
渤海湾口虾蛄假蚤状幼体的密度分布及影响因素研究
2018-03-07 08:49谷德贤王婷王娜王刚尤宏争李文雯刘国山
大连海洋大学学报 2018年1期
谷德贤,王婷,王娜,王刚,尤宏争,李文雯,刘国山
(1.天津市水产研究所,天津300221;2.天津渤海水产研究所,天津300457)
口虾蛄Oratosquilla oratoria隶属于节肢动物门Arthropoda、甲壳纲Crustacea、口足目Stomatopoda、虾蛄科Squillidae、口虾蛄属Oratosquilla[1],俗称虾爬子、螳螂虾、虾虎、琵琶虾、虾拔弹等[2]。近年来,近海渔业资源衰退明显,在各类渔业经济品种衰退的同时,口虾蛄的价值不断体现出来。2011—2015年期间,中国的口虾蛄产量一直维持在29万t左右[3-7],已经成为近岸主要的渔业经济品种之一。
国外关于口虾蛄已有较多的研究,Provenzano等[8]研究了虾蛄的养殖方法,大富潤等[9]研究了虾蛄产卵场的特征和产卵动态,Hamano等[10-13]进行了虾蛄孵化、幼虫发育等各方面的研究。同时,国内也开展了大量的口虾蛄研究工作,主要有口虾蛄基础生物学[14-15]、 形态学[16-17]、 生理学[18-20]、发育生物学[21]、人工繁育养殖[22-24]、资源学[25-27]、分子生物学[28-29]、 生物制药[30]研究等。而关于自然海域口虾蛄假蚤状幼体的研究目前仅见Hamano等[31]的报道。
口虾蛄到达繁殖季节后,首先产卵、抱卵,然后假蚤状幼体破膜而出,经历11期假蚤状幼体的浮游阶段,发育为仔虾,转为底栖性生活。本研究中,从自然海域口虾蛄假蚤状幼体的资源密度及环境影响因子两个方面开展研究,分析了口虾蛄早期生活史的规律和环境影响因素,并辅助分析了口虾蛄的繁殖习性,旨在为口虾蛄渔业资源开发和保护提供技术支撑,为政府管理部门提供决策依据。
1 材料与方法
1.1 数据来源
本研究数据来源于天津市水产研究所2015—2016年5、6、7、8、10月进行的10个航次口虾蛄假蚤状幼体及同步进行的环境因子调查资料。调查范围为渤海湾,调查船主机功率为176 kW,船长为24.88 m,型宽为5 m,型深为2.2 m,总吨位为64 t。假蚤状幼体调查采用大型浮游生物网(网口直径为0.8 m,网口面积为0.5 m2,网长为2.8 m,筛绢孔径为0.507 mm)水平拖网,每站拖网大约10 min,网口系有流量计。在每个站点,同
步测定表层温度、表层盐度、水深、溶解氧、营养盐、叶绿素a等理化因子。水质的检测分析参照《海洋调查规范》(GB12763—2007)[32]的相关要求进行。
1.2 方法
1.2.1 资源密度估算 口虾蛄假蚤状幼体的资源量密度计算公式为
