
短日照处理对小豆生长、光合特性及产量形成的影响
2018-02-10 00:49:43董伟欣尹宝重路战远张月辰
福建农林大学学报(自然科学版) 2018年1期
董伟欣, 尹宝重, 路战远, 张月辰
(1.河北农业大学农学院,河北 保定 071001;2.河北广播电视大学,河北 石家庄 050080;3.内蒙古自治区农牧业科学院,内蒙古 呼和浩特市 010000)
我国是小豆(Vignaangularis)生产的第一大国,种植面积和总产量居世界首位,也是世界上最主要的小豆出口国,常年出口量40 000~60 000 t[1].其蛋白质含量高,富含多种维生素及丰富的钙、铁、磷元素以及人体必需的赖氨酸、色氨酸、苯丙氨酸、蛋氨酸、苏氨酸、异亮氨酸、亮氨酸和缬氨酸[2].但小豆落花落荚严重、产量低且不稳定一直是生产中的难题.光周期在植物生活周期中起着重要作用,植物的生长发育及光形态建成等都受日照长度的调节[3-4].小豆属于典型的短日照作物,在出苗至开花时具有很强的感光性,但不同品种敏感程度不同[5],导致小豆品种具有不同的生育进程.因此,利用光周期来调控不同小豆品种的生长、光合特性以及产量形成对今后小豆的栽培技术改良以及适期育种具有重要的现实意义.
光周期处理使大豆的全生育期发生改变且形态指标也随之发生改变,大豆的株高、分枝数、主茎节数减少,茎秆重在短日照处理下分别降低[6],另有研究表明短日照处理显著降低株高、节数、单株干重且降低干物质积累量[7].日照长度不仅影响作物的生长,还影响作物的光合特性.短日照下‘Royal touch’的光合速率、气孔导度、以及细胞间CO2浓度均高于长日照下的[8].短日照处理提高了小麦旗叶的Fo值,降低了ΦPSⅡ和qP,表明ΦPSⅡ光化学反应中心活性已受到影响[9].加拿大紫荆的叶绿素含量在短日照下增加,叶绿素a/b下降,Fv/Fm也下降[10].日照长度对产量和产量构成因素有明显的影响,Chaturvedi et al和Thangarag et al[11-12]的研究表明,短日照后水稻有效分蘖数和有效穗数降低,籽粒充实度和千粒重呈下降趋势,从而导致产量降低.尹淑丽等[13]的研究也表明短日照处理对于不同熟性红小豆品种的产量均为负效应,日照长度越短、处理时间越长,产量降低的幅度越大.
小豆作为典型的短日照作物,以往只针对某个单一品种进行光周期研究,通过对光照敏感程度不同的早、中、晚熟小豆品种进行短日照处理来研究其生长、叶片光合特性和产量形成以及不同品种之间的差异却少见报道.本研究采用对光照敏感程度不同的早熟品种白红2号、中熟品种冀红9218和晚熟品种唐山红小豆作为供试材料,研究其生长、叶片光合特性以及产量形成对不同日照长度的反应,为当地及相似的生态区域适宜品种的选择、适期播种、加速生育进程、光周期调控等方面的栽培技术改良提供理论依据.
1 材料与方法
1.1 试验材料
试验于2013—2014年在河北农业大学教学实验基地进行(河北保定,经度和纬度分别为115°28′E和38°52′N),以2014年试验结果为准,选用早熟品种‘白红2号’、中熟品种‘冀红9218’和晚熟品种‘唐山红小豆’作为试验材料,3个品种均由河北省农林科学院粮油作物研究所提供.
1.2 试验设计
本试验设置的日照长度为12 h光/12 h暗,在出苗1周后设置SD-12h-1WK(12 h光/12 h暗处理1周)、SD-12h-2WKS(12 h光/12 h暗处理2周)和SD-12h-3WKS(12 h光/12 h暗处理3周)3种处理,以自然光照(简称:NPD,试验季节自然光大约每天的光照时间为14.5 h)作为对照.
2年日期均选择在6月24日播种,处理方法是每天19:00用不透光布遮光,次日早晨7:00解除短日照.田间种植采用裂区设计,品种为主区,光周期处理为副区,处理时间为副副区,采用穴播方法,每穴2 粒,小区面积5 m×1 m,每个小区种植2行,株距15 cm,行距40 cm,3次重复,不同生育时期短日照处理完毕后在自然条件下生长至成熟,生育期间的管理同大田.
1.3 取样方法
在苗期、分枝期、开花期、结荚期和鼓粒期5个生育时期,于上午8:00采取自上数第2个完全展开的三出复叶测定生理指标.
1.4 测定指标与方法
1.4.1 光合参数测定 采用美国LI-COR公司LI-6400光合测定系统,选取生长一致的三出复叶的中间小叶,每个小区选取3叶,3次重复,共取9叶,在上午9:00—11:00或下午14:00—16:00光照达到1 400 lx时测定净光合速率(Pn)、细胞间CO2浓度(Ci)、蒸腾速率(Tr)和气孔导度(Gs).
