
黄土高原北部生长季土壤氮素矿化对植被和地形的响应*
2018-02-08 01:56付晓莉李学章贾小旭邵明安魏孝荣
中国生态农业学报(中英文) 2018年2期
刘 娇, 付晓莉, 李学章, 贾小旭, 邵明安, 魏孝荣**
黄土高原北部生长季土壤氮素矿化对植被和地形的响应*
刘 娇1,2, 付晓莉3, 李学章4, 贾小旭3, 邵明安1,2, 魏孝荣1,2**
(1. 西北农林科技大学资源环境学院 杨凌 712100; 2. 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室 杨凌 712100; 3. 中国科学院地理科学与资源研究所 北京 100101; 4. 中国科学院亚热带农业生态研究所 长沙 410125)
氮素矿化是陆地生态系统氮循环的重要过程, 对氮素有效性有着重要影响。本文在黄土高原北部六道沟小流域选取退耕年限相近的油松和柠条坡地, 用原位培养法测定生长季节(4—10月)不同坡位冠层下和冠层外0~10 cm和10~20 cm土层土壤氮素矿化速率, 以确定该区氮素矿化的季节动态特征和主要影响因素。结果表明, 研究区生长季土壤矿质氮以铵态氮为主, 其含量在0~10 cm和10~20 cm土层分别占矿质氮总量的61%和70%, 并随生长季的推移而升高。油松林上坡位和中坡位土壤铵态氮显著高于下坡位土壤, 柠条林不同坡位铵态氮差异不显著。土壤硝态氮和矿质氮不受坡位的影响, 但与林型和采样位置有关, 冠层下硝态氮在油松林与冠层外相近, 在柠条林则高于冠层外。生长季土壤氮素矿化在0~10 cm土层由硝化作用引起, 在10~20 cm土层则由硝化和铵化作用共同引起。铵化速率在生长季初期较高, 中期较低, 并受坡位、林型和采样位置的影响。土壤硝化和矿化速率在油松林不受采样位置影响, 但是在柠条林则以冠层下较高。硝化和矿化速率在冠层下以下坡位土壤最高, 在冠层外则以下坡位土壤最低。柠条林促进了冠层下土壤氮素的硝化和矿化过程, 有利于矿质氮的积累; 油松林对矿质氮和氮素矿化的影响不受采样位置影响。
植被; 坡位; 矿化作用; 矿质氮; 采样位置
土壤氮素矿化是由微生物参与的复杂土壤生物化学过程, 直接决定着氮素的有效性, 是氮素生物地球化学循环的重要环节[1-3], 并受多种因素的影响[4-5]。巨晓棠等[6]和刘杏认等[7]的研究结果表明, 土壤水分的变幅与土壤的净氮矿化率呈正相关关系; 王常慧等[8]发现, 氮素矿化量在-10~15 ℃对温度响应微弱, 在15~35 ℃随温度升高矿化量显著增加。除土壤温度和水分, 氮素矿化还受土壤微生物活性、植被类型及植物根系等的影响。如Paul等[9]认为, 由于微生物活性在土层中呈下降趋势, 氮素矿化势也随土壤深度的加深而迅速下降; 傅声雷等[10]在鼎湖山的研究表明, 季风常绿阔叶林下土壤氮的矿化高于其他植被类型; Mueller等[11]的结果表明, 土壤氮素矿化速率与根系生物量极显著正相关。因此, 这些因素直接决定着土壤氮素矿化的季节动态特征及其对土地利用等人类活动的响应, 并影响着脆弱生态系统植被恢复和环境治理的效果。
黄土高原北部不但是风蚀和水蚀交错区, 同时是农牧业交错区, 生态脆弱, 水土流失严重[12]。该区坡地面积占土地面积的70%以上[13], 是土地退化的主要地理单元。20世纪70年代以来, 该区采取的飞播造林、退耕还林还草等措施, 在防治水土流失和土地退化方面取得了显著效果[14-15], 也使坡地的植被类型发生了显著变化。坡面植被的变化, 不但对坡面土壤养分的分布和吸收利用等产生影响[16], 而且直接改变了坡面不同位置的水热环境和生物学(土壤微生物、植物根系等)特征[17], 从而对土壤养分循环过程(特别是氮素的转化)产生影响[18], 并对坡面植被产生反馈作用。此外, 目前开展的植被类型对土壤养分循环的影响研究中, 很少考虑采样位置和坡位因素的影响, 而不同坡位、植被周围不同位置土壤养分含量及植物根系分布差异巨大, 必将对养分转化产生影响[19]。
本文以黄土高原北部水蚀风蚀交错带和农牧交错带核心地带的柠条()和油松()坡地为对象, 研究了不同坡位植被冠层下和冠层外土壤矿质氮储量和氮素矿化动态特征, 以分析植被类型、坡位和采样位置对坡地土壤氮素转化的影响, 从而为生态环境脆弱区植被恢复条件下土壤氮素有效性评价提供科学依据。
1 材料与方法
1.1 研究区概况
本研究在中国科学院水利部水土保持研究所神木侵蚀与环境试验站六道沟小流域进行, 研究区位于黄土高原水蚀风蚀交错带和农牧交错带的核心位置, 属半干旱大陆性气候, 年均气温8.4 ℃, 月均最低气温-9.7 ℃(1月), 最高气温23.7 ℃(7月), 无霜期169 d, 年均降水量437.4 mm, 主要集中在6—9月份(占全年降水量的77.4%), 年蒸发量785.4 mm。研究区生态环境脆弱, 植被退化严重[20]。20世纪70年代末期, 为了遏制土壤侵蚀并改善生态环境, 该区的坡耕地开始弃耕, 之后自然演替为以当地优势草种本氏针茅()为主的草地; 80年代初期开始在弃耕草地大量种植柠条和油松。研究区土壤类型包括干润砂质新成土、淤积人为新成土和侵蚀砂壤质新成土, 土壤容重介于1.26~1.53 g×cm-3, 土壤砂粒含量较高(66%~80%), 黏粒含量很低(9%~15%), 土壤质地为砂壤土。
1.2 试验设计和氮素矿化速率测定