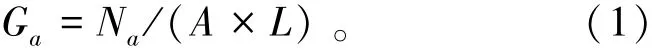
其中:Ga为单位面积海水中口虾蛄假蚤状幼体的个体数 (ind./m2);Na为全网假蚤状幼体的个体数(ind.);A为网口直径 (m);L为拖网距离 (m),根据流量计计算。
1.2.2 环境影响因子分析 应用广义加性模型(Generalized additive model,GAM)分析口虾蛄假蚤状幼体与影响因素间的关系,具体计算过程参见谷德贤等[33]的方法。用作GAM分析的预测变量为月份 (MO)、经度 (LON)、纬度 (LAT)、水温(SST)、水深 (WD)、盐度 (S)、溶解氧 (DO)、无机氮 (DIN)、活性磷酸盐 (DIP)、化学耗氧量(COD)、叶绿素a(Chl-a),响应变量为口虾蛄假蚤状幼体的资源密度 (ind./m2)。月份、经度和纬度可看作分类变量,也可看作数值变量,本研究中按上述两种情形计算GAM,并进行比较,得出最优模型。通过校正后决定系数的变化量比较各预测变量的相对重要性,计算方法见谷德贤等[33]的研究,越大则相对重性越大。
2 结果与分析
2.1 口虾蛄假蚤状幼体的密度分布
2015年和2016年进行10个航次的调查,共采集到口虾蛄假蚤状幼体4734尾,平均密度为0.095 ind./m2(表 1)。其中,2015年 2354尾,平均密度为0.120 ind./m2;2016年2383尾,平均密度为0.068 ind./m2,低于2015年的平均密度。5月和10月航次均未采集到口虾蛄假蚤状幼体。2015年6—8月,口虾蛄假蚤状幼体的密度逐月增大,8月达到最大值,为0.478 ind./m2。2016年6—8月,口虾蛄假蚤状幼体的密度先减后增,其中6月的密度最大,为0.169 ind./m2,7月最低,为0.001 ind./m2。
2.2 资源密度的环境影响因子
2.2.1 预测变量分析 对预测变量MO、LON、LAT、 SST、 WD、 SSS、 DO、 DIN、 DIP、 COD 和Chl-a绘制箱形图,其中,DIP的离散值偏离较大,为减少异常值对模型的影响[34],对DIP进行平方根转换。经方差膨胀因子 (Variance inflation factor,VIF)检验,所有预测变量的VIF均小于阈值,均可用于后续的GAM分析。资源密度分布如图1所示,近似正态分布,响应变量的分布类型设为正态分布,连接函数为Identity link。
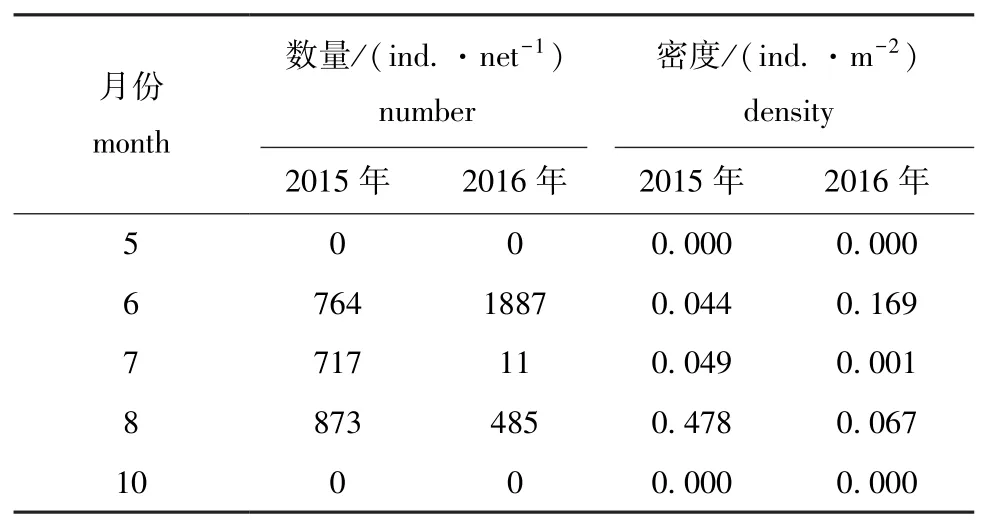
表1 口虾蛄假蚤状幼体的密度分布Tab.1 Population density of mantis shrimp Oratosquilla oratoria pseudozoea

