
利用微卫星分子标记研究东方田鼠的多父权现象
2018-03-08 07:53:50郭宗明
生物学杂志 2018年1期
郭宗明, 王 勇, 郭 聪
(1. 烟台南山学院, 烟台 265713; 2. 四川大学 生命科学学院, 成都 610064; 3. 中国科学院 亚热带农业生态研究所, 长沙 410125)
动物行为是在一定环境中,为满足生存、繁衍后代等生理需要而完成的诸多动作[1]。而繁殖行为是动物诸多行为中极为重要的行为。通过复杂的繁殖行为,保证繁殖成功,才能维持种族延续和发展。婚配制度是动物种群中的个体在繁殖期为获得配偶而采取的普遍的行为对策,其包括以下几种含义[2]:1)个体获得配偶数量;2)两性是否有双亲行为;3)配对关系的紧密程度及持续时间。依据动物个体在一个繁殖季节里与其交配的配偶数,婚配制度分为单配制和多配制,其中多配制又分为一雄多雌制,一雌多雄制以及混交制3种类型。
田鼠亚科动物(Microtine)的婚配制度差异较为明显,是研究婚配制度的理想动物[3]。运用活捕和无线电遥测技术的研究表明:草原田鼠(Microtusochrogaster)雌雄共巢且巢面积大致相同,显示单配制特征[4]。邰发道等[5]通过实验室内的观察,结合野外生态研究的资料分析后认为棕色田鼠(M.mandarinus)也具有单配制特征。泰加田鼠(M.xanthognathus)雌雄鼠很少共巢,雌雄巢面积差异显著,雄性和多个雌性巢重叠,显示一雄多雌特征。而山地田鼠(M.montanus)雌雄巢的特点大致同泰加田鼠,但雌雄巢区相互重叠,显示混交制特征[6]。
通过野外和室内对啮齿类动物的观察来推测其婚配制度有着一定的局限性。例如,Getz等[7]依据草原田鼠(M.ochrogaster)的繁殖排他性、领域、单配行为以及双亲关心行为推断该鼠为单配制型。然而,分子标记的分析表明草原田鼠56%的窝仔中存在多父权现象[8]。因此,分子生物学的技术优势解决了传统方式不能解决的问题。具有高度变异的分子遗传标记,如微卫星,已被广泛用于动物婚配制度的研究[9]。
栖息于洞庭湖区的东方田鼠是近些年来新兴的危害性啮齿动物。郭聪等[10-11]根据野外和室内的行为观察推测该鼠的婚配制度是一夫一妻制的可能性极小,为混交制的可能性较大。由于野外不能真实看到东方田鼠的交配行为,室内观察又受到多种因素的影响,因此,有必要运用分子遗传标记的方法进一步探究和证实东方田鼠的婚配行为。
1 材料与方法
1.1 样品采集
以葵花籽为诱饵,采用鼠夹捕获了7只怀孕雌鼠。取母鼠腿部肌肉及其胚胎放入酒精中,编号记录。回实验室后冷冻保存。
1.2 试剂及仪器
PCRMix、PCR引物(上海生工合成)、聚丙烯酰胺、硝酸银、尿素、甲叉丙烯酰胺、TEMED、过硫酸铵。
PCR仪、紫外分光光度计、离心机、凝胶成像系统(Bio-Rad)、垂直电泳仪(北京六一)。
1.3 DNA提取
取少许肌肉组织、胚胎组织,用双蒸水清洗2次后,运用酚-氯仿法[12]完成抽提DNA过程。用紫外分光光度计检测提取的DNA质量。
1.4 引物来源及筛选
