
Tum-5基因对肝细胞癌生长的抑制作用和对血管生成的影响
2018-01-24 08:43关新刚王英楠徐聪睿盖晓东
吉林大学学报(医学版) 2018年1期
历 春,关新刚,王英楠,徐聪睿,盖晓东
(北华大学基础医学院免疫学教研室,吉林 吉林 132013)
实体瘤的生长和转移依赖于新生血管的形成,因此肿瘤血管生成是实体瘤侵袭转移的关键步骤[1-2]。肝细胞癌(hepatocellular carcinoma,HCC)是人类常见的恶性肿瘤之一,肿瘤血管丰富,恶性程度高,发展迅速,生存率低。因此,寻找特异性抗血管生成的靶点成为提高HCC患者生存率的研究热点。肿瘤抑素(tumstatin,Tum)属于血管基底膜Ⅳ型胶原蛋白α3链的一部分,是Saus等[3]在20世纪80年代作为肺-肾出血综合征的自身抗原而被发现,具有抗血管生成和抗肿瘤活性。Tum是目前发现活性最强的内源性血管生成抑制因子,可通过特异性地抑制血管内皮细胞蛋白的合成,导致内皮细胞特异性凋亡,使新生血管合成受到抑制,从而抑制肿瘤细胞的生长、浸润和转移[4-6]。Tum-5片段是Tum抗血管生成的活性片段,位于第54~132位氨基酸之间,具有与全长Tum相同的抗血管生成活性[7],且不含肺-肾出血综合征的抗原表位。
本课题组前期研究人Tum-5目的基因,构建了逆转录病毒pLXSN-Tum-5重组体,使用包装细胞PA317将其包装成假病毒颗粒感染NIH3T3细胞,获得了pLXSN-Tum-5病毒颗粒[8]。前期实验[9]证实:Tum-5可明显抑制人脐静脉内皮细胞(HUVEC)的增殖,通过下调Bcl-2、促进Bax和Caspase-3表达诱导HUVEC发生凋亡。在此基础上,本课题以肝癌细胞为研究对象,体外检测Tum-5对人肝癌细胞的增殖抑制作用,体内建立小鼠肝细胞癌移植瘤模型,检测Tum-5对移植瘤生长的作用和对移植瘤内新生血管的影响,以期为临床肿瘤患者提供抗血管生成治疗的新途径。
1 材料与方法
1.1 实验动物、细胞及主要试剂8~12周龄雌性昆明小鼠20只(体质量15~20 g),由吉林大学实验动物中心提供,动物合格证号:SCXK(吉)2015-0005。人肝癌细胞株HepG-2和小鼠肝癌细胞株H22均购自中国科学院细胞库。RPMI-1640购自美国Hyclone公司,新生小牛血清购自杭州四季青公司,MTT 试剂购自美国Sigma公司,兔抗鼠CD31单克隆抗体购自美国eBioscience公司,免疫组织化学试剂盒购自北京中杉试剂有限公司。
1.2 细胞培养HepG-2和H22细胞分别培养于含有 10%新生小牛血清的 RPMI-1640 培养液(含 100 U·mL-1青霉素和 100 mg·L-1链霉素)中,在含有 5% CO2的 37℃孵箱中培养。
1.3 体外细胞增殖实验取处于对数生长期的HepG-2细胞,以8×103个/孔接种96孔板中,每组5个复孔。待细胞贴壁后分别加入不同感染复数(MOI)(0、1、5、10、25 和 50)的pLXSN和pLXSN-Tum-5病毒颗粒,置于37℃、含5% CO2的孵箱中继续培养72 h。每孔加入MTT溶液20 μL(终浓度为0.5 g·L-1),培养4 h后每孔加入DMSO 150 μL,充分溶解后酶标仪检测各孔吸光度(A)值,检测波长590 nm,测定各组细胞增殖率。细胞增殖率=(实验组A值-对照组A值)/对照组A值×100%。