1.4.2 荧光参数测定 叶绿素荧光参数测定采用英国Hansatech公司FMS-2调制叶绿素荧光仪测定,在晴朗无风的天气,先测定光下稳态荧光(Fs)、最大荧光(Fm′),而后暗适应20 min获取初始荧光(Fo)、最大荧光(Fm)和PSⅡ原初光能转化效率Fv/Fm,经计算得到PSⅡ实际光化学量子产量(ΦPSⅡ).计算公式为:(1) Fv/Fm=(Fm-Fo)/Fm和(2)ΦPSⅡ=(F′m-Fs)/F′m.
1.5 产量指标测定
不同短日照处理后,每个小区选取具有代表性植株9株,用直尺测量株高、叶柄长度和节间长度,用3 V锂电数显卡尺测定植株直径,再折算成茎粗,数出分枝数和主茎节数;干物重在烘箱内105 ℃杀青后在60 ℃下烘干48 h;在成熟期测定单株荚数、单株荚重、单株粒重、单荚粒数、单荚粒重及小区产量.
1.6 数据处理
每个处理重复3次,所有数据采用Microsoft Excel 2007整理和作图,采用DPS v7.0和SPSS 19.0软件的Duncan′s方法做差异显著性(P<0.05)分析.
2 结果与分析
2.1 短日照处理对小豆植株形态指标的影响
由表1可见,白红2号的株高在SD-12h-3WKS处理下较对照显著降低34.3%,冀红9218和唐山红小豆株高在SD-12h-2WKS和SD-12h-3WKS处理下显著降低.白红2号和冀红9218的茎粗和主茎节数在SD-12h-2WKS和SD-12h-3WKS处理下显著降低,在SD-12h-1WK处理下降低不明显,而唐山红小豆降低都不显著.短日照对3个小豆品种的主茎分枝数和节间长度无明显影响.唐山红小豆的叶片数在SD-12h-2WKS和SD-12h-3WKS处理下显著降低34.5%和54.9%,而白红2号和冀红9218降低不明显.3个品种的叶柄长度在SD-12h-2WKS和SD-12h-3WKS处理下显著降低,白红2号分别降低13.9%和30.7%,冀红9218分别降低16.6%和30.7%,唐山红小豆分别降低12.9%和35.1%,而在SD-12h-1WK处理下降低不明显.以上说明,短日照处理使3个品种的植株形态指标呈现下降趋势且唐山红小豆>冀红9218>白红2号,且处理时间越长,降低就越多.

表1 短日照处理对不同熟性小豆品种植株形态指标的影响1)Table 1 Effect of short-day photoperiod treatment on plant morphological index of adzuki bean with different maturities
1)均值±SE,同列中小写字母不同者表示经新复极差法检验在0.05水平上的统计显著性.
2.2 短日照处理对小豆干物质积累的影响
从表2可以看出,白红2号和唐山红小豆的地上部干重在SD-12h-3WKS处理下显著降低35.1%和31.6%,冀红9218在SD-12h-2WKS和SD-12h-3WKS处理下显著降低24.1%和43.3%.短日照对3个品种的茎秆干重、叶片干重和单株荚皮重无明显影响.白红2号的叶柄干重在SD-12h-2WKS和SD-12h-3WKS处理下显著降低36.4%和37.7%,冀红9218和唐山红小豆在SD-12h-3WKS处理下分别降低43.6%和53.9%,其它2个处理降低不明显.白红2号的单株荚重和单株粒重在SD-12h-3WKS处理下分别降低46.9%和47.4%,而冀红9218在3个处理下与对照相比都显著降低,而唐山红小豆降低不明显.以上可见,短日照使3个品种的干物质积累呈现降低趋势且短日照处理时间越长,干物质积累就越少,且表现为短日照处理时间的剂量效应,晚熟品种对日照较敏感,下降也较多.
