
四川凉山烟区烟蚜种群空间分布型及抽样技术研究
2018-01-19 09:06王小强刘虹伶蒲德强刘东阳周肆维
中国烟草科学 2017年6期
王小强,刘虹伶,蒲德强*,王 勇,刘东阳,卢 军,周肆维,李 斌,刘 杨
(1.四川省农业科学院经济作物育种栽培研究所,成都 610300;2.四川省农业科学院植物保护研究所,成都 610066;3.四川省烟草公司凉山州公司,四川 西昌 615000;4.四川省烟草公司,成都 610041)
烟蚜(Myzus persicaeSulzer)属半翅目蚜科,亦称桃蚜,是世界范围内危害烟草最严重的害虫之一,在我国各主要烟区均有发生。烟蚜寄主范围极广且可传播病毒,在不同地域表现出黄色型、红色型和绿色型的体色差异。不同色型烟蚜在种群动态、生殖特征、稳定性、寄主选择及抗药性等方面的差异,为烟蚜的单一性化学防治带来困难。目前对烟蚜的防治主要以化学农药为主,但药剂不合理施用使烟蚜长期处于高残留环境中,加上其较强的环境适应能力和较短的生活史,致使敏感性快速减弱,耐药性问题日益严重[1-4]。同时,烟蚜个体小,世代短,繁殖快,数量大,其种群数量和空间分布在不断变化,给调查及测报工作带来了极大的困难。为此,烟蚜空间分布型的研究成为解决这一难题的主要技术手段。空间分布格局是昆虫种群的客观属性,是种群的主要特征之一,不仅对了解昆虫种群的爆发、扩散具有一定的意义,而且在抽样技术及预测预报中也发挥着重要的指导作用[5]。
近年来,夏鹏亮等[6],陈斌等[7],黄本荣等[8],巫厚长等[9],增维爱等[10]利用聚集度指标、Iwaom*-m回归分析法(m*=α+βm)和Taylor幂法则分别对湖北恩施烟区,云南烟区,福建三明烟区,安徽烟区及长沙烟区的烟蚜空间分布型、抽样技术及种群发生动态做了详细研究。而凉山州作为四川省主要烟区,占全省烟草种植面积的60%以上,其气候条件、烟草品种及栽培模式与全国其他烟区存在一定差异。因此,对凉山州烟区烟蚜的空间分布型进行研究有一定实际意义,有助于对烟蚜开展科学的抽样调查和预测预报,为烟蚜的综合治理提供依据。
1 材料与方法
1.1 田间调查
本试验于2016年6—7月在四川省凉山州主要烟区进行,于烟蚜为害高峰期采用五点调查法分别在会理、会东、德源、盐源及冕宁5县选取烟草集中种植区样地进行调查,每点系统调查30株,试验地均不施任何农药。
1.2 聚集度指标测定
参照文献[10-14]。丛生指标:I=S2/m−1[式中S2为方差,m为烟蚜的种群平均密度(头/株)]。负二项分布指标:K=m2/(S2-m)。久野指数:CA=(S2-m)/m2。平均拥挤度指标:m*=m+(S2-m)/m。扩散系数:C=S2/m。田间个体平均大小测定:当种群属于负二项分布时,用L*指数表示其个体群的平均大小。用L*/(m+1)表示扩散型。
1.3 空间分布型格局分析
本试验运用Iwaom*-m回归分析法(m*=α+βm)和Taylor幂法则(lgS2=lg(a+b)×lgm或S2=a×mb)进行空间分布型格局分析[15-16]。
1.3.1 烟蚜聚集成因分析 运用种群聚集均数(λ)分析烟蚜的聚集原因,λ=mν/2k,式中m为烟蚜的平均值(头/株),k为负二项分布中的值,ν为自由度等于2k时的值,但由于2k常为小数,因此,值可用比例内插法求得。当λ<2时,昆虫聚集原因主要是由于某些环境因素所致;当λ≥2时,昆虫聚集原因主要是昆虫本身的聚集习性或与环境因素综合作用所致[17]。
1.3.2 种群密度与聚集度指标的关系 作为一个合理的扩散型指标在理论上应与种群平均密度m相互独立,这样的指标才适合各种密度水平的聚集程度变化的比较与分析,因此,以各种指标为变量,平均密度为自变量,计算各种聚集指标与平均密度的相关系数r[8]。
1.4 理论抽样数模型
1.4.1 Southwood的Kc法为理论抽样数;t为一定置信度下t分布值;D为允许误差;KC为负二项分布公共K值。
1.5 序贯抽样模型
1.5.1 Iwao序贯抽样技术