2008年4月在六道沟小流域选取退耕年限相近的油松坡地和柠条坡地各3个。每个坡地按上、中、下坡位分别设置样地(10 m×10 m), 在每个样地设置冠层下和冠层外采样小区各3个。在每个小区用环刀法测定0~10 cm和10~20 cm土层土壤容重, 用原位培养法测定生长季(4—10月)土壤氮素矿化速率。
测定时, 在每个小区选取5个点, 将直径5 cm、长22 cm的聚氯乙烯管筒(矿化管)垂直压进土体, 管上端留2 cm, 用透气薄膜覆盖并扎住, 原位培养1个月左右取出矿化管, 之后将5个点土样混匀分别测定0~10 cm和10~20 cm土层土壤矿质氮(硝态氮和铵态氮)含量, 同时测定土壤含水量, 并将下一批矿化管按同样方法埋在上批矿化管附近。每次埋设矿化管时, 在矿化管周围采集0~10 cm和10~20 cm土层土壤样品, 测定矿质氮含量, 作为培养期初始矿质氮含量, 并用其分析土壤矿质氮动态变化。利用培养前后矿质氮含量的差异和实际培养天数计算土壤氮素净矿化速率。
本研究分别在4月25日(T1¢)、5月25日(T2¢)、6月21日(T3¢)、7月22日(T4¢)、8月23日(T5¢)和10月15日(T6¢)测定土壤矿质氮含量, 在4月25日—5月25日(T1)、5月25日—6月21日(T2)、6月21日—7月22日(T3)、7月22日—8月23日(T4)和8月23日—10月15日(T5)测定土壤氮素矿化速率。采集的土壤样品(包括原位培养前和培养后的样品)装入样品袋中, 带回实验室测定矿质氮含量。土壤矿质氮含量用2 mol×L-1KCl浸提—流动分析仪(AA3, 德国Seal公司)测定。5个培养期降水量分别为21.7 mm、63.5 mm、19.1 mm、150.6 mm和140.5 mm, 平均气温分别为12.1 ℃、21.0 ℃、23.9 ℃、20.6 ℃和9.9 ℃。
1.3 数据处理
土壤硝态氮、铵态氮和总矿质氮储量计算公式为:
=C´´´10–2(1)
式中:C为各土层硝态氮、铵态氮和总矿质氮含量(mg×kg-1),为土壤容重(g×cm-3),为土层厚度(cm),为相应的矿质氮储量(g×m-2)。
土壤矿化、硝化和铵化速率计算公式为:
NR=(S+1–S)/(t+1–t) (2)
AR=(¢+1¢)/(t+1–t) (3)
MR=NR+AR (4)
式中: NR、AR和MR分别为硝化速率、铵化速率和矿化速率(g×m-2×d-1),与¢分别为相应时期硝态氮与铵态氮的储量,t与t1为培养前后时间。
本研究中用马氏距离(Mahalanobis distance)法去除异常值(马氏距离大于2.0的测定结果为异常值), 异常值判定用JMP(10.0)软件进行; 用Microsoft Excel 2007进行数据的原始计算, 用SPSS 22.0软件进行统计分析和作图; 用成对检验法(Paired T test)表示不同采样位置的差异显著性; 用多元方差分析检验植被类型、采样位置和采样(培养)时期或坡位对土壤硝态氮、铵态氮和矿质氮储量及硝化速率、铵化速率和矿化速率的影响。
2 结果与分析
2.1 生长季土壤水分对林型、坡位和采样位置的动态响应特征
研究区土壤水分含量呈现出明显的季节动态特征, 在生长季前期(T1和T2)和末期(T4和T5)较高, 生长季中期(T3)较低(图1a)。如生长季前期、中期和后期土壤水分含量在0~10 cm土层分别介于2.79%~13.89%、2.48%~7.65%和2.36%~14.42%, 在10~20 cm土层分别介于3.17%~16.23%、4.70%~ 10.06%和3.97%~16.67%。土壤水分含量的季节动态受采样位置和土层的影响。总体上0~10 cm土层土壤水分变异系数(38%~51%)大于10~20 cm土层(27%~36%), 而且冠层外的变异系数(31%~51%)大于冠层内(27%~40%), 表明冠层对降水的截留作用, 使其下土壤和深层土壤对水分的变化有着较为明显的缓冲作用。此外, 不同采样位置土壤水分含量变化还与坡位有关, 如上坡位和中坡位冠层下土壤水分含量在2个土层均低于冠层外土壤, 而下坡位则与此相反(图1b)。
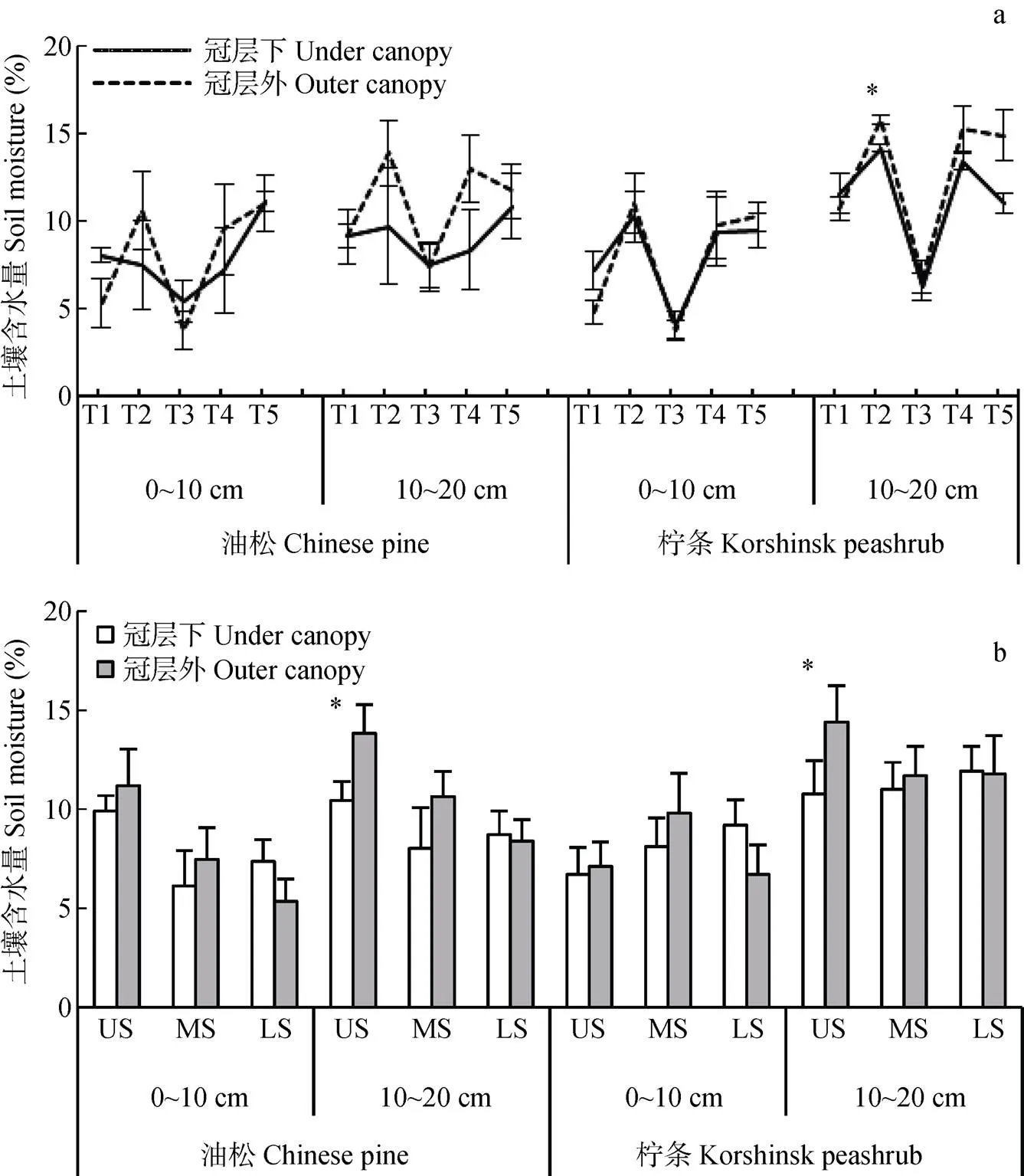
图1 生长季不同时期(a)和不同坡位(b)油松及柠条坡地冠层内、外土壤水分含量特征
T1: 4月25日—5月25日; T2: 5月25日—6月21日; T3: 6月21日—7月22日; T4: 7月22日—8月23日; T5: 8月23日—10月15日。US: 上坡位; MS: 中坡位; LS: 下坡位。图中*表示冠层内外含水量差异显著(<0.05)。T1, T2, T3, T4 and T5 represent 25th April to 25th May, 25th May to 21st June, 21st June to 22nd July, 22nd July to 23rd August, 23rd August to 15th October, respectively. The US, MS and LS represent upper slope, middle slope and lower slope, respectively. The * in the figures represent significant difference at< 0.05 between under canopy and outer canopy.