1.4 小鼠肝细胞癌移植瘤模型的构建剔去小鼠右侧腋窝下及周围毛发,碘伏皮肤常规消毒。将对数生长期的H22细胞配制成1×107mL-1的细胞悬液,以RPMI-1640培养基重悬。取0.1 mL分别注射到小鼠右侧腋下已经事先备皮的皮肤下,回抽无血,缓慢注入后迅速拔针,用酒精棉球稍按压进针处皮肤,即可完成接种,约1周左右成瘤,挑选移植瘤体积在30~70 mm3的小鼠分组实验,移植瘤模型成功建立。
1.5 体内抑瘤实验15只荷瘤小鼠随机分成saline(生理盐水)组、pLXSN组和pLXSN-Tum-5组,每组5只。在第 0、2、4、6和8天分别实施瘤内注射saline和病毒颗粒(MOI=5),共注射5次。治疗效果和安全性分别通过测量肿瘤的体积、质量和小鼠体质量来评估。每2 d用游标卡尺测量肿瘤的长径(a)和短径(b),按公式V = ab2/2计算肿瘤体积,绘制肿瘤体积-时间生长曲线。10 d后,采用颈椎离断处死荷瘤鼠,完整剥离出瘤体,电子天平称质量,计算各组瘤体质量的平均值。
1.6 免疫组织化学染色及结果判定小鼠肝癌移植瘤组织石蜡包埋后制备成5 μm切片,CD31单克隆抗体标记血管内皮细胞。实验步骤参照免疫组织化学试剂盒说明书,PBS代替一抗作阴性对照,计算各组肿瘤微血管密度(MVD)。结果判定:高倍镜(×200)视野下计数高密度区被抗CD31抗体染成棕色的血管数目,每个高密度区计数5个视野下的微血管计数,取其平均值为该标本的MVD。
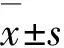
2 结 果
2.1 各组HepG-2细胞的增殖率采用不同MOI(0、1、5、10、25 和 50)的pLXSN-Tum-5和pLXSN病毒颗粒感染HepG-2细胞 72 h检测细胞增殖率。pLXSN-Tum-5组随着MOI的增高HepG-2细胞增殖率无明显变化,与pLXSN组比较差异无统计学意义(P> 0.05)。见表1。
表1 各组HepG-2细胞增殖率
Tab.1 Proliferation rates of HepG-2 cells in various groups

表1 各组HepG-2细胞增殖率
GroupProliferationrate(MOI)015102550pLXSN 10098.0±4.296.0±5.195.0±4.697.0±3.894.0±5.0pLXSN⁃Tum⁃5 10097.0±3.893.0±5.492.0±6.295.0±4.696.0±3.5
2.2 各组小鼠移植瘤体积各组荷瘤鼠在实验前6 d皮下移植瘤生长速度未见明显差异;第7天后瘤体生长差异逐渐显现,pLXSN-Tum-5组小鼠皮下移植瘤生长速度明显减慢,滞后于pLXSN组和saline组。治疗结束后,pLXSN-Tum-5组小鼠皮下移植瘤体积明显低于saline组和pLXSN组(P< 0.05 或P< 0.01);pLXSN组与saline组比较差异无统计学意义(P> 0.05)。见图1。
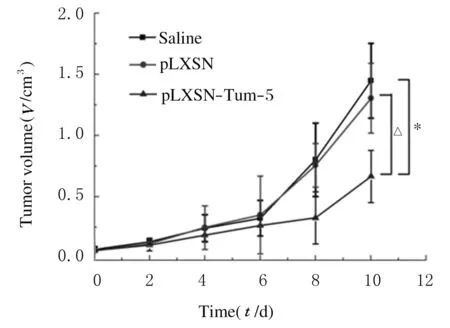
*P<0.01 compared with saline group;△P< 0.05 compared with pLXSN group.
2.3 各组小鼠移植瘤质量治疗10 d后处死荷瘤鼠,切取肿瘤称质量并拍照,pLXSN-Tum-5组肿瘤大小与pLXSN组和saline组相比明显缩小,见图2(插页三)。pLXSN-Tum-5组肿瘤质量明显低于saline组和pLXSN组(P< 0.05或P< 0.01),见表2。

表2 各组小鼠移植瘤质量
*P< 0.01 compared with saline group;△P< 0.05 compared with pLXSN group.