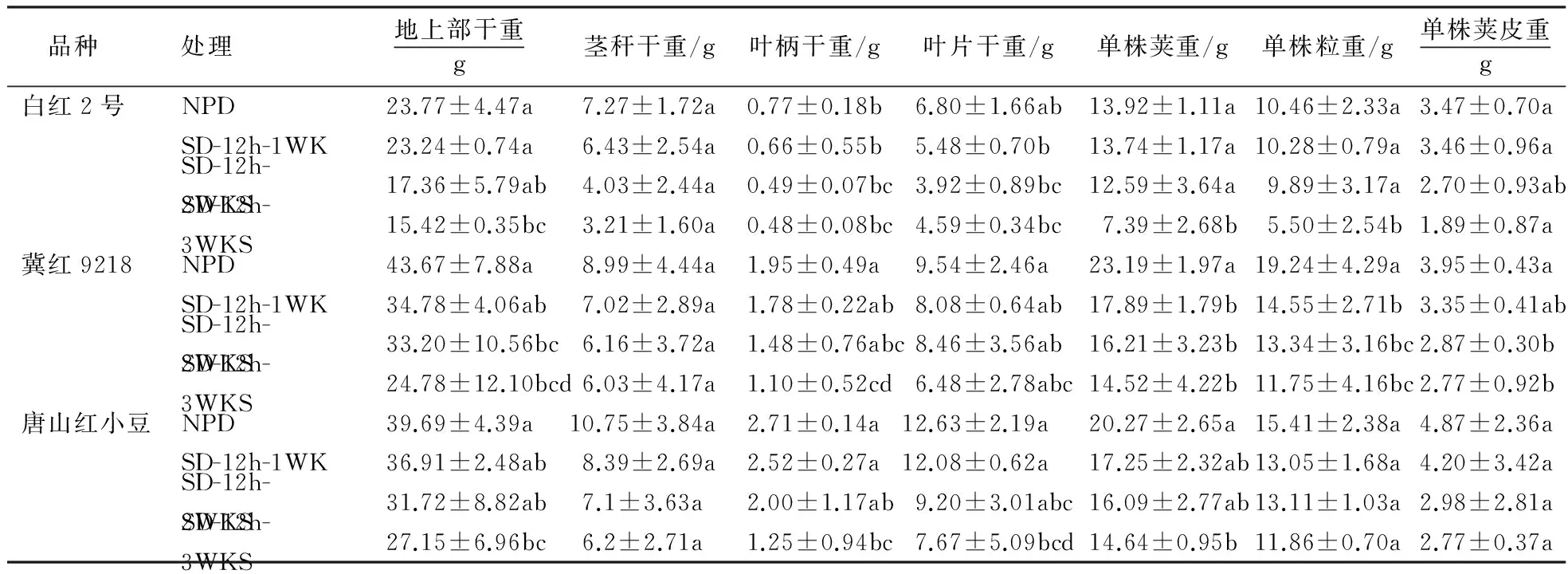
表2 短日照处理对不同熟性小豆品种干物质积累的影响1)Table 2 Effects of short-day photoperiod on dry weight accumulation of adzuki bean with different maturity
1)均值±SE,同列中小写字母不同者表示经新复极差法检验在0.05水平上的统计显著性.
2.3 短日照处理对小豆净光合速率的影响
由图1可知,白红2号的净光合速率在苗期、分枝期和鼓粒期的3个处理较对照都降低且处理时间越长,降低越多,在SD-12h-3WKS处理下显著降低23.5%、14.8%和18.9%,在结荚期也降低,但随着诱导时间的延长,净光合速率随之升高,差异都不明显,在开花期的3个处理较对照分别显著升高12.7%、13.2%和15.3%(图1A).冀红9218在苗期、开花期和鼓粒期净光合速率都升高,但差异不显著,而在分枝期和结荚期都降低且处理时间越长,降低就越多(图1B).唐山红小豆分枝期的3个处理较对照升高,从开花期开始,短日照处理时间越长,净光合速率较对照降低越多,在苗期3个处理有所降低,差异不明显(图1C).从以上可以看出,开花期是小豆的一个转折点,此时期3个品种的净光合速率都升高.

A.白红2号;B.冀红9218;C.唐山红小豆.图1 短日照处理对不同熟性小豆品种净光合速率的影响Fig.1 Effect of short-day photoperiod on net photosynthetic rate of adzuki bean with different maturity
2.4短日照处理对小豆细胞间二氧化碳浓度的影响
由图2可知,白红2号的细胞间二氧化碳浓度在开花期略有升高,在其它几个生育时期都降低,差异不明显(图2A).而冀红9218在分枝期较对照降低,差异不显著,在其它几个生育时期都升高,在苗期和开花期处理时间越长,净光合速率越高,而在结荚期和鼓粒期却相反(图2B).唐山红小豆在苗期至鼓粒期都降低,都呈现出短日照处理时间越长,净光合速率降低就越多的趋势,但差异不显著(图2C).以上表明,到了开花期和鼓粒期,小豆需要积累更多的营养物质来供自身需求,因此,这两个时期细胞内积累了较多的二氧化碳作为光合作用的原料.