2.2 生长季土壤矿质氮对林型、坡位和采样位置的动态响应特征
研究区植物生长季土壤矿质氮以铵态氮为主, 其在0~10 cm和10~20 cm土层铵态氮储量分别占矿质氮总量的61%和70%; 随生长季推移, 铵态氮的比例逐渐升高, 分别从生长季初期的37%和54%增长到生长季末期的75%和82%(图2a)。此外, 油松冠层下和冠层外土壤铵态氮占矿质氮的比例接近(分别为69%和71%), 而柠条冠层下铵态氮比例显著低于冠层外(分别为54%和67%)(图2a)。
土壤铵态氮在生长季中期较高(图2a), 而且不受采样位置的影响, 但受坡位及其与林型之间交互作用的显著影响(表1)。对于油松林来说, 上坡位和中坡位土壤铵态氮(0.37 g×m-2和0.42 g×m-2)显著高于下坡位土壤(0.27 g×m-2); 对于柠条林来说, 不同坡位土壤铵态氮差异不显著(图3a)。此外, 不同类型林地冠层下和冠层外土壤铵态氮差异也不显著(表1, 表2), 土壤硝态氮储量随生长季推移逐渐降低, 而且油松林冠层下2个土层硝态氮(0~0.61 g×m-2和0~1.09 g×m-2)和冠层外相近(0~0.85 g×m-2和0~0.94 g×m-2), 而柠条林冠层下相应土层硝态氮(0.08~1.08 g×m-2和0.06~0.97 g×m-2)则高于冠层外土壤(0.06~0.92 g×m-2和0.03~0.68 g×m-2)(图2b)。此外, 本研究土壤硝态氮不受坡位的影响, 如上、中、下坡位硝态氮在0~10 cm土层分别介于0.02~0.92 g×m-2、0~1.07 g×m-2和0~1.09 g×m-2, 在10~20 cm土层分别介于0~0.94 g×m-2、0~1.09 g×m-2和0.04~1.08 g×m-2(图3b)。
土壤矿质氮整体上也呈现出随生长季推移而降低的趋势(图2), 如各个处理矿质氮平均值在0~10 cm和10~20 cm土层分别从生长季初期的0.80 g×m-2和0.56 g×m-2降低到生长季末期的0.53 g×m-2和0.50 g×m-2。此外, 土壤总矿质氮受植被类型、采样位置和坡位的共同影响, 柠条林地0~10 cm和10~20 cm土层土壤矿质氮显著高于油松林地(分别为31%和28%), 且柠条冠层下土壤显著高于冠层外土壤(分别为18%和50%)(图2和图3)。
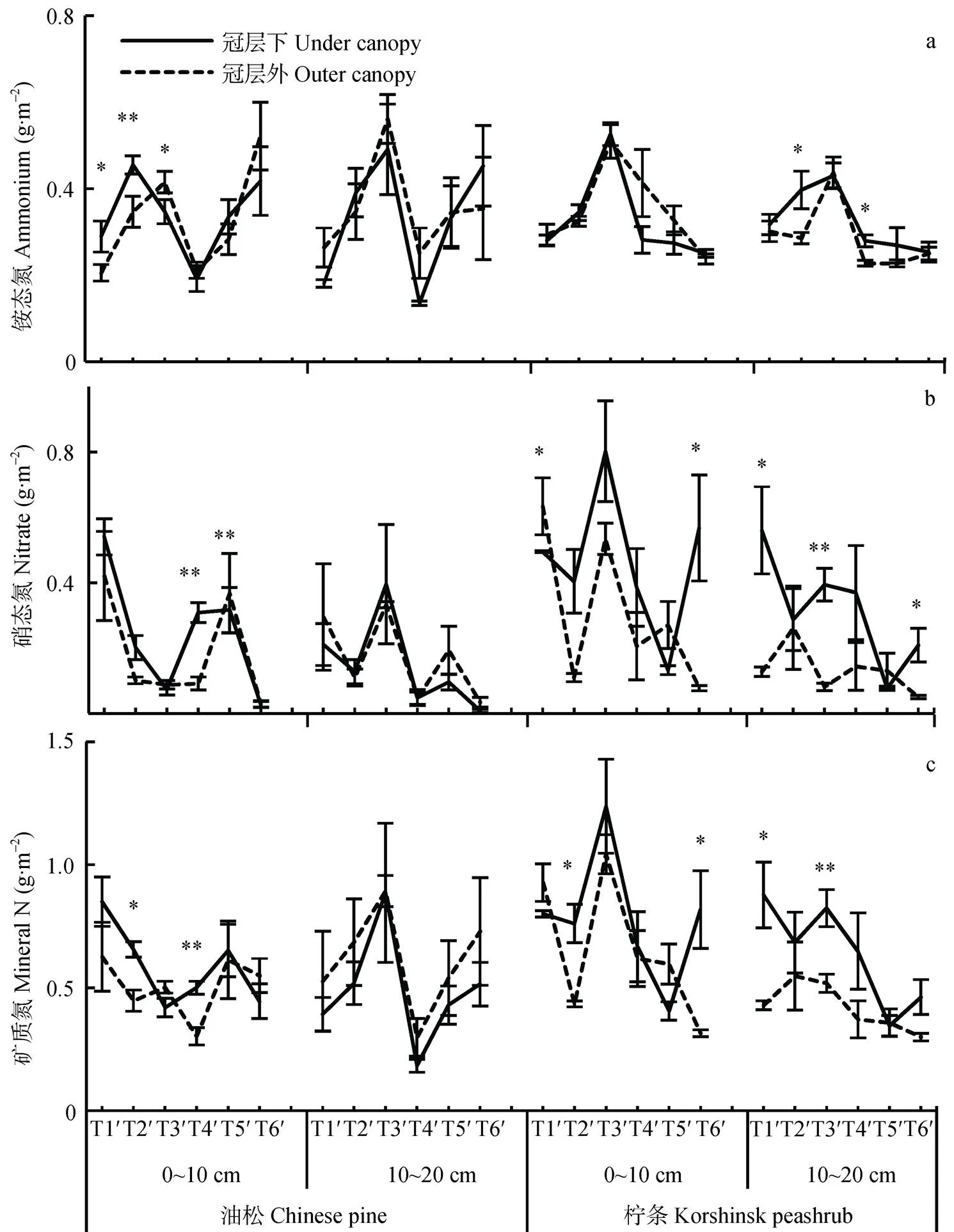
图2 生长季不同时期油松和柠条坡地冠层内和冠层外土壤铵态氮(a)、硝态氮(b)和总矿质氮(c)动态特征
*和**分别表示冠层内外矿质氮储量的差异显著(<0.05)和极显著(<0.01)。T1¢: 4月25日; T2¢: 5月25日; T3¢: 6月21日; T4¢: 7月22日; T5¢: 8月23日; T6¢: 10月15日。* and ** in the figures represent significant differences at< 0.05 and< 0.01 of soil mineral N between under canopy and outer canopy. T1¢, T2¢, T3¢, T4¢, T5¢and T6¢represent 25th April, 25th May, 21st June, 22nd July, 23rd August and 15th October, respectively.
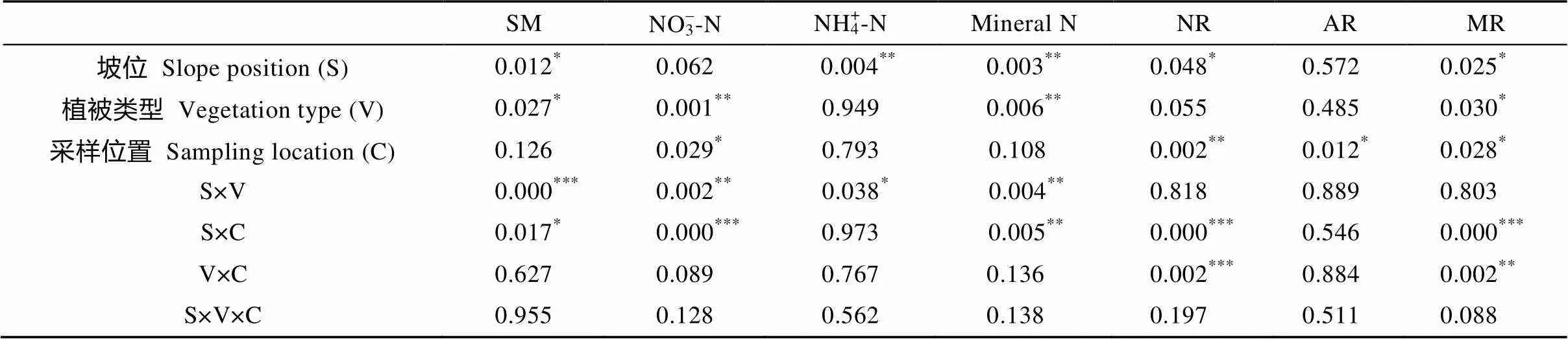
表1 坡位、植被类型和采样位置对生长季土壤含水量、硝态氮、铵态氮、矿质氮以及硝化、铵化、矿化速率影响的方差分析结果(P值)
*代表<0.05, **代表<0.01, ***代表<0.001。SM: 土壤含水量; NR: 硝化速率; AR: 铵化速率; MR: 矿化速率。*, ** and *** represent< 0.05,< 0.01 and< 0.001, respectively. SM: soil moisture; NR: nitrification rate; AR: ammonification rate; MR: mineralization rate.