本研究使用与东方田鼠亲缘关系较近的属种的微卫星引物以及GenBank中的微卫星序列自行设计引物进行扩增。所有引物经优化、扩增、琼脂糖凝胶电泳检测,选择条带单一、且稳定扩增的6对引物为下一步使用的引物(表1)。
1.5 PCR体系
用筛选的引物分别对母鼠及其胚胎的DNA进行PCR扩增。根据PCRMix,设计PCR扩增体系为15 μL(包括约100 ng模板DNA,0.3 mmol/L dNTP,上下游引物各0.24 mmol/L,30 mmol/L KCl,6 mmol/L Tris-HCl,0.9~1.2 mmol/L MgCl2,0.75 UTaqDNA 聚合酶)。PCR反应程序为94℃预变性4 min;94℃变性40 s,54℃~63℃退火温度(依据每对引物而定)50 s,延伸1 min,完成30个循环反应;最后延伸10 min结束。取PCR产物各10 μL用1%琼脂糖凝胶电泳检测扩增情况。
1.6 聚丙烯酰胺凝胶制备
将配套的玻璃板放入NaOH溶液中浸泡一昼夜,然后冲洗后晾干备用。将晾干的玻璃板放平,用酒精擦3次。无耳玻璃板用亲和硅烷擦2次,有耳玻璃板用剥离硅烷擦2次。将隔板放在两玻璃板之间,然后固定。用水平仪将玻璃板调节呈水平状态。再将配制好的凝胶液[丙烯酰胺和甲叉丙烯酰胺混合液(45%),10×TBE,1.6%过硫酸铵]徐徐灌入玻璃板中。胶灌好后,小心插入梳子,放置1.5~2 h,凝胶即可制备好。

表1 微卫星引物及重复序列,退火温度,等位基因数
1.7 PAGE电泳、银染
1.7.1 电泳
将制备好的凝胶放入竖直电泳槽,固定。然后加入1×TBE缓冲液,预电泳3 h。将变性的PCR产物取5 μL 点样,最后点上PUC18 Marker,1600 V,50 W状态下电泳3 h。
1.7.2 银染
电泳完毕后将胶与玻璃板放入固定液中固定20 min,然后用双蒸水漂洗2次,再放入硝酸银溶液中染色约15 min。染色结束后漂洗2次,放入显色液中,观察条带清晰时取出漂洗2次,拍照保存。
1.8 微卫星分析
将所有样品在同一凝胶上电泳、银染、显色。观察每一母鼠与其胎仔在每个位点上的条带情况,若判断有多父权的可能,再重复一次PCR、电泳、银染、显色过程,保证判断的准确性。运用Labwork软件进行等位基因大小的判断。
1.9 多父权分析
对每一母鼠的等位基因和其胎仔的等位基因进行比较,判断每一窝胎仔其可能父亲的等位基因数,进而判断父权数。若每一窝胎仔中雄鼠的等位基因超过2个,证明此窝仔拥有多父权现象。
2 结果
本研究中每窝平均窝仔数为5.29±1.60,除2窝胎仔的父权数明显超过一个外,其他窝仔的父权数判断至少为1个(表2)。
图1 显示,03M组母鼠有A、B等位基因,是杂合子,胎仔04个体与母鼠相同。01、03、05和06 个体都有一个不同于母鼠的等位基因C,而02个体有不同于母鼠的等位基因D,表明02个体与01、03、05和06有不同的父权,因此判断03组胎仔中至少拥有2个父权。