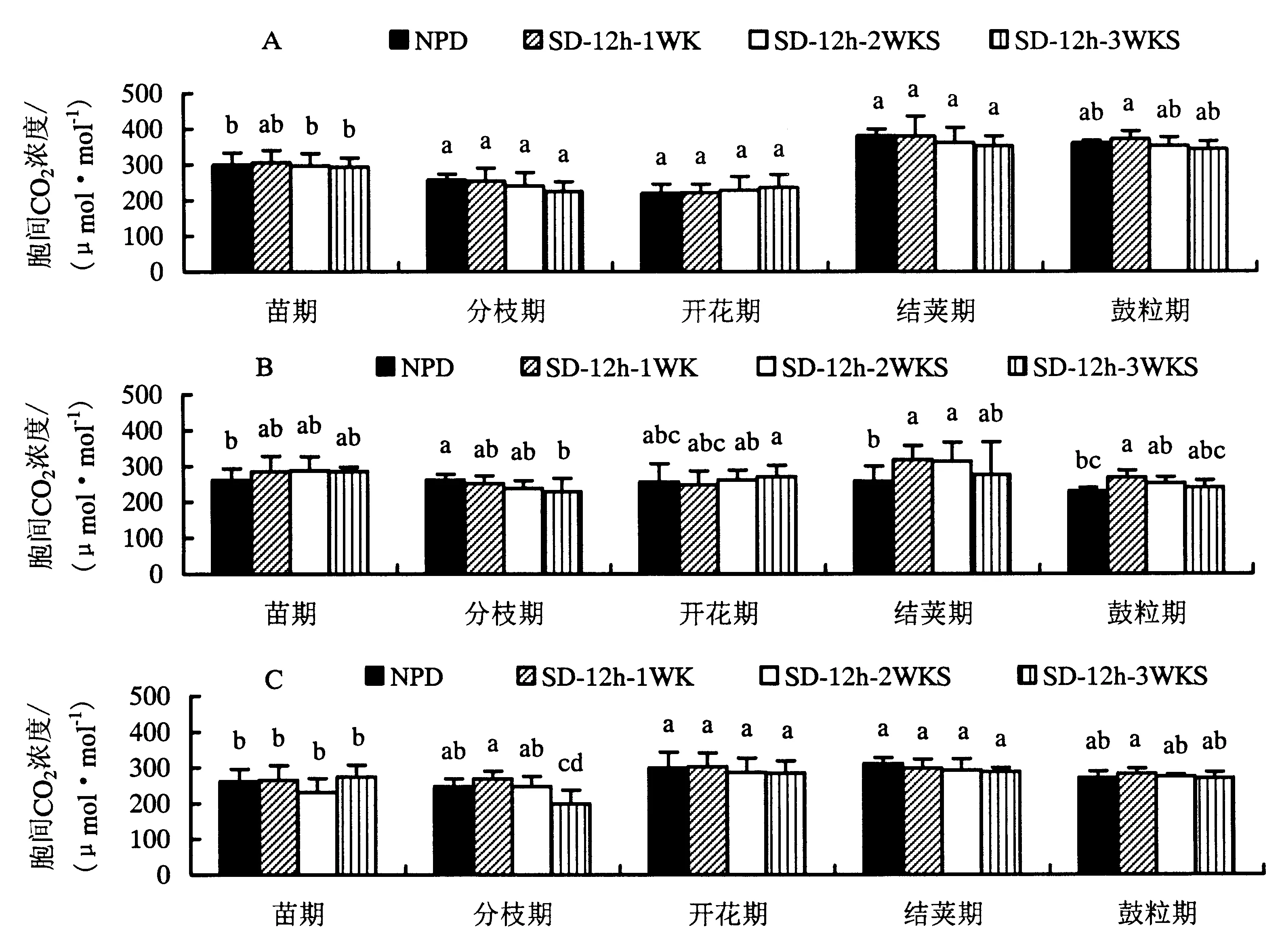
A.白红2号;B.冀红9218;C.唐山红小豆.图2 短日照处理对不同熟性小豆品种细胞间二氧化碳浓度的影响Fig.2 Effect of short-day photoperiod on intercellular CO2 concentration of adzuki bean with different maturity
2.5 短日照处理对小豆气孔导度的影响
由图3可知,白红2号的气孔导度在苗期、分枝期和鼓粒期的3个处理都较对照降低,在开花期和结荚期都较对照升高,但差异不明显(图3A).冀红9218在分枝期和开花期稍有降低,差异不明显,在苗期、结荚期和鼓粒期升高且短日照处理时间越长,升高越少,差异都未达到显著水平 (图3B).唐山红小豆在苗期和分枝期降低,从开花至鼓粒期升高且短日照处理时间越长升高越少(图3C).从以上可以看出,3个小豆品种的气孔导度从开花期开始升高,而细胞间二氧化碳浓度也升高,说明气孔导度的升高有利于细胞间二氧化碳的积累,从而促进净光合能力的增强.
2.6 短日照处理对小豆蒸腾速率的影响
从图4可以看出,白红2号的蒸腾速率在苗期和分枝期稍有下降,差异都未达到显著水平,开花期在SD-12h-2WKS和SD-12h-3WKS处理下显著升高11.4%和12.5%,结荚期都升高,但差异不显著,在鼓粒期SD-12h-3WKS处理显著升高29.1%,其余2个处理升高不显著(图4A).冀红9218只有在分枝期稍有降低,差异不明显,在其它生育时期都有一定程度的升高,但差异都不显著(图4B).唐山红小豆的3个处理在苗期和分枝期稍有降低,开花至鼓粒期都升高且短日照处理时间越长,升高越少,开花期在SD-12h-1WK和SD-12h-2WKS处理下显著升高21.2%和16.0%,结荚期只有SD-12h-1WK显著升高23.5%,在鼓粒期升高都不显著(图4C).可见,短日照处理对3个品种蒸腾速率的影响与气孔导度类似,苗期和分枝期的3个处理都低于对照,开花至鼓粒期都高于对照,因品种不同而稍有差异.