图1 口虾蛄假蚤状幼体密度分布图Fig.1 Distribution curve of the population density of mantis shrimp Oratosquilla oratoria pseudozoea
2.2.2 GAM分析 MO、LON和LAT即可看作数值变量,又可看作分类变量,本研究中按上述两种情形计算GAM,并进行比较,得出最优模型。
(1)基于分类变量的 GAM分析。将 MO、LON和LAT视为分类变量,对口虾蛄假蚤状幼体密度和各预测变量进行逐步运算,得到最优GAM模型的表达式为
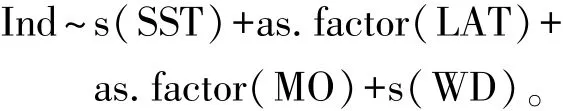
从表2可见:纬度在38.67°N和38.83°N处时对幼体密度有极显著性影响 (P<0.01),在39.10°N处时有显著性影响 (P<0.05);从月份来看,6月、7月和8月对口虾蛄幼体密度均有极显著性影响 (P<0.01);水温对幼体密度有极显著性影响 (P<0.01),水深对幼体密度有显著性影响(P<0.05)。根据校正后决定系数的变化量, 4个预测变量的相对重要性依次为水温 (0.224)>月份 (0.178)>纬度 (0.143)>水深 (0.111)。

表2 GAM最优模型中各预测变量的显著性Tab.2 Significance of the predictors in GAM
从图2可见:从水温来看,表层水温对幼体密度的影响明显,在15~18℃时,其对幼体密度的影响随着表层水温的增加而增加,在18℃左右时影响最大,在23~27℃时,主要表现为水温与幼体密度呈负相关,在25℃左右时负相关最大,30℃左右时水温对幼体密度的影响较小;从纬度来看,以38.50°N为基准,在38.67°N和38.83°N处时纬度对幼体密度的影响较大,其次是39.10°N,均与幼体密度呈负相关,39.00°N的数据离散程度最大;从月份来看,以5月为基准,6月、7月和8月对幼体密度的影响较大,10月影响较小;从水深来看,水深对幼体密度的影响主要在11.5 m以浅,在11.5~20.0 m时水深的影响趋于平稳,在5.0~7.0 m时水深对幼体密度的影响为负相关,随着水深的增加其对幼体密度影响逐渐减少,然后变化曲线下降后又再次上升,在10.5 m时达到最大值。

图2 口虾蛄假蚤状幼体密度与各预测变量间的关系 (纬度和月份设为分类变量)Fig.2 Relationship between the popultion density of mantis shrimp Oratosquilla oratoria pseudozoea and predictor variables(latitude and month are treated as categorical variables)