2.4 各组荷瘤鼠体质量隔日测量各组荷瘤鼠体质量,saline组、pLXSN组和pLXSN-Tum-5组荷瘤小鼠在治疗过程中体质量呈稳步增加。治疗10 d后3组小鼠的体质量组间比较差异无统计学意义(P> 0.05)。见图3。
2.5 各组移植瘤组织新生血管形态及MVD移植瘤组织内可见形态不规则的新生血管,血管分布以肿瘤组织边缘处多见。与saline组和pLXSN组比较,pLXSN-Tum-5组小鼠移植瘤组织中新生血管数目明显减少,pLXSN-Tum-5组平均MVD值与saline组和pLXSN组比较明显降低(P< 0.05)。见图4(插页三)和表3。
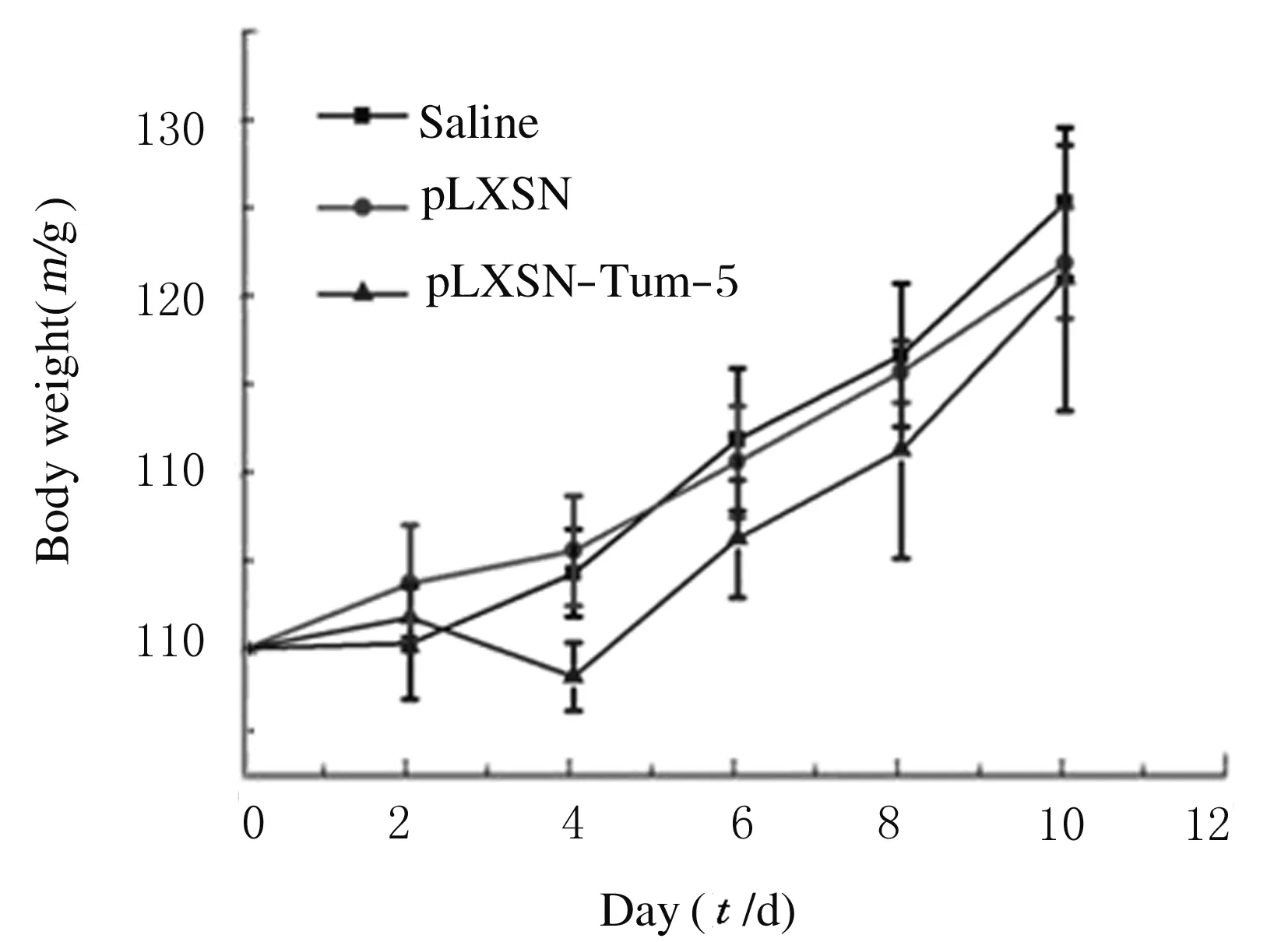
图3 各组荷瘤鼠体质量变化曲线
表3 各组小鼠移植瘤组织平均MVD值
*P< 0.05 compared with saline group;△P< 0.05 compared with pLXSN group.