A.白红2号;B.冀红9218;C.唐山红小豆.图3 短日照处理对不同熟性小豆品种气孔导度的影响Fig.3 Effect of short-day photoperiod on stomata conductance of adzuki bean with different maturity

A.白红2号;B.冀红9218;C.唐山红小豆.图4 短日照处理对不同熟性小豆品种蒸腾速率的影响Fig.4 Effect of short-day photoperiod on transpiration efficiency of adzuki bean with different maturity
2.7 短日照处理对小豆叶片最大荧光Fm变化的影响
由表3可见,白红2号的Fm值苗期在SD-12h-1WK处理下稍有降低,SD-12h-2WKS和SD-12h-3WKS处理显著升高17.9%和20.0%,分枝期SD-12h-1WK处理降低,其余2个处理升高,差异都不显著,开花期SD-12h-3WKS显著升高17.3%,而对结荚期和鼓粒期的影响不大.冀红9218在分枝期的各个处理较对照降低,在其它生育时期都升高,差异都不显著.唐山红小豆在分枝期和开花期稍有降低,在其它生育时期都升高,苗期升高不明显,结荚期和鼓粒期的升高幅度较对照都达到显著水平.说明短日照处理下小豆叶片的 PSⅡ反应中心增强,光合能力也增强,但在某些时期会遭到一定程度的破坏而导致光合能力下降,但不同品种表现有所差异.
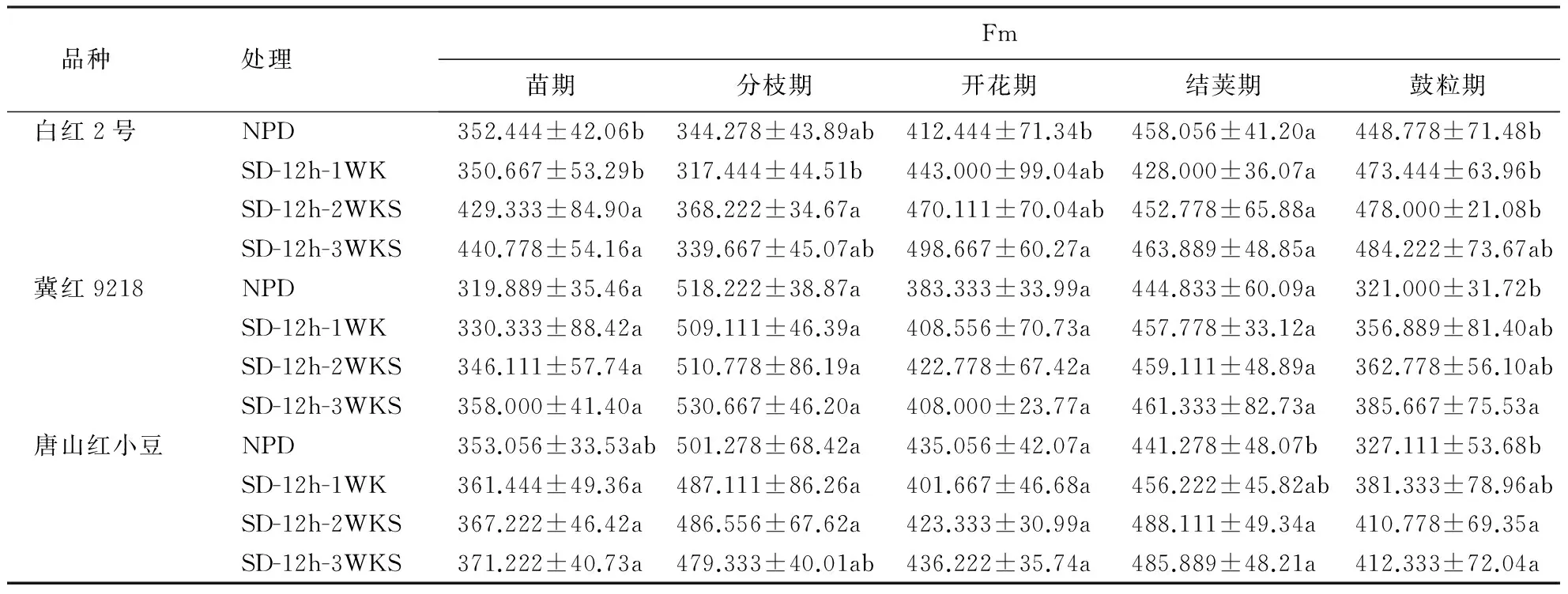
表3 短日照处理对不同熟性小豆品种最大荧光Fm变化的影响1)Table 3 Effect of short-day photoperiod on Fm of adzuki bean with different maturity
1)均值±SE,同列中小写字母不同者表示经新复极差法检验在0.05水平上的统计显著性.
2.8 短日照处理对小豆PSⅡ原初光能转化效率(Fv/Fm)的影响
由表4可见,白红2号在苗期、开花期和结荚期的3个处理较对照都有不同程度的升高,但差异都未达到显著水平,在分枝期只有SD-12h-3WKS处理显著升高3.5%,鼓粒期 SD-12h-WK和SD-2h-WKS处理显著降低7.2%和5.6%,SD-2h-WKS降低不明显.冀红9218在5个生育时期都有一定程度的升高,随着短日照处理时间的延长,Fv/Fm值不断增大.唐山红小豆在苗期和分枝期的Fv/Fm值较对照稍有降低,开花期有所升高,差异不显著,在结荚期显著升高5.0%、7.4%和7.5%,鼓粒期显著升高3.9%、5.7%和6.3%,且短日照处理时间越长,Fv/Fm值升高越多.表明短日照处理后小豆的PSⅡ潜在活性中心增强,光合作用原初反应过程增强,从而加速电子传递.