式中,T''n为累计虫数上限;T'n为累计虫数下限;T
n为累计虫数;mc为防治指标;t为对应于某一概率水平下的正态误差;α、β为m*-m回归式的截距和斜率;n为抽样数。若Tn≥T''n,则实际种群密度高于防治指标;若Tn≤T''n,则实际种群密度低于防治指标;若,则不能确定实际种群密度大小,还应继续抽样。
1.5.2 kuno序贯抽样技术

式中,Tn为停止抽样累计虫数;D0为精确度,α、β同上。当田间调查的累计虫量达到或超过所对应的抽样数量下的预定截止线时,便停止抽样,此时便可求得在抽样精度为D0下的田间烟蚜种群密度为Tn/n[18]。
1.6 数据分析
数据及图表均用Excel 2007及SPSS 16.0处理。
2 结果
2.1 主要试验地概况
本试验分别在凉山州会理、会东、冕宁、德昌及盐源县的南阁、姜州、回坪、德州及梅雨进行(表1),海拔最低为德昌县德州1400 m,最高为盐源县梅雨2330 m,种植的烟草品种主要包括红花大金元(红大)、云烟85及云烟87,另外,本试验调查面积均不低于800 m2。

表1 试验地相关参数Table 1 Related parameters about the test regions
2.2 烟蚜在主要烟区的聚集度指标测定
如表2所示,通过对烟蚜调查数据统计分析得到其在主要烟区的聚集度指标,其丛生指标(I)和久野指数(CA)均大于0,负二项指标(K)大于0且小于8,扩散系数(C)、平均拥挤度(m*/m)及L*/(m+1)均大于1,故烟蚜在烟田的主要分布型为聚集分布。
2.3 烟蚜聚集成因分析
由表3可知,在烟蚜危害高峰期,聚集原因主要是其本身的聚集习性或与环境因素综合作用所致(λ≥2);对聚集均数(λ)与种群平均密度(m)进行线性回归得λ=0.310+0.408m图1),依据1.3.1的烟蚜聚集成因分析,当λ=2时代入

表2 烟蚜在主要烟区的聚集度指标测定值Table 2 Indexs of aggregation of Myzus persicae on tobacco regions

表3 烟蚜聚集均数的确定Table 3 Aggregation average(λ)of Myzus persicae

图1 烟蚜λ-m的关系Fig.1 Relationship between aggregated mean and mean density
公式λ=0.310+0.408m得到m=4.14。因此,当烟蚜种群密度m≥4.14时,烟蚜种群的聚集为其本身的聚集习性或与环境因素综合作用所致,当m<4.14时,烟蚜种群聚集主要是由于某些环境因素所致。
2.4 种群密度与聚集度指标的关系
由表4表明,只有平均拥挤度(m*/m)、负二项分布指标(K),久野指数(CA)及L*/(m+1)与烟蚜种群平均密度(m)相关性不显著,其余与平均密度(m)相关性显著或极显著。所以,用平均拥挤度(m*/m)、负二项分布指标(K),久野指数(CA)及L*/(m+1)分析烟蚜空间分布型更具说服力。
2.5 空间分布型格局分析
2.5.1 Iwaom*-m回归 根据表2中m*-m值作回归分析得烟蚜的回归方程为m*=0.591+3.581m由此可知α=0.591>0,说明个体间相互吸引,β=3.591>1说明烟蚜的空间分布是聚集的(图2)。
2.5.2 Taylor幂法则 根据表2中S2-m值作Taylor幂函数得m1.083,由此可知a=1.449>0,b=1.083>1,说明烟蚜种群为聚集分布,聚集强度随种群密度的升高而增加;此外,利用负二项分布理论曲线公式:S2=m+Kc×m2结合实测的S2和m得到较好的吻合曲线,说明烟蚜在烟田为负二项分布(图 3)。

表4 种群平均密度与各聚集度指标之间的相关性Table 4 The correlation between average(m)and each aggregation index
2.6 理论抽样模型
2.6.1 Southwood的Kc法 由图2结合Kc计算公式可知Kc=0.1224,则有模型公式取D=0.1、0.2、0.3,t=1、1.64、1.96。
2.6.2 Iwao回归法 通过Iwao回归得α=0.591,β=3.581,则有模型公式D=0.1、0.2、0.3,t=1、1.64、1.96。
通过Southwood的Kc法,Iwao回归法及Taylor幂法理论抽样模型即可求得烟蚜在不同密度与精度要求下抽取的最合理理论抽样数n,且n随平均密度(m)、允许误差(D)及置信度(t)的变化而变化(表 5)。