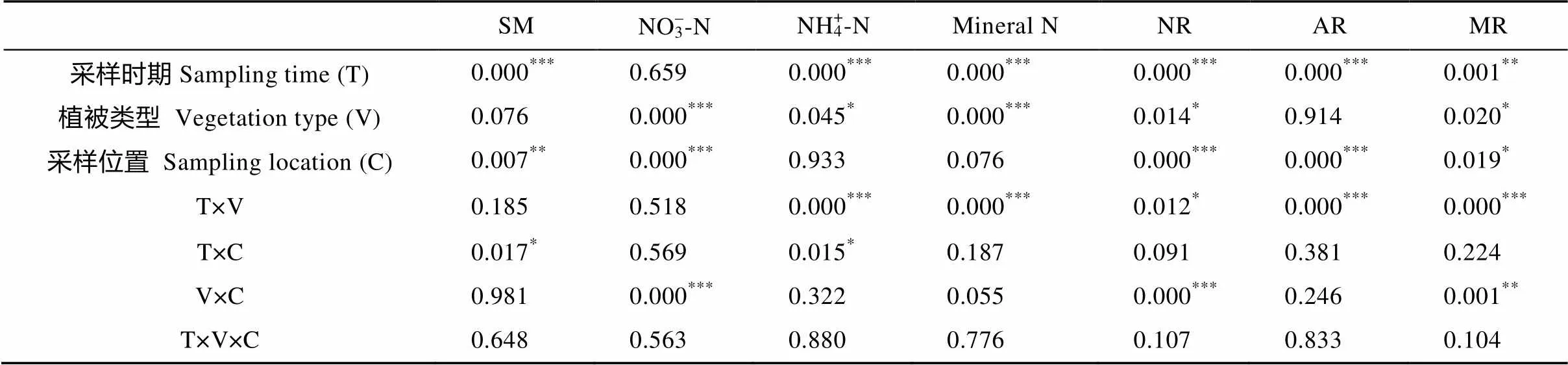
表2 采样时期、植被类型和采样位置对生长季土壤含水量、硝态氮、铵态氮、矿质氮以及硝化速率、铵化速率和矿化速率影响的方差分析结果(P值)
*代表<0.05, **代表<0.01, ***代表<0.001。SM: 土壤含水量; NR: 硝化速率; AR: 铵化速率; MR: 矿化速率。*, ** and *** represent< 0.05,< 0.01 and< 0.001, respectively. SM: soil moisture; NR: nitrification rate; AR: ammonification rate; MR: mineralization rate.
2.3 生长季土壤氮素矿化对林型、坡位和采样位置的动态响应特征
研究区生长季土壤氮素矿化作用在0~10 cm土层主要由硝化作用主导(占氮素矿化速率的94%±16%), 在10~20 cm土层则由硝化和铵化作用共同引起(分别占氮素矿化速率的44%±14%和56%±14%)。此外, 不同土层硝化和铵化作用对矿化速率的贡献在不同林型、采样位置和不同坡位均表现出相似的变化趋势(>0.1)(图4)。
不同林型冠层下和冠层外土壤铵化速率在生长季初期较高(1.18~14.02 mg×m-2×d-1), 中期较低(-11.80~8.64 mg×m-2×d-1)(图4a)。冠层外土壤铵化速率在0~10 cm土层高于冠层下, 在10~20 cm土层与冠层下接近(图4a, 图5a)。油松冠层下和冠层外以及柠条冠层下土壤铵化速率随坡位降低逐渐增大或者由负值逐渐变为正值; 柠条冠层外土壤在不同坡位均表现出铵化作用, 而且铵化速率以坡中部最高, 坡上部最低(图5a)。
本研究中土壤硝化速率的季节动态特征与土层、林型和采样位置有关(图4b)。油松林地0~10 cm土层硝化速率在生长季中期(3.02~34.87 mg×m-2×d-1)显著高于生长季初期(-65.47~-7.85 mg×m-2×d-1)和末期(-7.00~4.63 mg×m-2×d-1), 且均高于10~20 cm土层硝化速率(-14.12~3.03 mg×m-2×d-1), 但冠层下与冠层外差异不显著。柠条林生长季大多数时期0~10 cm土层土壤硝化速率(4.97~48.11 mg×m-2×d-1)高于10~20 cm土层(-6.32~27.31 mg×m-2×d-1), 且冠层下土壤(-0.68~32.51 mg×m-2×d-1)高于冠层外土壤(-6.29~6.90 mg×m-2×d-1)。土壤硝化速率还受坡位影响, 2种林型冠层下硝化速率以下坡位最高, 而冠层外则以下坡位最低(图5b)。
由于0~10 cm土层氮素矿化主要由硝化作用引起, 而10~20 cm土层铵化作用占比例较大, 不同林型和采样位置下矿化速率在0~10 cm土层呈现出与硝化速率相似的季节动态特征, 在10~20 cm土层则呈现出与铵化速率相似的季节动态特征。油松林0~10 cm土层矿化速率在生长季中期显著高于生长季初期和末期,且均高于10~20 cm土层, 但冠层下与冠层外差异不显著(图4c)。柠条林生长季大多数时期0~10 cm土层矿化速率高于10~20 cm土层, 且冠层下高于冠层外(图4和图5)。2种林型冠层下矿化速率以下坡位最高, 而冠层外则以下坡位最低(图5c)。
3 讨论
3.1 生长季土壤无机氮库的季节动态
黄土高原北部水蚀风蚀交错区生长季土壤无机氮库以铵态氮主导, 这与研究区土壤质地较粗有关。研究区土壤砂粒含量介于66%~80%, 具有较高的入渗速率[21]。在降雨过程中, 雨水很容易从地表向深层土壤运动, 从而造成表层土壤硝态氮的淋溶损失, 使其在表层土壤的相对储量显著降低, 而铵态氮的相对储量显著增加。