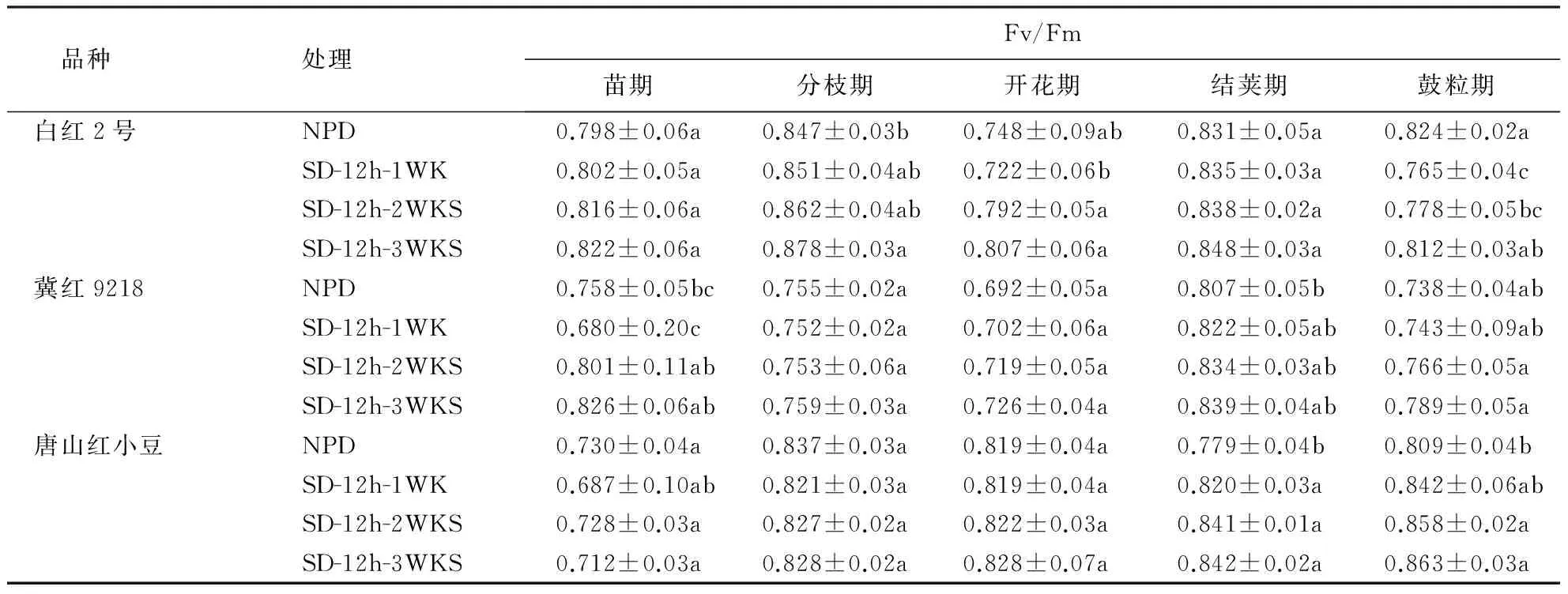
表4 短日照处理对不同熟性小豆品种PSⅡ原初光能转化效率Fv/Fm的影响1)Table 4 Effect of short-day photoperiod on Fv/Fm of adzuki bean with different maturity
1)均值±SE,同列中小写字母不同者表示经新复极差法检验在0.05水平上的统计显著性.
2.9 短日照处理对小豆PSⅡ实际光化学量子产量(ΦPSⅡ)的影响
由表5可知,短日照处理后,小豆叶片的PSⅡ实际光化学量子产量ΦPSⅡ呈现升高趋势,但3个品种升高的幅度有所不同且在分枝期和结荚期都分别出现2个高峰期.白红2号从苗期至结荚期的3个处理较对照都有一定程度的提高,差异不显著,鼓粒期分别显著升高32.6%、38.8%和50.8%,且随着短日照处理时间的延长,ΦPSⅡ也随之升高.冀红9218的ΦPSⅡ值在5个生育时期都有一定程度的升高且短日照处理时间越长,ΦPSⅡ值越大,但与对照相比差异都未达到显著水平.唐山红小豆分枝期的SD-12h-2WKS和SD-12h-3WKS处理较对照显著升高20.7%和22.7%,鼓粒期SD-12h-3WKS处理显著升高28.3%,其余2个处理升高不明显.

表5 短日照处理对不同熟性小豆品种PSⅡ实际光化学量子产量ΦPSⅡ的影响1)Table 5 Effect of short-day photoperiod on ΦPSⅡ of adzuki bean with different maturity
1)均值±SE,同列中小写字母不同者表示经新复极差法检验在0.05水平上的统计显著性.