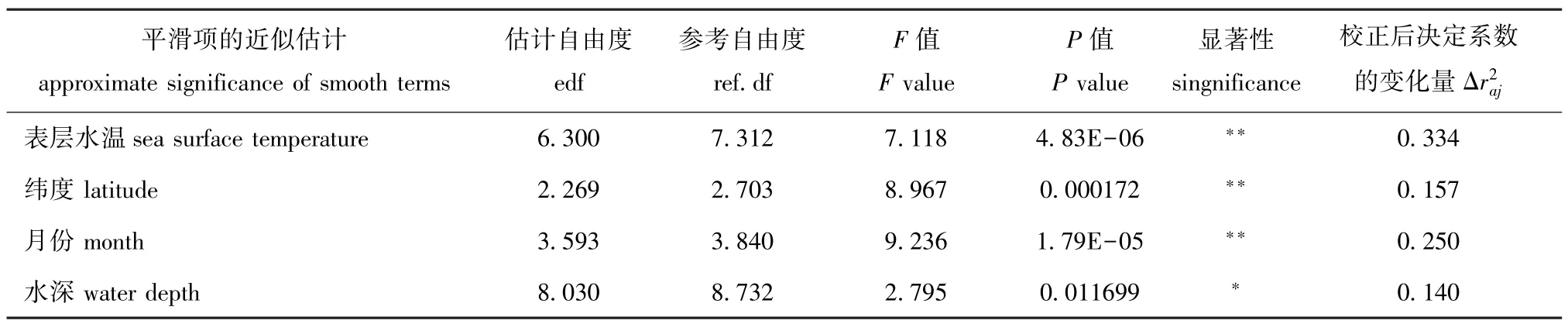
表3 GAM最优模型中各预测变量的平滑函数自由度及显著性Tab.3 Significance of smooth terms of the predictors in GAM
(2)基于数值变量的 GAM分析。将 MO、LON和LAT视为数值变量,对口虾蛄假蚤状幼体和各预测变量进行逐步运算,得到最优GAM模型的表达式为
Ind~s(SST)+s(LAT)+s(MO, k=5)+s(WD)。其中,该模型的累计偏差解释率为78.5%,为0.683,AIC为241.20。根据校正后决定系数的变化量4个预测变量的相对重要性依次为表层水温 (0.334)>月份 (0.250)>纬度 (0.157)>水深 (0.140)(表3)。
从图3可见:表层水温对幼体密度的影响明显,在15~18℃时,对幼体密度的影响随着表层水温的增加而增加,在18℃左右时影响最大,在23~27℃时,水温与幼体密度呈负相关;纬度对幼体密度的影响主要在38.50°N处,对幼体密度的影响随纬度的增加呈先减少后趋于平稳的趋势;月份对幼体密度的影响主要体现在6—8月,影响程度呈逐月减少,5月和10月与幼体密度呈负相关;水深对幼体密度的影响主要在11.5 m以浅,在11.5~20.0 m时水深对幼体密度的影响趋于平稳,在5.0~6.5 m时随着水深的增加,水深对幼体密度的负向影响逐渐减少,随后变化曲线下降后又再次上升,在10.5 m时达到最大值。
从表层水温和月份的交互作用 (图4)分析,6月,表层水温为18℃时,其对幼体密度的影响最大,线性预测值接近0.6;其次是8月,表层水温为30℃时其对幼体密度的影响较大,线性预测值为0.4~0.6。从表层水温和纬度的交互作用(图5)分析,在表层水温低于15℃、纬度为38.50°N时,对幼体密度的影响最大,线性预测值为 0.8;其次,表层水温为 30℃、纬度为38.50°N时,对幼体密度的影响较大,线性预测值接近0.7;表层水温在18℃、纬度为39.00°N~39.10°N区域时,对幼体密度的影响较大,线性预测值约为0.65。
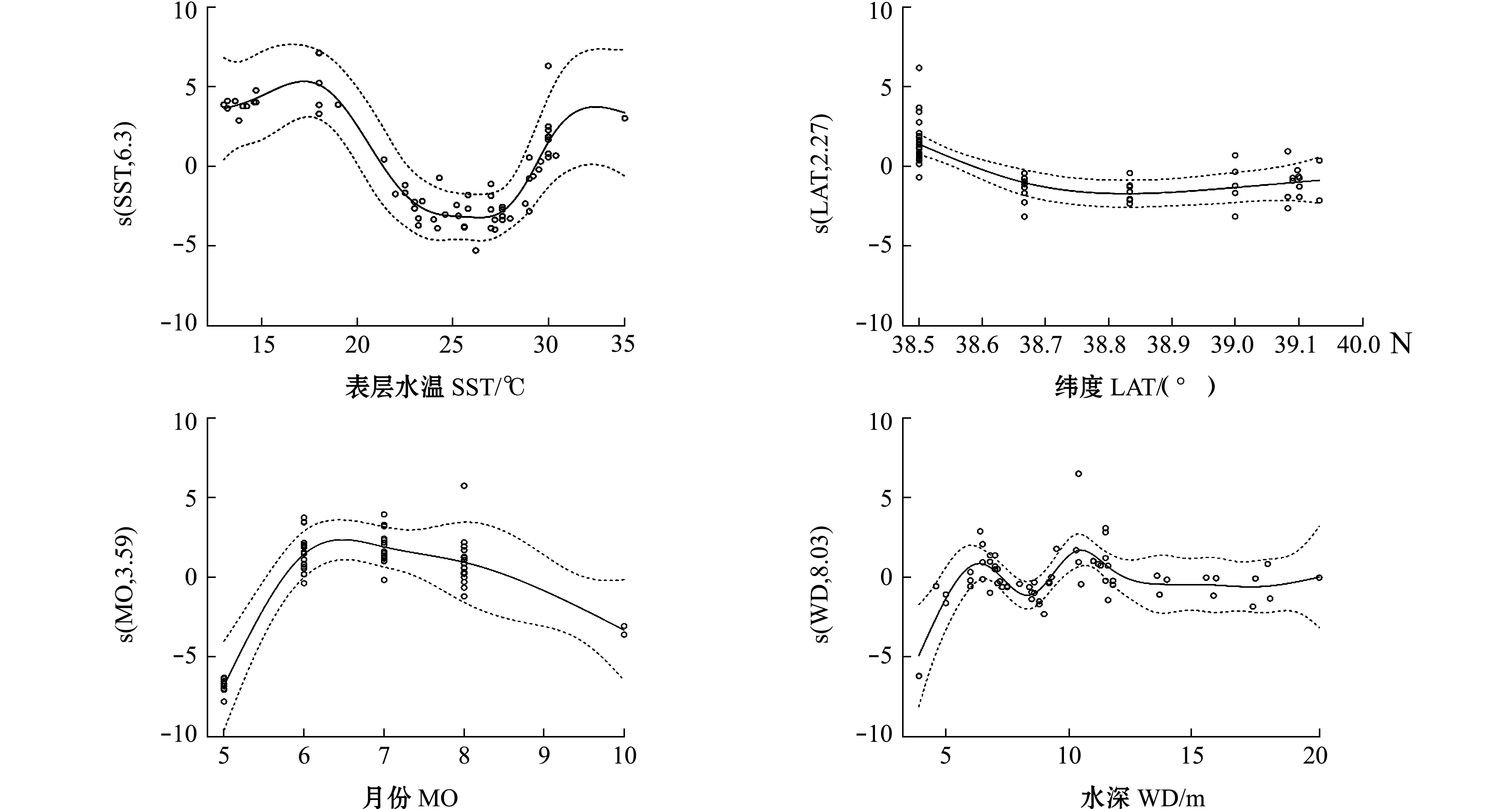
图3 口虾蛄假蚤状幼体密度与各环境因子的关系 (纬度和月份设为数值变量)Fig.3 Relationship between the popultion density of mantis shrimp Oratosquilla oratoria pseudozoea and predictor variables(latitude and month are treated as numeric variables)