3 讨 论
研究[10-12]表明:Tum全长基因具有抑制肿瘤细胞增殖和促进肿瘤细胞凋亡的作用。Tum-5作为Tum的抗血管生成活性片段,主要通过特异性抑制血管内皮细胞增殖发挥抗肿瘤活性,而不直接作用于肿瘤细胞[5,7,13]。有研究[14]表明:Tum-5体外可明显抑制胃癌细胞的增殖,促进胃癌细胞凋亡。本研究为了验证Tum-5体外是否对肝癌细胞有直接的增殖抑制作用,选用HepG-2细胞株作为研究对象,选用不同浓度pLXSN-Tum-5和pLXSN病毒颗粒感染HepG-2细胞,检测HepG-2细胞的增殖情况。本研究结果显示:随着pLXSN-Tum-5病毒颗粒浓度的增加,HepG-2细胞的增殖率并无明显变化,与pLXSN对照组比较也无明显变化,表明Tum-5体外对HepG-2细胞没有直接的抗肿瘤活性。考虑这种结果差异可以归因于不同类型的肿瘤细胞。
肿瘤血管生成过程中,血管内皮细胞在肿瘤细胞活化、增殖、迁移和浸润等过程中发挥重要作用[15-16]。 HCC血管丰富,通过特异性抗血管靶向干预抑制上述过程可成为HCC潜在的治疗靶点。因此,本文作者采用小鼠皮下肝癌移植瘤模型,通过局部多点注射pLXSN和pLXSN-Tum-5病毒颗粒来检测Tum-5的体内抗肿瘤效果。治疗5次后,pLXSN-Tum-5组小鼠肿瘤的体积和质量明显低于saline和pLXSN组,主要体现在小鼠皮下移植瘤体积的增大受到明显抑制,并且最终肿瘤质量的增加明显减慢,说明pLXSN-Tum-5组肿瘤细胞的成瘤活性和增殖能力均处于较低水平。此外,治疗结束后3组小鼠体质量无明显差异,说明pLXSN-Tum-5病毒颗粒在体内起到抗肿瘤作用的同时未引起明显的不良反应。
MVD目前被认为是评价肿瘤血管生成的金标准,以抗CD31抗体等标记肿瘤血管内皮细胞,计数单位面积中的微血管数目即为MVD[17-20]。为进一步明确Tum-5对小鼠肝癌移植瘤内血管生成的影响,对各治疗组小鼠的瘤组织进行CD31免疫组织化学染色,检测MVD。本研究结果显示:肿瘤组织内可见形态不规则的新生血管,血管分布以肿瘤组织的边缘处为多见。与saline和pLXSN组比较,pLXSN-Tum-5组小鼠肿瘤组织内的平均MVD明显降低,新生血管密度明显减少,表明Tum-5体内对肝癌组织有明显的抗血管生成作用。
综上所述,Tum-5对肝癌细胞的增殖没有直接的抑制作用,但可通过抑制肝癌组织内新生血管的生成抑制肝癌细胞的生长。Tum-5基因片段可作为一种有效的血管生成抑制剂,为肝癌患者提供新的治疗策略。
[1] 赵 川,李俊萱,刘雪梅,等. 肿瘤血管生成机制的研究进展[J].中华中医药学刊,2017,35(1):130-132.
[2] Bielenberg DR,Zetter BR.The contribution of angiogenesis to the process of metastasis[J].Cancer J,2015,21(4):267-273.