不同林型冠层下和冠层外土壤铵态氮差异不显著, 可能与不同采样位置下土壤有机质含量接近有关。随有机质含量的增加, 土壤阳离子交换位点逐渐增加, 对铵态氮的吸附能力增强, 土壤铵态氮储量增加[11]。但是研究区冠层下和冠层外土壤有机质差异不显著[19], 因此铵态氮差异也不显著。与此相似, 研究区油松林上坡位和中坡位有机质显著高于下坡位[19], 土壤铵态氮也因此显著高于下坡位。此外, 油松林下坡位植被生长状况优于上坡位和中坡位, 其对下坡位土壤铵态氮的吸收消耗也较大, 土壤中剩余的铵态氮因此较低。对于柠条林地来说, 由于豆科植物固定的氮素是土壤铵态氮的最直接来源[22], 其对土壤铵态氮的影响大于植物吸收利用和不同坡位水热环境差异的影响, 而且不同坡位柠条生长状况差异不显著[19], 因此柠条林不同坡位铵态氮差异不显著。
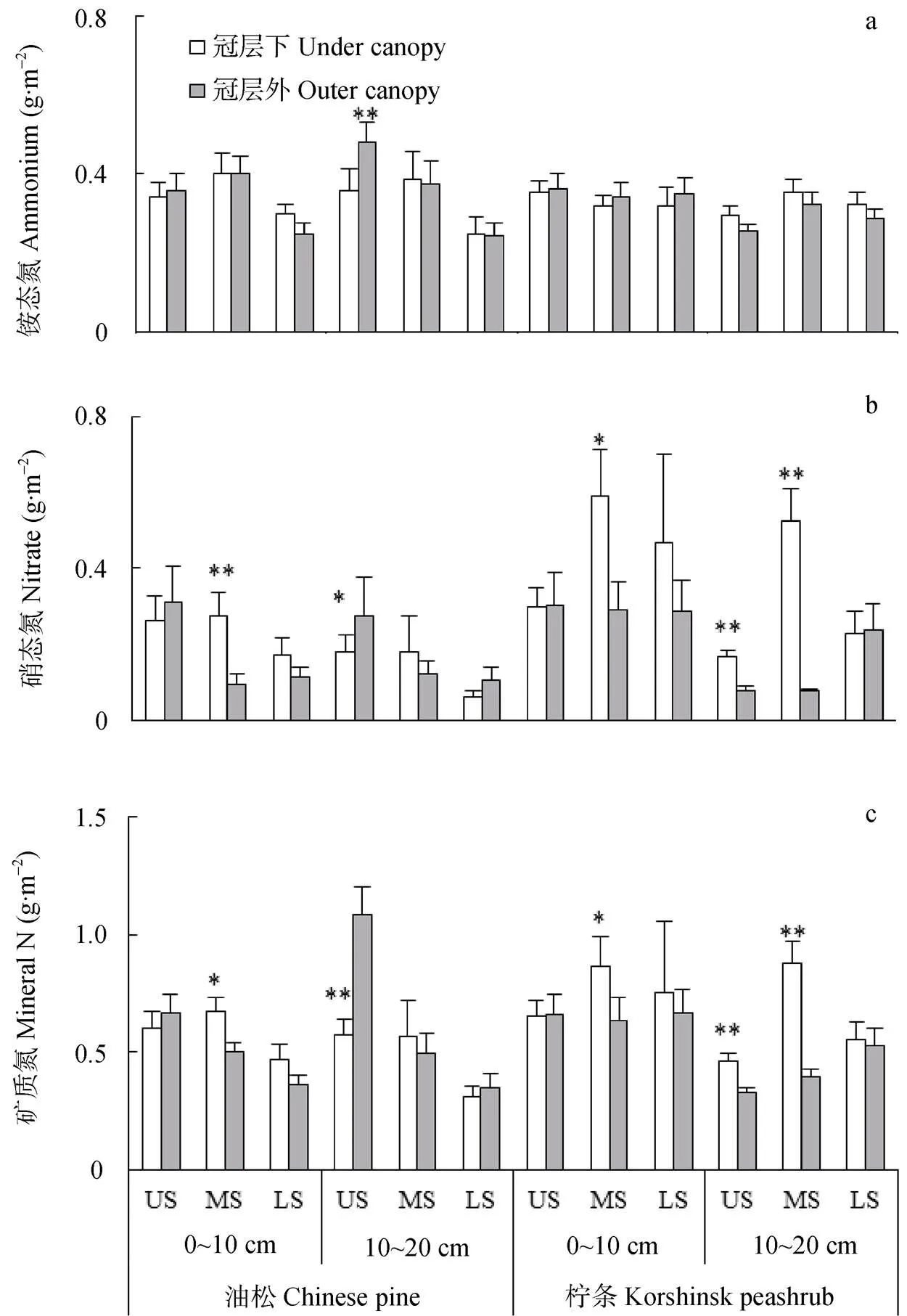
图3 生长季不同坡位油松和柠条坡地冠层内和冠层外土壤铵态氮(a)、硝态氮(b)和总矿质氮(c)动态特征
Fig. 3 Effects of slope position on contents of soil ammonium (a), nitrate (b) and total mineral N (c) across growing season in soils under canopy and outer canopy of the Chinese pine and korshinsk peashrub slopes
US: 上坡位; MS: 中坡位; LS: 下坡位。图中*和**分别表示冠层内外矿质氮储量的差异显著(<0.05)和极显著(<0.01)。The US, MS and LS represent upper slope, middle slope and lower slope, respectively. The * and ** in the figures represent significant at< 0.05 and< 0.01 between under canopy and outer canopy.
硝态氮和矿质氮的季节动态特征与林型和采样位置有关, 由于生长季植被的吸收作用, 土壤硝态氮和矿质氮逐渐降低, 这与肖好燕等[23]在亚热带地区的研究结果一致。油松林冠层下和冠层外硝态氮和矿质氮差异不显著, 主要因为研究区油松根系较深[24], 对表层土壤氮素吸收利用较弱。柠条林冠层下硝态氮和矿质氮显著高于冠层外, 这与柠条根系固氮作用有关。柠条为豆科植物, 生长过程中可以固定大气中的氮素, 不但能够满足植物生长对氮素的需求, 还向土壤中释放大量的氮素。柠条地上部分氮素含量也显著高于油松, 其凋落物向土壤的归还, 以及归还之后的矿化分解, 会显著增加土壤无机氮的储量。因此柠条林冠层下土壤硝态氮和矿质氮显著高于冠层外土壤。此外, 本研究中不同坡位硝态氮差异不显著, 表明其季节动态主要受植被作用的影响。
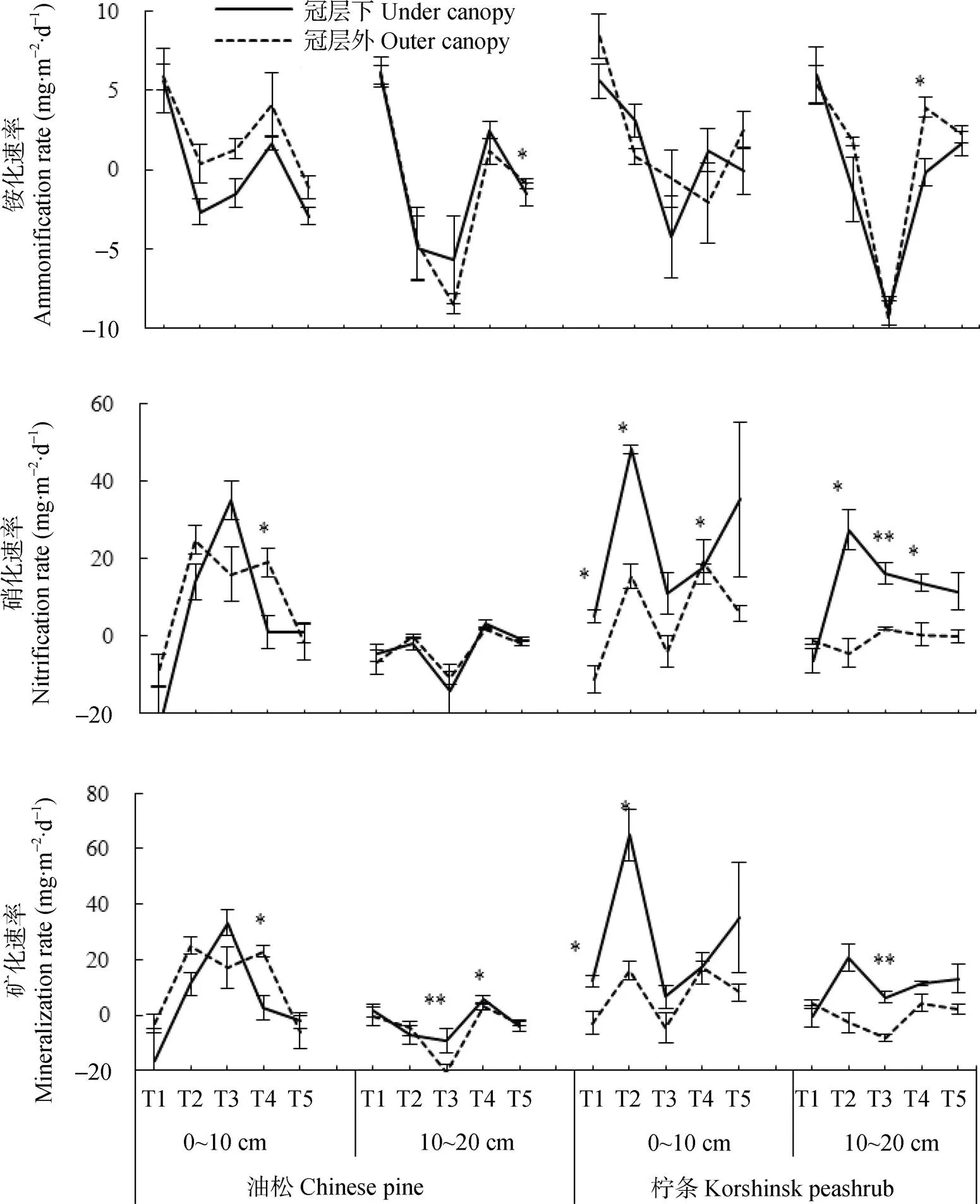
图4 生长季不同时期油松和柠条坡地冠层内和冠层外土壤铵化速率(a)、硝化速率(b)和矿化速率(c)动态特征
图中*和**分别表示冠层内外铵化速率、硝化速率、矿化速率的差异显著(<0.05)和极显著(<0.01)。T1: 4月25日—5月25日; T2: 5月25日—6月21日; T3: 6月21日—7月22日; T4: 7月22日—8月23日; T5: 8月23日—10月15日。* and ** in the figures represent significant at< 0.05 and< 0.01 between under canopy and outer canopy. T1, T2, T3, T4 and T5 represent 25th April to 25th May, 25th May to 21st June, 21st June to 22nd July, 22nd July to 23rd August, 23rd August to 15th October, respectively.