图4 水温和月份的交互作用效应图Fig.4 Contour plot views of GAM model predictions of SST and month

图5 水温和纬度的交互作用效应图Fig.5 Contour plot views of GAM model predictions of SST and latitude
3 讨论
3.1 口虾蛄假蚤状幼体的分布规律和繁殖期的推测
根据本研究的调查结果,口虾蛄假蚤状幼体的出现时间为6—8月,其资源密度存在明显的年际变化。可能的原因有:繁殖季节,不同大小亲体的繁殖时间不同,以及每个海域亲体的组成比例不同,因此,口虾蛄假蚤状幼体分布规律较复杂[35];口虾蛄假蚤状幼体密度受口虾蛄产卵群体资源密度
(生物量)的影响,而口虾蛄资源量的变化可能因海区而异,如莱州湾[26]、辽东湾[25]、渤海湾[27]口虾蛄8月资源量最大,而海州湾[36]5月份口虾蛄资源量最大,并受环境、食物、敌害等诸多因素的影响。
根据本研究中口虾蛄假蚤状幼体的出现时间和口虾蛄抱卵时长,推测渤海湾口虾蛄的产卵时间为5月—8月 (或9月) (本研究中9月未开展调查)。据王波等[2]对口虾蛄生态习性的综述,口虾蛄的繁殖期为4—9月,繁殖盛期为5—7月。各地口虾蛄繁殖期略有差异:大连市皮口海域口虾蛄的繁殖盛期在5—9月[37];日本东京湾[9]口虾蛄的产卵期为4—8月;日本Hakata湾[31]口虾蛄产卵期为4—9月,高峰期为6月,幼体主要出现在8月(水温为28.0~29.7℃),其次为10月 (21.7~22.2℃),再次为9月 (24.2~24.5℃)。口虾蛄的繁殖期、繁殖盛期可能受水温的影响较大,Hakata湾[31]10月的水温为21.7~22.2℃,此时仍有大量口虾蛄幼体出现,而渤海湾在10月的水温已不足20℃,2015—2016年10月均未采到幼体。
3.2 GAM模型分析方法
月份、经度和纬度既可视为数值变量,又可视为分类变量,如唐浩等[38]将月份、李德伟等[39]将经度和纬度视为数值变量,苏巍等[40]将月份视为分类变量。本研究中利用2015—2016年的调查数据,通过上述两种处理方式分别进行GAM分析,结果表明,用两种处理方法得到最优GAM公式,选取的预测变量及各预测变量的相对重性排序均一致,依次为表层水温>月份>纬度>水深。以数值方式处理,可以进行更多的分析,如月份和水温的交互作用分析。以分类变量处理,则能更好地反映各因素水平的显著性差异。
3.3 环境影响因子对口虾蛄假蚤状幼体密度的影响