2.10 短日照处理对不同熟性小豆品种产量构成因素和产量的影响
由表6可以看出,白红2号的单株荚数、单株荚重、单株粒重、单荚粒数和单荚粒重在SD-12h-3WKS处理下与对照相比分别显著降低36.9%、46.9%、47.4%、25.9%和23.5%,小区产量在SD-12h-2WKS和SD-12h-3WKS处理下显著降低48.0%和68.0%,在SD-12h-1WK处理下降低不明显.冀红9218的单株荚数、单株荚重和单株粒重都显著降低,而单荚粒数、单荚粒重和小区产量在SD-12h-2WKS和SD-12h-3WKS处理下显著降低,在SD-12h-1WK处理下降低不明显.唐山红小豆的单株荚数、单株荚重和单株粒重较对照无明显差异,单荚粒数和小区产量在SD-12h-3WKS处理下显著降低15.1%和46.5%,单荚粒重在SD-12h-2WKS和SD-12h-3WKS处理下显著降低17.1%和29.3%,在SD-12h-1WK处理下无显著差异.可见,短日照处理对于不同熟性小豆品种的产量均为负效应,即日照长度越短、处理时间越长,产量构成因素及小区产量降低的幅度就越大,下降幅度的大小因不同的品种而异.

表6 短日照处理对不同熟性小豆品种产量构成因素和产量的影响1)Table 6 Effect of short-day photoperiod on yield components and yields of adzuki bean with different maturity
1)均值±SE,同列中小写字母不同者表示经新复极差法检验在0.05水平上的统计显著性.
3 讨论
日照长度影响作物的生长,本研究发现短日照降低小豆植株的生长及干物质积累且日照时间越短,处理时间越长,降低就越多,影响程度为晚熟品种唐山红小豆>中熟品种冀红9218>早熟品种白红2号,其中短日照对成熟时期主茎分枝数和节间长度的影响较小,表明短日照处理对不同形态学指标的影响略有不同,品种之间也有差异.前人研究表明,短日照处理使大豆单株出生叶数、株高、分枝数、主茎节数减少,茎秆重降低[14].吉林3号、晋豆2号大豆品种在始荚后进行短日照处理可使节数、分枝数与上部荚、粒数减少,并加快灌浆鼓粒与成熟,短日照处理后因荚、粒减少而产量降低[15],这与本研究结果相一致.但也有研究发现,番茄品种百利和毛粉802在短日照处理下的植株形态增加,其中以百利表现最明显,百利的茎长为171.88 cm,节数34.6节,叶数22.6片,花序量4.6个,毛粉802的果实横径6.715 cm,纵径6.374 cm,单果重167.25 g,明显高于对照[16].中棉所49号地上部的总干物质和养分积累量在短日照处理下较对照差异较小[17].这与本研究结果不同,分析原因可能与作物品种不同有关,也可能与短日照处理时间和长短有关,还需通过不同的作物品种进行深入研究.
短日照也是影响作物光合作用的重要因子之一,如刘国顺等[18]研究发现烟草在短日照条件下的净光合速率(Pn)降低,同时ΦPSⅡ和Fv/Fm迅速上升,杨兴洪等[19]的研究发现棉花在短日照条件下净光合速率明显升高,李冬梅[20]研究表明短日照处理使桃叶片的净光合速率、气孔导度降低,胞间CO2浓度升高.另有研究发现玉米在自然光转入短日照条件下时,其净光合速率升高,同时ΦPSⅡ、Fv/Fm和qP也迅速上升[21].本研究发现3个品种的净光合速率、细胞间二氧化碳浓度、气孔导度和蒸腾速率在开花或鼓粒期呈现高峰,这与前人研究总体趋势相一致,说明在这2个生育时期需要积累较多的营养物质,营养物质转移至果荚中之后净光合速率又开始下降,白红2号在分枝期和开花期,冀红9218在苗期、分枝期和鼓粒期,唐山红小豆在鼓粒期的光合特性表现出一致的规律性,说明在不同的品种在不同的生育时期受气孔因素的影响,净光合速率升高或降低,气孔导度、细胞间二氧化碳浓度和蒸腾速率也升高或降低,原因是气孔导度升高或降低使细胞间二氧化碳供应增加或减少所致,气孔导度升高或降低而引起蒸腾速率升高或降低,以上可以看出,冀红9218受气孔因素的影响较大,白红2号次之,晚熟品种最小,3个品种的其它生育时期均表现不一致的规律,推测可能是受非气孔因素的影响,如其它某些化学活性物质的影响,其内在机理需要深入探究.小豆叶片的最大荧光(Fm)、PSⅡ的原初光能转化效率(Fv/Fm)、PSⅡ实际光化学量子产量(ΦPSⅡ)呈现升高趋势且短日照时间越长,升高越多,推测原因各个生育时期在短日照下所积累的营养物质不能满足其自身生长发育的需求,所以叶片就要提高PSⅡ反应中心开放比例和PSⅡ光化学效率来获取更多的光能来供应植株的生长发育.
本研究发现3个小豆品种的产量构成因素和产量在短日照处理下呈降低趋势,且短日照处理时间越长,降低幅度就越大,前人对棉花[17]和水稻[22]研究也发现短日照处理后产量及产量构成因素降低,但王久兴研究发现,短日照下番茄可通过提高单果重、单株结果数来显著提高单株产量,增幅可达63.7%[16].这与本研究结果相反,因为本研究是进行12 h短日照处理且处理时间长短不一致,前人在其它作物上分为不同的阶段进行短日照处理,不同的时期对产量的影响不同,而且品种之间也有差异,本课题组在之前的研究中也在开花之前分为出苗—分枝、分枝—开花以及出苗至开花期进行短日照处理,其中出苗至开花期对小豆产量的影响最大,出苗—分枝期次之,分枝-开花期的影响最小,而且晚熟品种唐山红小豆较早熟品种白红2号受短日照的影响较大,至于开花之后短日照是否还存在光周期反应以及对产量的影响,本课题组还在进行中.
