretical and Applied Mechanics Letters, Elsevier Ltd, 2015, 5(1): 13-15
[15]Guan Z, Yu Y. Aerodynamic mechanism of forces generated by twisting model-wing in bat flapping flight[J]. Applied Mathematics and Mechanics (English Edition), 2014, 35(12): 1607-1618
[16]Hartman F A. Some flight mechanisms of bats[J]. The Ohio Journal of Science, 1963, 63(2): 59-65
[17]Norberg U M, Rayner J M V. Ecological morphology and flight in bats (mammalia; chiroptera): wing adaptations, flight performance, foraging strategy and echolocation[J]. Philosophical Transactions of the Royal Society B, 1987, 316(1179): 335-427
[18]Norberg U M. Bat wing stuctures important for aerodinamics and rigidity (mammalia, chiroptera)[J]. Journal of Morphology, 1972, 73: 45-61
[19]Norberg U M L. Aerodynamics of hovering flight in the long-eared bat plecotus auritus[J]. Journal of Experimental Biology, 1976, 65(2): 459-470
[20]Norberg U M L, Winter Y. Wing beat kinematics of a nectar-feeding bat, Glossophaga soricina, flying at different flight speeds and Strouhal numbers[J]. Journal of Experimental Biology, 2006, 209(19): 3887-3897
[21]Watts P, et al. A computational model for estimating the mechanics of horizontal flapping flight in bats: model description and validation[J]. Journal of Experimental Biology, 2001, 204(16): 2873-98
[22]Britton A, et al. Flight performance, echolocation and foraging behaviour in pond bats, Myotis dasycneme (Chiroptera: Vespertilionidae)[J]. Journal of Zoology, 1997, 241(3): 503-522
[23]Aldridge H D. Turning flight of bats[J]. Journal of Experimental Biology, 1987, 128: 419-425
[24]Aldridge H D. Kinematics and aerodynamics of the greater horseshoe bat, Rhinolophus ferrumequinum, in horizontal flight at various flight speeds[J]. Journal of Experimental Biology, 1986, 126: 479-497
[25]Rayner J M V, Aldridge H D J N. Three-dimensional reconstruction of animal flight paths and the turning flight of microchiropteran bats[J]. Journal of Experimental Biology, 1985, 118(1): 247-265
[26]Norberg U N. Vertebrate flight: mechanics, physiology, morphology, ecology and evolution[M]. BioScience, 1990
[27]Swartz S M, et al. Wing structure and the aerodynamic basis of flight in bats[A]. AIAA paper[C]//45th AIAA Aerospace Sciences Meeting and Exhibit 8-11 January 2007, Reno, Nevada: 2007: 2007-42
[28]Wolf M, et al. Kinematics of flight and the relationship to the vortex wake of a Pallas’ long tongued bat (Glossophaga soricina)[J]. The Journal of Experimental Biology, 2010, 213(12): 2142-2153
[29]von Busse R, et al. The aerodynamic cost of flight in the short-tailed fruit bat (Carollia perspicillata): comparing theory with measurement[J]. Journal of the Royal Society Interface, 2014, 11: 20140147
[30]Hubel T Y, et al. Wing motion and wake structure of bat flight[J]. Comparative Biochemistry and Physiology A-molecular & Integrative Physiology, 2008, 150(3): s77
[31]Muijres F T, et al. Leading-edge vortex improves lift in slow-flying bats[J]. Science, 2008, 319(5867): 1250-1253
[32]Hedenstrom A, et al. Bat flight generates complex aerodynamic tracks[J]. Science, 2007, 316(5826): 894-897
[33]Hakansson J, et al. The wake of hovering flight in bats[J]. Journal of the Royal Society Interface, 2015, 12(109): 20150357
[34]Muijres F T, et al. Leading edge vortices in lesser long-nosed bats occurring at slow but not fast flight speeds[J]. Bioinspiration & Biomimetics, 2014, 9(2): 25006
[35]Taylor G K, et al. Wing morphing in insects, birds and bats: mechanism and function[A]. Morphing aerospace vehicles and structures[M]. John Wiley & Sons, Ltd., 2012: 11-40
[36]Muijres F T, et al. Comparative aerodynamic performance of flapping flight in two bat species using time-resolved wake visualization[J]. Journal of the Royal Society Interface, 2011, 8(63): 1418-1428
[37]Waldman R M, et al. Aerodynamic Behavior of Compliant Membranes as Related to Bat Flight[R]. AIAA paper 2008: 3716
[38]Bahlman J W, et al. Design and characterization of a multi-articulated robotic bat wing[J]. Bioinspiration & Biomimetics, 2013, 8(1): 16009
[39]Swartz S M, et al. Mechanical properties of bat wing membrane skin[J]. Journal of Zoology, 1996, 239(2): 357-378
[40]Swartz S M, et al. Dynamic complexity of wing form in bats: implications for flight performance[A]. Z. Akbar, et al. Functional and evolutionary ecology of bats[M]. Oxford University Press, 2006: 110-130
[41]Swartz S M. Skin and bones functional, architectural, and mechanical differentiation in the bat wing[A]. T.H. Kunz, P.A. Racey. Bat Biology and Conservation[M]. Boston University: Smithsonian Institution Press, 1998: 109-126
[42]Ray C W. Modeling, control, and estimation of flexible, aerodynamic structures[D]. PHD Thesis, Oregon State University, 2012
[43]Guan Z, Yu Y. Aerodynamics and mechanisms of elementary morphing models for flapping wing in forward flight of bat[J]. Applied Mathematics and Mechanics (English Edition), 2015, 36(5): 669-680
[44]Hedenstrom A, Johansson L C. Bat flight: aerodynamics, kinematics and flight morphology[J]. Journal of Experimental Biology, 2015, 218(5): 653-663
[45]Wang S, et al. Lift enhancement on spanwise oscillating flat-plates in low-Reynolds-number flows[J]. Physics of Fluids, 2015, 27(6): 61901
[46]Wang S, et al. Lift enhancement by dynamically changing wingspan in forward flapping flight[J]. Physics of Fluids, 2014, 26(6): 1-56
[47]Viswanath K, et al. Straight-line climbing flight aerodynamics of a fruit bat[J]. Physics of Fluids, 2014, 26(2): 21901
[48]管子武. 蝙蝠平飞时其模型翼主动大变形气动响应及被动变形效应研究[D]. 中国科学技术大学, 2014.
猜你喜欢
天然气与石油(2022年4期)2022-09-21
北京航空航天大学学报(2021年6期)2021-07-20
北京航空航天大学学报(2021年6期)2021-07-20
燃气涡轮试验与研究(2019年2期)2019-05-18
空气动力学学报(2019年2期)2019-05-08
科学Fans(2019年4期)2019-04-30
劳动保护(2018年5期)2018-06-05
无人机(2017年10期)2017-07-06
中学课程辅导·教师教育(中)(2017年2期)2017-03-24
少年科学(2014年7期)2014-08-18