图2 显示,07M组母鼠有A、B等位基因,是杂合子,胎仔06个体与母鼠相同。02、04和05个体有一个不同于母鼠的等位基因C,而01个体拥有不同于等位基因A、B、C的等位基因基因D。由于母鼠拥仅有A、B 2个等位基因,而胎仔个体中出现与母鼠不同的等位基因C和D。01和02分别拥有一个不同于母鼠的等位基因C和D,表明01和02个体拥有不同的父权,06个体拥有A、B两个等位基因,其父权与01、02个体不同,因此可判断07M组胎仔中至少拥有3个父权。

表 2 7组胎仔的父权分析结果
图 3中,06M组母鼠有A、B 2个等位基因,母鼠是杂合子。02、05和06胎仔个体也是杂合子,而01、03和04胎仔个体都只有等位基因B,是纯合子。来自雄性鼠的等位基因有2个,推断06M组胎仔中至少拥有1个父权。
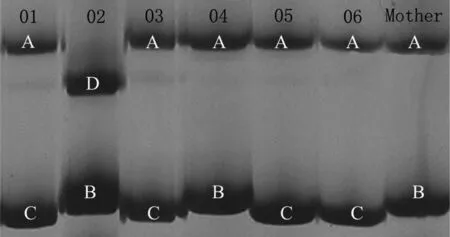
图1 Moe4位点引物扩增03M组母鼠与其胎仔的电泳图
字母表示每个个体的等位基因

图 2 Ma11位点引物扩增07M组母鼠与其胎仔的电泳图
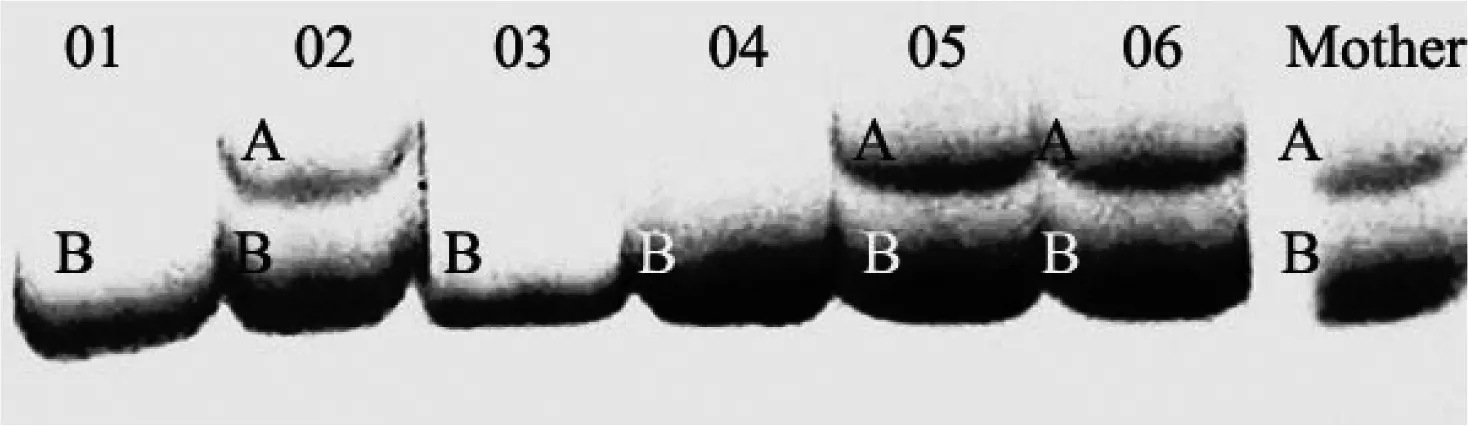
图 3 Moe4位点引物扩增06M组母鼠与其胎仔的电泳图
3 讨论
Jennion和Petrie[14]综述了多父权带给子代的遗传优势:多父权不仅增加子代遗传多样性,还增加选择上的适合度和繁殖成功率。多父权也影响有效种群的大小,多父权比单父权能增加种群大小,是保证种群多样性降低近交繁殖的一种策略。这些理论研究可以解释东方田鼠具有多父权婚配行为的生物学意义。洞庭湖每年有季节性洪水发生,形成了盛水期与枯水期交替出现的景观变化格局。伴随这种变化,东方田鼠也周期性在湖滩和垸内农田、岗地之间往返迁移。迁移至农田的东方田鼠,受到夏季高温及迁移时体力消耗过大的影响,表现出很低的繁殖力[15]。不同生境中,东方田鼠的繁殖力差别较大。避难于稻田、岗地的东方田鼠其繁殖强度远低于湖滩生境。郭聪等[16]研究洪水期东方田鼠的迁移机理,发现不同年份迁入垸内的东方田鼠的雄性比例差异显著。当湖水位下降,湖滩逐渐显露时,东方田鼠回迁湖滩。回迁时雄性的回迁比率大于雌性,且个体较大。洞庭湖特殊景观生境周期性变更、不同年份雄性比的显著差异及洪水期种群数量的减少可能是东方田鼠形成混交制的外部影响因素。东方田鼠采取混交婚配制度可能是适应湖滩生境的一种繁殖行为策略。
如图3所示,06M组中的窝仔拥有来自父权的2个等位基因,但不能排除是多父权的可能,因为多个不同的父亲可以传递相同的等位基因[17]。另外,同一物种种群随环境条件的差异多父权也有所变化,种群密度和生境结构的不同也影响婚配行为[18-19]。分布在洞庭湖和长沙市区的东方田鼠在种群密度和生境结构上存在着差异,这些差异是否影响着洞庭湖和长沙东方田鼠的婚配行为有待进一步研究探讨。此外,本研究在微卫星位点的选择上仅选择扩增条带单一的位点,没有研究微卫星位点的多态性情况。因此,对洞庭湖东方田鼠婚配行为上表现的多父权行为的研究有待进一步深入。
[1]蒋志刚, 李春旺, 彭建军, 等. 行为的结构、刚性和多样性[J].生物多样性, 2001, 9(3): 265-274.