[3] Saus J,Wieslander J,Langeveld JP,et al.Identification of the Goodpasture antigen as the alpha 3(Ⅳ) chain of collagen Ⅳ[J].J Biol Chem,1988,263(26):13374-13380.
[4] Sudhakar A,Boosani CS.Inhibition of tumor angiogenesis by tumstatin:insights into signaling mechanisms and implications in cancer regression[J].Pharm Res,2008,25(12):2731-2739.
[5] Wei C,Xun AY,Wei XX,et al.Bifidobacteria expressing tumstatin protein for antitumor therapy in tumor-bearing mice[J].Technol Cancer Res Treat,2016,15(3):498-508.
[6] Ye HX,Yao Y,Jiang XJ,et al.Tumstatin transfected into human glioma cell line U251 represses tumor growth by inhibiting angiogenesis[J].Chin Med J,2013,126(9):1720-1725.
[7] Maeshima Y,Yerramalla UL,Dhanabal M,et al.Extracellular matrix-derived peptide binds to alpha(v)beta(3) integrin and inhibits angiogenesis[J].J Biol Chem,2001,276(34):31959-31968.
[8] 盖晓东,罗 宏,历 春,等.人Tum-5基因逆转录病毒载体及包装细胞株的构建[J].中国老年学杂志,2009,29(10):1194-1196.
[9] 唐泽波,温 娜,齐 玲,等.pLXSN-Tum-5致人脐静脉内皮细胞凋亡作用及机制[J].中国公共卫生,2017,33(9):1360-1363.
[10]孟昱希,邓志华,牛 鑫.重组慢病毒LV-hTERT-tumstatin的构建及其对人肝癌HepG2细胞增殖和凋亡的影响[J].中国现代医药杂志,2014,16 (5):7-9.
[11]Wang Y,Yin RF,Teng JS.Tumstatin induces apoptosis and stimulates phosphorylation of p65NF-κB in human osteoblastic osteosarcoma Saos-2 cells[J].Oncol Rep,2016,35(6):3403-3408.
[12]王淑静,刘 欢,赵健凯,等.肿瘤抑素改造T-7R肽体外抗肿瘤活性研究[J].中国药理学与毒理学杂志, 2016,30 (10):1064.
[13]Maeshima Y,Colorado PC,Torre A,et al.Distinct antitumor properties of a type IV collagen domain derived from basement membrane[J].J Biol Chem,2000,275(28):21340-21348.
[14]张 军.Tum-5 基因对人胃癌细胞增殖及凋亡影响的研究[D].福州:福建医科大学,2011.
[15]杨 霞.靶向诱导内皮细胞凋亡抑制肿瘤血管新生的研究进展[J].中山大学学报:医学科学版,2017,38 (2):204-214.
[16]Wang YH,Dong YY,Wang WM,et al.Vascular endothelial cells facilitated HCC invasion and metastasis through the Akt and NF-κB pathways induced by paracrine cytokines[J].J Exp Clin Cancer Res,2013,32(1):51.
[17]云 芬,贾永峰,韩 昭,等.HIF-2α在非小细胞肺癌组织中的表达及其与微血管密度、Ki67和GST-π的关系[J].吉林大学学报:医学版,2016,42(5):954-957.
[19]杨霄霄,马 红.肝细胞癌肿瘤标志物的研究进展[J].临床肝胆病杂志,2016,32(4):811-815.
[20]杨 柳,朱曦龄,黎 功.程序性死亡因子配体1在肝细胞癌中的表达及意义[J].临床肝胆病杂志,2016,32(8):1630-1634.
猜你喜欢
现代临床医学(2021年5期)2021-11-02
中国眼镜科技杂志(2019年9期)2019-11-11
天然产物研究与开发(2018年11期)2018-11-30
河北渔业(2018年7期)2018-09-15
天然产物研究与开发(2018年8期)2018-09-10
健康必读·下旬刊(2018年6期)2018-07-24
中成药(2018年3期)2018-05-07
中成药(2017年8期)2017-11-22
天然产物研究与开发(2016年6期)2016-06-05
中国病理生理杂志(2015年8期)2015-12-21