3.2 生长季土壤氮素矿化季节动态
本研究中0~10 cm土层土壤硝化作用占主导地位, 而10~20 cm土层硝化和铵化作用共同主导着矿化过程, 这与不同土层土壤空气中氧气含量有关。0~10 cm土层土壤与大气之间气体交换较为强烈, 土壤空气中氧气含量较高[25], 土壤长期处于氧化状态, 利于硝化作用的进行; 而10~20 cm土层土壤与大气之间气体交换较弱, 且土壤含水量较高, 铵化作用逐渐增强, 其对矿化的贡献也逐渐增加。这也与本研究中铵化速率与土壤含水量显著正相关的结果(=0.044,=231,=0.033 8)一致。
土壤水分和温度对氮素矿化有着显著影响, 硝化速率随温度的升高而增加[26], 铵化速率也随温度和含水量的升高而增加, 但更取决于土壤水分条件[27]。在黄土高原北部水蚀风蚀交错带, 生长季中期土壤温度较高, 而降水和土壤含水量较低, 在温度和水分的交互作用下, 硝化速率呈现较高水平, 而铵化速率呈现较低水平, 并影响着净矿化速率的季节动态。此外, 生长季中期铵化速率较低, 可能也与这一时期较为活跃的微生物活动造成的铵态氮的生物固定有关[28]。
本研究中铵化速率受采样位置和林型的影响较小, 而硝化和矿化速率受这2个因素的显著影响。硝化与矿化速率与不同林型下植物根系分布有关。油松根系分布较深, 0~20 cm土层根系生物量较少[24], 因此采样位置对油松林土壤硝化和矿化速率没有影响。柠条林根系分布较浅[29], 冠层下根系生物量显著高于冠层外土壤, 而且由于柠条的固氮作用, 冠层下土壤具有较低的C/N值, 利于氮素矿化[30]。此外, 固氮作用造成的氮素在冠层下土壤的积累也会促进土壤氮素的矿化从而使硝化和矿化速率显著高于冠层外土壤[31]。
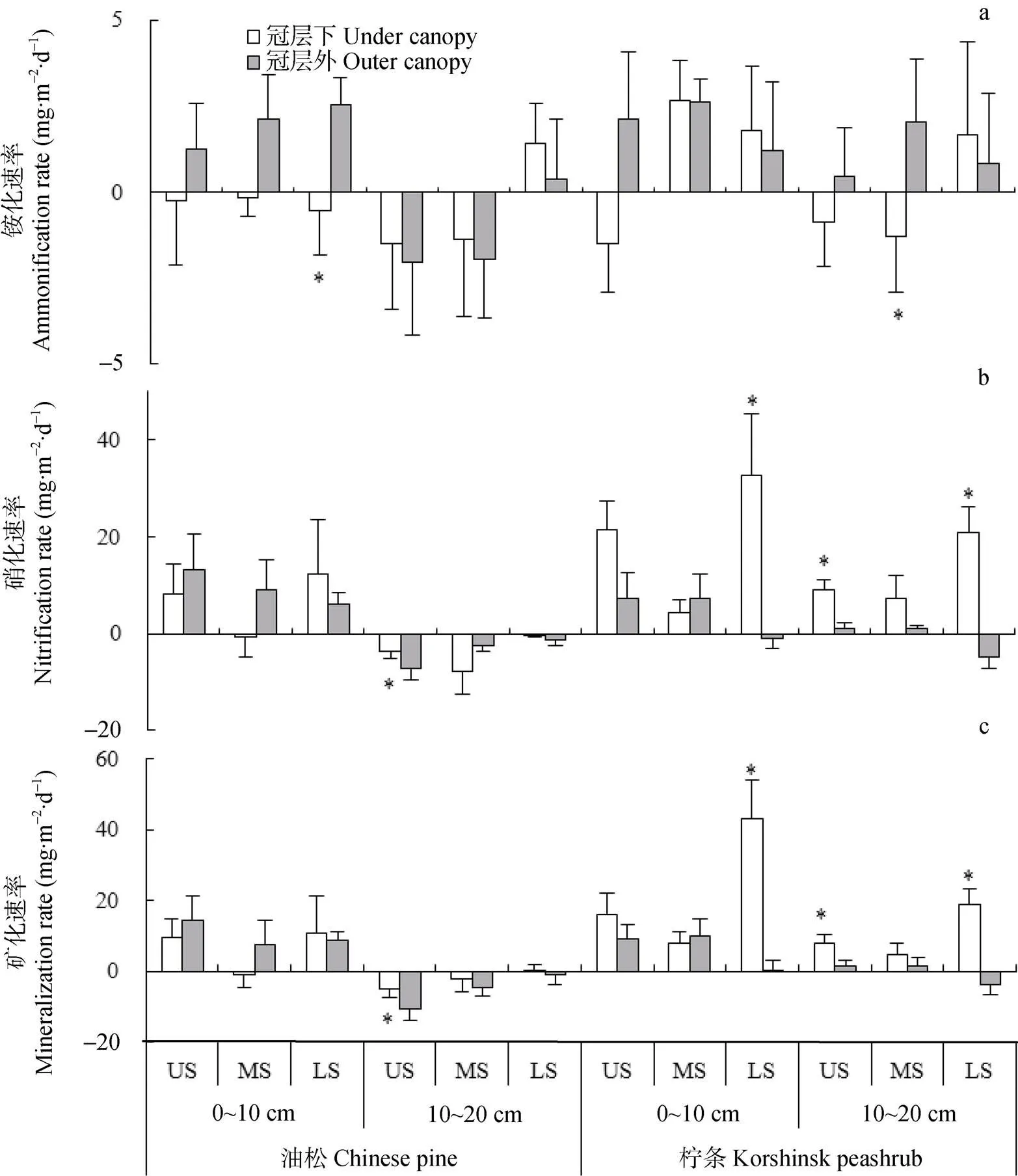
图5 生长季不同坡位油松和柠条坡地冠层内和冠层外土壤铵化速率(a)、硝化速率(b)和矿化速率(c)特征
US: 上坡位, MS: 中坡位, LS: 下坡位; 图中*表示冠层内外铵化速率、硝化速率、矿化速率的差异显著(<0.05)。US, MS and LS represent upper slope, middle slope and lower slope, respectively. * in the figures represent significant at< 0.05 between under canopy and outer canopy.