为便于描述,本研究中只讨论基于数值分析的结果。模型结果表明:表层水温是影响幼体密度的第一要素,月份变化也体现了表层水温的变化,纬度和水深则反映了幼体的空间分布;盐度、溶解氧、无机氮、活性磷酸盐、化学耗氧量和叶绿素a均对幼体密度的影响较小。
水温对幼体密度的影响明显,在15~18℃时,其对幼体密度的影响随着水温的增加而增加,在18℃左右时影响最大。据王波等[2]的研究,在水温为25℃时,口虾蛄的孵化需要近2周,水温越低孵化时间越长;从孵化到Ⅺ期幼体需要32~51 d,由此推测,6月采集到的幼体应来自5月或4月的产卵群体。即水温低于18℃时,口虾蛄可进行产卵,而刘海映等[41]推断口虾蛄产卵期为6—7月,海区的水温均高于18℃,可能是地区差异。本研究中,从水温看,18℃左右时对幼体密度影响最大;从月份看,6—8月对幼体密度的影响较大 (图3),8月的影响较7月有所下降。从水温和月份的交互作用分析 (图4),6月,水温为18℃时对幼体密度的影响最大;其次是8月,水温在30℃左右时影响较大。薛梅等[37]对大连市皮口海域口虾蛄繁殖群体的调查表明,口虾蛄的繁殖盛期为5—9月。本研究中,10月未采集到幼体,除水温因素外,9月1日禁渔期结束可能对产卵群体产生了影响。
空间分布方面,基于GAM的分析结果,纬度对幼体密度的影响较大。整体上看,纬度为38.50°N时对幼体密度的影响最大,其他几个纬度的差异不大。从水温和纬度的交互作用分析 (图5),在水温低于15℃、纬度为38.50°N时对幼体密度的影响最大;其次,水温为18℃、纬度为39.10°N时对幼体密度影响较大。纬度影响显著的原因,可能源于站点设置的变化,2015年的调查主要集中在纬度38.50°N处,2016年的调查主要集中在其他纬度。
从水深分析,6.5 m和10.5 m水深对幼体密度的影响较大。结合月份和水深,6月10 m以深的水域,随着水深的增加水深对幼体密度的影响呈负相关。这与辽东湾口虾蛄成体的分布较相似。6月辽东湾的口虾蛄生物量与水深呈负相关 (Pearson相关),8月与水深的相关性不显著[25]。本研究中,6—8月水深对幼体密度影响较大,主要集中在6~12 m的区域 (出现2个峰值区),7月水深为10~11 m的区域,以及7月水深为6 m的区域。
综上所述,从时间上分析,对口虾蛄假蚤状幼体密度影响较大时期为表层水温为18℃左右的6月,以及表层水温为30℃左右的8月,这个时间段属于渤海禁渔期,可对口虾蛄假蚤状幼体的保护起到较好的作用。
从空间上分析,水深为10.5 m和6.5 m的区域对幼体密度影响最大,而空间分布模式仍需长期调查,并考虑幼体阶段的饵料生物、敌害生物、捕捞和污染物的影响,以便更好地开发和保护口虾蛄渔业资源。
[1] 刘瑞玉.中国海洋生物名录[M].北京:科学出版社,2008:659.