[2]TRIVERS R L. Parental investment and sexual selection [M]. Chicago: Aldine, 1972: 139-179.
[3]WANG Z X, INSEL T R. Parental behavior in voles [J]. Advances in study of behavior, 1996, 25:361-384.
[4]GULIN S J C, FITZGERALD R W. Home range size as a predictor of mating system in Microtus [J]. Journal of Mammalogy, 1988, 69(2): 311-319.
[5]邰发道, 王廷正, 赵亚军. 棕色田鼠的配偶选择和相关特征[J].动物学报,2001, 47(3): 266-273.
[6]WOLFF J O, LIDICKER W Z L JR. Communal winter nesting and food sharing in Taiga Voles[J]. Behavioral Ecology and Sociobiology, 1981, 9(4): 237-240.
[7]GETZ L L, CARTER C S, GAVISH L. The mating system of the prairie vole,Microtusochrogaster: field and laboratory evidence for pair-bonding[J]. Behavioral Ecology and Sociobiology, 1981, 8(3):189-194.
[8]SOLOMON N G, KEANE B, KNOCH L R, et al. Multiple paternity in socially monogamous prairie voles(Microtusochrogaster) [J]. Canadian Journal of Zoology, 2004, 82(10):1667-1671.
[9]THONHAUSER K E, THOβ M, MUSOLF K, et al. Multiple paternity in wild house mice(Musmusculusmusculus): effects on offspring genetic diversity and body mass[J]. Ecology and Evolution, 2014, 4(2): 200-209.
[10]郭 聪. 东方田鼠的杀幼行为及其繁殖对策研究[D]. 北京:中国科学院动物研究所, 1998: 30-150.
[11]郭 聪, 王 勇, 张美文, 等. 洞庭湖区东方田鼠洞群成员分析[J].兽类学报, 2001, 21(1): 44-49.
[12]KEANE B, WASER P M, CREEL S R, et al. Subordinate reproduction in dwarf mongooses[J]. Animal Behaviour, 1994, 47(1): 65-75.
[13]VAN DE ZANDE L, VAN APELDOORN R C, BLIJDENSTIN A F, et al. Microsatellite analysis of population structure and genetic differentiation within and between populations of the root vole,Microtusoeconomus, in the Netherlands[J]. Molecular Ecology, 2000, 9(10): 1651-1656.
[14]JENNION M D, PETRIE M. Why do females mate multiply? A review of the genetic benefits [J]. Biological reviews of the Cambridge Philosophical Society, 2000, 75(1): 21-64.
[15]郭 聪, 张美文, 王 勇, 等. 洞庭湖区夏季温光条件及被迫迁移对东方田鼠繁殖的影响[J].兽类学报, 1999,19(4): 298-307.
[16]郭 聪, 王 勇, 陈安国,等.洞庭湖区东方田鼠迁移的研究[J].兽类学报, 1997,17(4): 279-286.
[17]BURTON C. Microsatellite analysis of multiple paternity and male reproductive success in the promiscuous snowshoe hare[J]. Canadian Journal of Zoology, 2002, 80(11): 1948-1956.
[18]SAY L, PONTIER D, NATOLI E. High variation in multiple paternity of domestic cats(FeliscatusL.) in relation to environmental conditions[J]. Proceedings of the Royal Society of London. Series B. Biological sciences, 1999, 266(1433):2071-2074.
[19]MODOROV M V. The variability of allozyme and microsatellite loci of the narrow-headed voleLasiopodomysgregalisfrom the Southern Urals and Trans-Urals[J]. Biology Bulletin, 2016, 43(1):46-51.
猜你喜欢
意林·少年版(2022年2期)2022-03-09 00:36:42
名作欣赏·评论版(2020年3期)2020-03-01 01:19:17
中国动物保健(2019年3期)2019-12-11 18:26:23
文学教育(2019年10期)2019-11-07 11:38:48
卷宗(2019年13期)2019-06-17 01:22:38
发明与创新·小学生(2019年5期)2019-06-14 08:33:03
发明与创新(2019年19期)2019-05-20 11:40:56
阅读(科学探秘)(2017年4期)2017-05-30 10:48:04
农家之友(2017年5期)2017-03-27 09:28:56
饲料博览(2016年6期)2016-04-05 14:11:14