随着坡位的降低, 土壤温度逐渐降低, 微生物对铵的固定作用逐渐减弱, 铵化作用有所增强。硝化和矿化速率与坡位的关系则受林型和采样位置的影响。冠层下硝化和矿化速率以下坡位最高, 而冠层外则以下坡位最低, 这可能与不同坡位和采样位置土壤水分状况差异及其造成的土壤硝化细菌活性差异有关[32]。2种林型下, 与同一采样位置其他坡位相比, 冠层内土壤含水量以下坡位较高, 该坡位对应的土壤硝化细菌活性也较高, 硝化和矿化速率也高于其他坡位; 冠层外土壤含水量以下坡位最低, 硝化和矿化速率也因此低于其他坡位。
该研究结果表明, 采样位置、植被类型和地形条件共同影响着黄土高原北部坡地生态系统土壤水分、矿质氮和氮素矿化的季节动态。因此, 在评价土地利用变化或者植被生态恢复后土壤养分循环和转化时, 需要合理设计样品采集方案, 以使研究结果更具有代表性。
4 结论
黄土高原北部水蚀风蚀交错带生长季0~10 cm和10~20 cm土层土壤矿质氮由铵态氮主导, 氮素矿化在0~10 cm土层由硝化作用引起, 在10~20 cm土层则由硝化和铵化作用共同引起。
土壤铵态氮受林型、坡位和采样位置影响较小, 硝态氮和矿质氮与林型和采样位置有关, 油松林冠层下土壤硝态氮与冠层外相近, 柠条林冠层下硝态氮高于冠层外。
土壤铵化、硝化和矿化速率受坡位、林型和采样位置的影响, 油松林地硝化和矿化速率在冠层下与冠层外差异不显著, 柠条林地冠层下土壤高于冠层外土壤; 冠层下硝化和矿化速率以下坡位最高, 冠层外以下坡位最低。
[1] GEISSELER D, HORWATH W R, JOERGENSEN R G, et al. Pathways of nitrogen utilization by soil microorganisms — A review[J]. Soil Biology and Biochemistry, 2010, 42(12): 2058–2067
[2] WARING B G, ADAMS R, BRANCO S, et al. Scale-dependent variation in nitrogen cycling and soil fungal communities along gradients of forest composition and age in regenerating tropical dry forests[J]. New Phytologist, 2016, 209(2): 845–854
[3] 罗亲普, 龚吉蕊, 徐沙, 等. 氮磷添加对内蒙古温带典型草原净氮矿化的影响[J]. 植物生态学报, 2016, 40(5): 480–492 LUO Q P, GONG J R, XU S, et al. Effects of N and P additions on net nitrogen mineralization in temperate typical grasslands in Nei Mongol, China[J]. Chinese Journal of Plant Ecology, 2016, 40(5): 480–492
[4] FIERER N, SCHIMEL J P. Effects of drying–rewetting frequency on soil carbon and nitrogen transformations[J]. Soil Biology and Biochemistry, 2002, 34(6): 777–787
[5] SCHIMEL J, BALSER T C, WALLENSTEIN M. Microbial stress-response physiology and its implications for ecosystem function[J]. Ecology, 2007, 88(6): 1386–1394
[6] 巨晓棠, 李生秀. 土壤氮素矿化的温度水分效应[J]. 植物营养与肥料学报, 1998, 4(1): 37–42 JU X T, LI S X. The effect of temperature and moisture on nitrogen mineralization in soils[J]. Journal of Plant Nutrition and Fertilizer, 1998, 4(1): 37–42
[7] 刘杏认, 董云社, 齐玉春, 等. 温带典型草地土壤净氮矿化作用研究[J]. 环境科学, 2007, 28(3): 633–639 LIU X R, DONG Y S, QI Y C, et al. Soil net nitrogen mineralization in the typical temperate grassland[J]. Environmental Science, 2007, 28(3): 633–639
[8] 王常慧, 邢雪荣, 韩兴国. 温度和湿度对我国内蒙古羊草草原土壤净氮矿化的影响[J]. 生态学报, 2004, 24(11): 2472–2476 WANG C H, XING X R, HAN X G. The effects of temperature and moisture on the soil net nitrogen mineralization in angrassland, Inner Mongolia, China[J]. Acta Ecologica Sinica, 2004, 24(11): 2472–2476
[9] PAUL K, BLACK S, CONYERS M. Development of nitrogen mineralisation gradients through surface soil depth and their influence on surface soil pH[J]. Plant and Soil, 2001, 234(2): 239–246
[10] 傅声雷, 蚁伟民, 丁明懋. 鼎湖山不同植被类型下土壤微生物养分的矿化[J]. 植物生态学报, 1995, 19(3): 217–224 FU S L, YI W M, DING M M. Mineralization of soil microbial C, N, P and K in different vegetation types at Dinghushan biosphere reserve[J]. Acta Phytoecologica Sinica, 1995, 19(3): 217–224
[11] MUELLER K E, HOBBIE S E, TILMAN D, et al. Effects of plant diversity, N fertilization, and elevated carbon dioxide on grassland soil N cycling in a long-term experiment[J]. Global Change Biology, 2013, 19(4): 1249–1261
[12] 唐克丽. 中国水土保持[M]. 北京: 科学出版社, 2004 TANG K L. Soil and Water Conservation in China[M]. Beijing: Science Press, 2004
[13] 唐克丽. 中国土壤侵蚀与水土保持学的特点及展望[J]. 水土保持研究, 1999, 6(2): 2–7 TANG K L. Characteristics and perspectives on scientific discipline of soil erosion and soil and water conservation in China[J]. Research of Soil and Water Conservation, 1999, 6(2): 2–7
[14] 田均良. 黄土高原生态建设环境效应研究[M]. 北京: 气象出版社, 2010 TIAN J L. Study on Environmental Effects of Ecological Construction in Loess Plateau[M]. Beijing: China Meteorological Press, 2010
[15] LYU Y H, FU B J, FENG X M, et al. A policy-driven large scale ecological restoration: quantifying ecosystem services changes in the Loess Plateau of China[J]. PLoS One, 2012, 7(2): e31782
[16] 黄闰泉, 刘贵开, 袁传武, 等. 三峡库区坡面农林复合结构对土壤养分分布的影响[J]. 水土保持学报, 2000, 14(3): 41–45 HUANG R Q, LIU G K, YUAN C W, et al. Impact of agroforestry structure of slopeland on distribution of soil nutrient in Three Gorges Reservoir area[J]. Journal of Soil and Water Conservation, 2000, 14(3): 41–45
[17] 王巧利, 贾燕锋, 王宁, 等. 黄土丘陵沟壑区自然恢复坡面植物根系的分布特征[J]. 水土保持研究, 2012, 19(5): 16–22 WANG Q L, JIA Y F, WANG N, et al. Distribution of root on vegetation recovery slope in the hilly and gully Loess Plateau[J]. Research of Soil and Water Conservation, 2012, 19(5): 16–22
[18] 冯书珍, 苏以荣, 秦新民, 等. 喀斯特峰丛洼地土壤剖面微生物特性对植被和坡位的响应[J]. 生态学报, 2013, 33(10): 3148–3157 FENG S Z, SU Y R, QIN X M, et al. Responses of soil microbial properties in soil profile to typical vegetation pattern and slope in karst-cluster depression area[J]. Acta Ecologica Sinica, 2013, 33(10): 3148–3157
[19] WEI X R, SHAO M G, FU X L, et al. Changes in soil organic carbon and total nitrogen after 28 years grassland afforestation: effects of tree species, slope position, and soil order[J]. Plant and Soil, 2010, 331(1/2): 165–179
[20] 唐克丽, 侯庆春, 王斌科, 等. 黄土高原水蚀风蚀交错带和神木试区的环境背景及整治方向[J]. 中国科学院水利部西北水土保持研究所集刊, 1993, (2): 2–11TANG K L, HOU Q C, WANG B K, et al. The environment background and administration way of wind-water erosion crisscross region Shenmu experimental area on the Loess Plateau[J]. Research of Soil and Water Conservation, 1993, (2): 2–11
[21] 郑纪勇, 邵明安, 张兴昌. 黄土区坡面表层土壤容重和饱和导水率空间变异特征[J]. 水土保持学报, 2004, 18(3): 53–56 ZHENG J Y, SHAO M A, ZHANG X C. Spatial variation of surface soil’s bulk density and saturated hydraulic conductivity on slope in loess region[J]. Journal of Soil and Water Conservation, 2004, 18(3): 53–56
[22] PATRIARCA E J, TATÈ R, IACCARINO M. Key role of bacterial NH4+metabolism in rhizobium-plant symbiosis[J]. Microbiology and Molecular Biology Reviews, 2002, 66(2): 203–222
[23] 肖好燕, 刘宝, 余再鹏, 等. 亚热带不同林分土壤矿质氮库及氮矿化速率的季节动态[J]. 应用生态学报, 2017, 28(3): 730–738 XIAO H Y, LIU B, YU Z P, et al. Seasonal dynamics of soil mineral nitrogen pools and nitrogen mineralization rate in different forests in subtropical China[J]. Chinese Journal of Applied Ecology, 2017, 28(3): 730–738
[24] 刘秀萍, 陈丽华, 陈吉虎. 刺槐和油松根系密度分布特征研究[J]. 干旱区研究, 2007, 24(5): 647–651LIU X P, CHEN L H, CHEN J H. Study on the distribution of root density ofL.andCarr[J]. Arid Zone Research, 2007, 24(5): 647–651
[25] 范爱武. 资源-环境-植物系统中热物理问题的理论与实验研究[D]. 武汉: 华中科技大学, 2004 FAN A W. Theoretical and experimental investigation on thermophysical problems in resources-environment-plant system[D]. Wuhan: Huazhong University of Science and Technology, 2004