[2] 王波,张锡烈,孙丕喜.口虾蛄的生物学特征及其人工苗种生产技术[J].黄渤海海洋,1998,16(2):64-73.
[3] 农业部渔业局.2011中国渔业年鉴[M].北京:中国农业出版社,2011:230.
[4] 农业部渔业局.2012中国渔业年鉴[M].北京:中国农业出版社,2012:230.
[5] 农业部渔业渔政管理局.2013中国渔业年鉴[M].北京:中国农业出版社,2013:230.
[6] 农业部渔业渔政管理局.2014中国渔业年鉴[M].北京:中国农业出版社,2014:230.
[7] 农业部渔业渔政管理局.2015中国渔业年鉴[M].北京:中国农业出版社,2015:230.
[8] Provenzano Jr A J,Manning R B.Studies on development of stomatopod crustacea II.The later larval stages of Gonodactylus oerstedii Hansen reared in the laboratory[J].Bulletin of Marine Science,1978,28(2):297-315.
[9] 大富潤,清水誠,Vergara J A M.東京湾のシャコの産卵期について[J].日本水産學會誌,1988,54(11):1929-1933.
[10] Hamano T.Mating behavior of Oratosquilla oratoria(De Haan,1844)(Crustacea:Stomatopoda)[J].J Crust Biol,1988,8(2):239-244.
[11] Hamano T,Matsuura S.Egg laying and egg mass nursing behaviour in the Japanese mantis shrimp[J].Nippon Suisan Gakkaishi,1984,50(12):1969-1973.
[12] Hamano T,Matsuura S.Food habits of the Japanese mantis shrimp in the benthic community of Hakata Bay[J].Nippon Suisan Gakkaishi,1986,52(5):787-794.
[13] Hamano T,Matsuura S.Egg size,duration of incubation,and larval development of the Japanese mantis shrimp in the laboratory[J].Nippon Suisan Gakkaishi,1987,53(1):23-39.
[14] 徐善良,王春琳,梅文骧,等.浙江北部海区口虾蛄繁殖和摄食习性的初步研究[J].浙江水产学院学报,1996,15(1):30-36.
[15] 徐海龙,张桂芬,乔秀亭,等.黄海北部口虾蛄体长及体质量关系研究[J].水产科学,2010,29(8):451-454.
[16] 林月娇,刘海映,徐海龙,等.大连近海口虾蛄形态参数关系的研究[J].大连水产学院学报,2008,23(3):215-217.
[17] 徐善良,王春琳,梅文骧,等.口虾蛄形态参数关系的研究[J].浙江水产学院学报,1996,15(1):15-20.
[18] 钱云霞,蒋霞敏,王春琳,等.黑斑口虾蛄消化酶的初步研究[J].中国水产科学,2000,7(2):100-102.
[19] 吴耀华,赵延霞.黑斑口虾蛄对水温、盐度和pH的耐受性研究[J].水产科学,2015,34(8):502-505.
[20] 徐善良,王春琳,梅文骧,等.口虾蛄Oratosquilla oratoria(De Huan)性腺特征及卵巢组织学观察[J].浙江水产学院学报,1996,15(1):21-29.
[21] 刘海映,秦玉雪,姜玉声,等.口虾蛄胚胎发育的研究[J].大连海洋大学学报,2011,26(5):437-441.
[22] 谷德贤,洪星.口虾蛄人工繁殖试验[J].北京水产,2007(6):33-36.
[23] 谷德贤,洪星,刘海映.口虾蛄的繁殖行为[J].河北渔业,2008(1):37-40.
[24] 刘海映,谷德贤,李君丰,等.口虾蛄幼体的早期形态发育特征[J].大连水产学院学报,2009,24(2):100-103.