综上所述,短日照处理降低小豆植株的生长且抑制植株各部分的干物质积累,但可以提高小豆叶片的光合特性并加速电子传递过程,随着短日照处理时间的延长,产量及产量构成因素逐渐下降,通过合理的短日照处理可以调控植株的生长及叶片的生理变化来改变小豆的生育进程且对产量无明显影响,这为当地及相似生态区域的栽培技术改良及加速育种进程提供现实参考.
[1] 于振文.作物栽培学各论(北方本)[M].北京:中国农业出版社,2003:292-293.
[2] 中国医学科学院卫生研究所.食物成分表[M].北京:人民卫生出版社,1981.
[3] KODYM A, ZAPATA-ARIAS F G. Natural light as an alternative light source for theinvitroculture of banana (Musaacuminatacv. ‘Grande Naine’)[J]. Plant Cell, 1998,55(2):141-145.
[4] BAKER C M, BURNS J A, WETZSTEIN H Y. Influence of photoperiod and medium formulation on pea nut somatic embryogenesis[J]. Plant Cell Reports, 1994,13(3-4):159-163.
[5] 蒋陵秋,金文林.小豆生长发育规律的研究Ⅷ.小豆短日处理的效应[J].北京农学院学报,1991,6(1):22-27.
[6] SHANMUGASUNDARAM S. Variation in the photoperiodic response on several characters in soybean[J]. Euphytica, 1979,28(2):495-507.
[7] 韩天富,王金陵,范彬彬,等.开花后光照长度对大豆农艺性状的影响[J].应用生态学报,1996,7(2):169-173.
[8] 裴海霞,石雷,张金政,等.不同光周期对德国鸢尾‘Royal touch’的花芽分化和光合作用的影响[J].热带亚热带植物学报,2006,14(6):477-481.
[9] 牟会荣,姜东,戴廷波,等.遮阴对小麦旗叶光合及叶绿素荧光特性的影响[J].中国农业科学,2008,41(2):599-606.
[10] 张依,卓丽环,汪成忠.遮光对紫叶加拿大紫荆光合速率及叶绿素荧光参数的影响[J].河北农业大学学报,2009,32(6):52-56.
[11] CHATURVEDI G S, INGRAM K T. Growth and yield of lowland rice in response to shade and drainage[J]. Philippine Journal of Crop Science, 1989,14(2):61-67.
[12] THANGARAJ M, SIVASUBRAMANIAN V. EfFects of low light intensity on growth and productivity of irrigated rice (OryzasativaL.)[J]. Madras Agricultural Journal, 1990,77(2):220-224.
[13] 尹淑丽,张月辰,陶佩君,等.苗期日照长度对红小豆生育特性和产量的影响[J].中国农业科学,2008,41(8):2 286-2 293.
[14] 张桂茹.光周期对大豆品种生育进程及农艺性状的影响[J].中国油料,1997,19(4):37-41.
[15] 梁慧贤,刘汉中,张起刚,等.光照时数对结荚后大豆生育的影响及其原因的探讨[J].北京农业大学学报,1984,10(1):61-71.
[16] 王久兴,阎立英,毛秀杰.遮光对越夏番茄生长及产量的影响[J].江苏农业科学,2008(1):140-143.
[17] 侍伟红.红花铃期遮光对棉花产量、品质及光合生理特性的影响[D].乌鲁木齐:新疆农业大学,2013:11-14.
[18] 刘国顺,赵献章,韦凤杰,等.旺长期遮光及光照转换对不同烟草品种光合效率的影响[J].中国农业科学,2007,40(10):2 368-2 375.
[19] 杨兴洪,邹琦,王玮.遮荫棉花转入强光后光合作用的光抑制及其恢复[J].植物学报,2001,43(12):1 255-1 259.
[20] 李冬梅.光周期诱导设施桃树休眠的生理生化变化及蛋白质组学初探[D].泰安:山东农业大学,2012.
[21] 李潮海,栾丽敏,王群,等.苗期遮光及光照转换对不同玉米杂交种光合效率的影响[J].作物学报,2005,31(6):381-385.
[22] 姜楠.遮光对北方粳稻产量和品质的形成及其生理机制的研究[D].沈阳:沈阳农业大学,2013.
猜你喜欢
中国农学通报(2022年29期)2022-11-25 07:28:12
数学小灵通·3-4年级(2021年3期)2021-04-13 01:04:08
农民致富之友(2020年31期)2020-11-18 08:49:34
农村百事通(2019年24期)2019-12-31 06:11:38
家庭百事通(2017年9期)2017-09-11 15:44:09
Acta Mathematica Scientia(English Series)(2016年5期)2016-11-24 11:59:31
甘肃林业科技(2016年2期)2016-11-16 09:15:39
莫愁·家教与成才(2016年7期)2016-07-13 23:23:19
农村百事通(2014年14期)2015-02-09 10:36:18
中国火炬(2012年11期)2012-07-24 14:20:50