[26] 石薇, 王景燕, 魏有波, 等. 水热条件对华西雨屏区柳杉人工林土壤氮矿化的影响[J]. 土壤通报, 2014, 45(6): 1430–1436 SHI W, WANG J Y, WEI Y B, et al. Effects of moisture and temperature on soil nitrogen mineralization ofplantations in rainy area of Western China[J]. Chinese Journal of Soil Science, 2014, 45(6): 1430–1436
[27] 赵长盛, 胡承孝, 孙学成, 等. 温度和水分对华中地区菜地土壤氮素矿化的影响[J]. 中国生态农业学报, 2012, 20(7): 861–866 ZHAO C S, HU C X, SUN X C, et al. Influence of temperature and moisture on nitrogen mineralization in vegetable fields of central China[J]. Chinese Journal of Eco-Agriculture, 2012, 20(7): 861–866
[28] BURGER M, JACKSON L E. Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems[J]. Soil Biology and Biochemistry, 2003, 35(1): 29–36
[29] 朱元龙, 王桑, 林永刚, 等. 黄土高原丘陵区柠条根系生长发育特性研究[J]. 水土保持通报, 2011, 31(2): 232–237 ZHU Y L, WANG S, LIN Y G, et al. Development ofseedling root system in hilly regions of Loess Plateau[J]. Bulletin of Soil and Water Conservation, 2011, 31(2): 232–237
[30] 张名豪, 卢吉文, 赵秀兰. 有机物料对两种紫色土氮素矿化的影响[J]. 环境科学, 2016, 37(6): 2291–2297 ZHANG M H, LU J W, ZHAO X L. Effect of different organic materials on nitrogen mineralization in two purple soils[J]. Environmental Science, 2016, 37(6): 2291–2297
[31] MILCU A, PARTSCH S, SCHERBER C, et al. Earthworms and legumes control litter decomposition in a plant diversity gradient[J]. Ecology, 2008, 89(7): 1872–1882
[32] STARK J M, FIRESTONE M K. Mechanisms for soil moisture effects on activity of nitrifying bacteria[J]. Applied and Environmental Microbiology, 1995, 61(1): 218–221
刘娇, 付晓莉, 李学章, 贾小旭, 邵明安, 魏孝荣. 黄土高原北部生长季土壤氮素矿化对植被和地形的响应[J]. 中国生态农业学报, 2018, 26(2): 231-241
LIU J, FU X L, LI X Z, JIA X X, SHAO M A, WEI X R. Responses of soil nitrogen mineralization during growing season to vegetation and slope position on the northern Loess Plateau of China[J]. Chinese Journal of Eco-Agriculture, 2018, 26(2): 231-241
Responses of soil nitrogen mineralization during growing season to vegetation and slope position on the northern Loess Plateau of China*
LIU Jiao1,2, FU Xiaoli3, LI Xuezhang4, JIA Xiaoxu3, SHAO Ming’an1,2, WEI Xiaorong1,2**
(1. College of Resources and Environment, Northwest A&F University, Yangling 712100, China; 2. State Key Laboratory of Soil Erosion and Dry Land Farming on the Loess Plateau, Northwest A&F University, Yangling 712100, China; 3. Institute of Geographical Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China; 4. Institute of Subtropical Agriculture, Chinese Academy of Sciences, Changsha 410125, China)
Nitrogen (N) mineralization is critical for nitrogen cycle in terrestrial ecosystems and significantly influences the availability of soil N. In this paper, we studied the changes in soil mineral N and N mineralization rates in slope lands in the northern Loess Plateau region in relation to vegetation types, sampling sites and slope positions during vegetation growing season. The objectives of the study were to determine the dynamics of N mineralization during growing season and the influencing factors. Slope lands with Chinese pine () and korshinsk peashrub () were selected in the Liudaogou catchment and anmineralization method was used to measure soil N mineralization for the period from April through October. The measurements were conducted in upper, middle and lower positions of the slope with under- and non-under-canopy at the 0-10 cm and 10-20 cm soil depths. Soil mineral N in the growing season was dominated by ammonium, which accounted for 61% and 70% of total mineral N at the 0-10 cm and 10-20 cm soil depths, respectively. The proportion of ammonium to total mineral N increased during the growing season. Soil ammonium in the upper and middle slope positions was significantly higher than that in the lower slope position for Chinese pine, but not affected by slope positions for korshinsk peashrub. Furthermore, soil ammonium was not affected by sampling site in both Chinese pine and korshinsk peashrub plantations. Soil nitrate and total mineral N were affected by vegetation type and sampling site, rather than by slope position. Under-canopy soil nitrate was similar to that of non-under-canopy in Chinese pine vegetation, but it was greater than that for non-under-canopy in porshinsk peashrub vegetationSoil N mineralization during growing season resulted mainly from nitrification at the 0-10 cm soil depth, but also influenced by both nitrification and ammonification at the 10-20 cm soil depth. Ammonification rate was significantly high during the early growing season and low during the mid growing season. Moreover, ammonification rate was affected by slope position, vegetation type and sampling site. Nitrification and net N mineralization rates in under-canopy soils were similar to those in non-under-canopy soils in Chinese pine vegetation, but it was greater than that in non-under-canopy soils in korshinsk peashrub vegetation. Additionally, when compared with other slope positions, lower slope position had highest nitrification and net mineralization rates of soil N in under-canopy conditions, but it was the lowest in lower slope position in both Chinese pine and korshinsk peashrub plantations. The results suggested that korshinsk peashrub enhanced nitrification and mineralization of N in soils in under-canopy conditions, while the effects of Chinese pine on soil mineral N and N mineralization were not related to sampling site.
Vegetation type; Slope position; Nitrogen mineralization; Mineral nitrogen; Sampling position
, E-mail: xrwei78@163.com
Jul. 9, 2017;
Oct. 9, 2017
10.13930/j.cnki.cjea.170626
S153.6
A
1671-3990(2018)02-0231-11
魏孝荣, 主要从事土壤和生态系统物质循环研究。E-mail: xrwei78@163.com 刘娇, 主要从事土壤物质循环研究。E-mail: walj0522@163.com
2017-07-09
2017-10-09
* This study was funded by the National Natural Science Foundation of China (41571130082, 41571296 and 41622105) and the Key Research Project of Frontier Science of the Chinese Academy of Sciences (QYZDB-SSW-DQC039).
* 国家自然科学基金项目(41571130082, 41571296和41622105)和中国科学院前沿科学重点研究计划项目(QYZDB-SSW-DQC039)资助
猜你喜欢
土壤学报(2022年2期)2022-06-09
河南师范大学学报(自然科学版)(2022年3期)2022-04-14
现代农村科技(2021年5期)2021-12-07
乡村科技(2021年16期)2021-10-21
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
中国煤炭工业(2020年1期)2020-12-28
农业环境科学学报(2020年10期)2020-11-11
当代陕西(2019年7期)2019-04-25
绿色科技(2017年3期)2017-03-14
绿色科技(2017年1期)2017-03-01