[25] 刘修泽,郭栋,王爱勇,等.辽东湾海域口虾蛄的资源特征及变化[J].水生生物学报,2014,38(3):602-608.
[26] 吴强,陈瑞盛,黄经献,等.莱州湾口虾蛄的生物学特征与时空分布[J].水产学报,2015,39(8):1166-1177.
[27] 谷德贤,刘茂利.天津海域口虾蛄群体结构及资源量分析[J].河北渔业,2011(8):24-26.
[28] 刘海映,王桂娥,王秀利.大连海域口虾蛄资源遗传多样性的分析[J].大连水产学院学报,2009,24(4):350-353.
[29] 张代臻,丁鸽,张华彬,等.渤海湾葫芦岛海域口虾蛄mt COI基因序列多态性研究[J].江苏农业科学,2011,39(3):55-57.
[30] 邵松军,李明勇,张湘宁,等.口虾蛄乙酸乙酯提取物联合阿霉素或顺铂对人肝癌HepG2细胞的增殖抑制效应研究[J].中国医药生物技术,2013,8(5):344-348.
[31] Hamano T,Morrissy N M,Matsuura S.Ecological information on Oratosquilla oratoria(Stomatopoda,Crustacea)with an attempt to estimate the annual settlement date from growth parameters[J].The Journal of the Shimonoseki University of Fisheries,1987,36(1):9-27
[32] 国家技术监督局.GB/T 12763-2007海洋调查规范[S].北京:中国标准出版社,2007.
[33] 谷德贤,刘国山,王晓宇,等.基于GAM模型的天津海域鱼类资源和环境因子关系的初步研究[J].天津农学院学报,2017,24(1):38-43.
[34] Dong Xuhui,Bennion H,Maberly S C,et al.Nutrients exert a stronger control than climate on recent diatom communities in Esthwaite Water:evidence from monitoring and palaeolimnological records[J].Freshwater Biology,2012,57(10):2044-2056.
[35] Kodama K,Shimizu T,Yamakawa T,et al.Reproductive biology of the female Japanese mantis shrimp Oratosquilla oratoria(Stomatopoda)in relation to changes in the seasonal pattern of larval occurrence in Tokyo Bay,Japan[J].Fisheries Science,2004,70(5):734-745.
[36] 许莉莉,薛莹,焦燕,等.海州湾及邻近海域口虾蛄群体结构及资源分布特征[J].中国海洋大学学报,2017,47(4):28-36.
[37] 薛梅,闫红伟,刘海映,等.大连市皮口海域口虾蛄群体繁殖生物学特征初步研究[J].大连海洋大学学报,2016,31(3):237-245.
[38] 唐浩,许柳雄,陈新军.基于GAM模型研究时空及环境因子对中西太平洋鲣鱼渔场的影响[J].海洋环境科学,2013,32(4):518-522.
[39] 李德伟,张龙,王洋,等.基于GAM的阿根廷滑柔鱼CPUE与环境因子关系分析[J].渔业现代化,2015,42(4):56-61.
[40] 苏巍,薛莹,任一平.海州湾海域鱼类分类多样性的时空变化及其与环境因子的关系[J].中国水产科学,2013,20(3):624-634.
[41] 刘海映,谷德贤,姜玉声,等.口虾蛄繁殖周期及生殖细胞发育的研究[J].大连海洋大学学报,2013,28(3):269-272.
猜你喜欢
中国海洋大学学报(自然科学版)(2022年9期)2022-09-05
科学养鱼(2022年6期)2022-07-29
渔业科学进展(2022年1期)2022-01-14
生物学通报(2021年5期)2021-03-16
海洋通报(2020年3期)2020-10-20
齐鲁周刊(2017年29期)2017-08-08
方圆(2015年15期)2015-09-10
水生生物学报(2014年3期)2014-03-29
测绘学报(2014年2期)2014-01-11
大连海洋大学学报(2012年3期)2012-06-06