图2 烟蚜m*-m关系Fig.2 Relationship between mean crowding and mean density
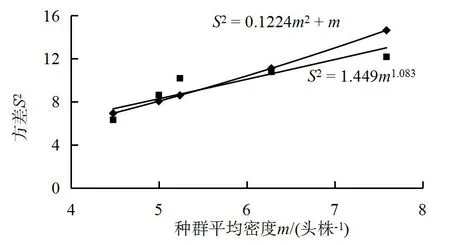
图3 烟蚜S2-m关系Fig.3 Relationship between variance and mean density

表5 烟蚜种群理论抽样数Table 5 Theoretical sample number for Myzus persicae
2.7 序贯抽样模型
2.7.1 根据Iwao序贯抽样技术 取t=1.64、1.96,防治指标mc=10头/株,15头/株及20头/株,则有公式:

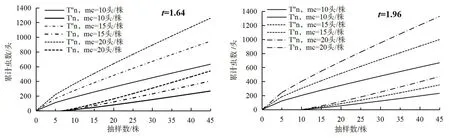
根据以上公式可得到烟蚜种群序贯抽样图,以t=1.64,mc=10头/株为例,若烟蚜调查累计虫数Tn>T''n,则应予以防治;若T'n 图4 烟蚜种群Iwao序贯抽样图Fig.4 Lwao’s sequential sampling 以D0=0.3为例,随机抽取30株后,当烟蚜累计虫量达到或超过401头时,则应停止抽样,而此时烟蚜种群密度为13.37头/株(图5)。 图5 烟蚜种群kuno序贯抽样图Fig.5 Kuno’s sequential sampling 种群是由个体组成的,种群内个体间的组合有一定的规律,从而使得种群动态既表现为数量的增减,也表现为空间的扩张和缩减,但某一时刻的种群空间分布型是一定的,是由种群个体本身的生物学特性以及环境因素共同决定的[19]。因此,研究昆虫空间格局不仅可以揭示种群内和种群间的空间结构,还可以为抽样方法和制定害虫防治指标提供依据。丛生指标(I)、聚集度指标(m*/m)、扩散系数(C)、久野指标(CA)等被用来研究昆虫种群的空间格局,但这些指标并不是研究同一昆虫种群空间格局的不同方法,而是反映空间格局的不同面,因此研究人员通过将众多指标综合分析种群空间格局[20-23],本研究发现无论采用何种聚集度指标,烟蚜种群均为聚集分布;此外,我国研究者提出了新的空间分布型研究方法并做了进一步优化,兰新平[24]提出了m*/v指标并在昆虫种群空间分布型研究中得到了验证,徐如梅等[25-26]对Iwao的m*-m回归模型做了改进并研究了温室白粉虱的空间分布型,本烟碱采用Iwao的m*-m回归模型研究表明,烟蚜种群呈聚集分布,且个体间相互吸引,这一结论与众多研究成果一致[5-9];本试验通过Taylor幂法则研究发现,存在参数b约为1的情况,因此通过马占山等[27]关于Taylor幂法则的结论解释烟蚜空间分布型更切合实际;通过种群聚集均数(λ)分析昆虫的聚集原因已被广泛利用[7-9,17],本研究通过聚集均数(λ)与种群密度(m)线性回归发现当烟蚜种群密度m≥4.14时,烟蚜种群的聚集为其本身的聚集习性或与环境因素综合作用所致,当m<4.14时,烟蚜种群聚集原因主要是由于某些环境因素所致,烟蚜种群呈聚集分布发生,这为利用天敌防治烟蚜提供了便利,但对天敌释放的时间,密度及位置等重要参数还应做进一步研究。 烟蚜种群呈聚集分布,聚集强度随种群密度的升高而增加,且聚集原因由烟蚜习性与环境因素共同决定。序贯抽样方法可以为害虫的抽样调查及防治提供理论依据,本研究结合Iwao及Kuno序贯抽样明确了在不同精度下烟蚜的科学抽样次数及防治参考线,在实际生产中可作为烟蚜调查与防治的参考依据。 [1]杨效文,张孝羲.我国不同地区烟蚜种群生殖特征研究[J].生态学报,2000,20(1):140-144. [2]何应琴,吴佳星,唐元满,等.黄瓜花叶病毒和马铃薯Y病毒混合侵染烟株对烟蚜取食行为的影响[J].植物保护学报,2017,44(1):32-38. [3]程新胜,韩移旺.不同体色烟蚜的消长动态及对药剂的敏感性[J].中国烟草科学,1998,18(3):33-34. [4]李勇,何林.不同虫态烟蚜对杀虫剂敏感性及其与解毒酶活力关系[J].环境昆虫学报,2015,38(2):408-413. [5]丁岩钦.昆虫数学生态学[M].北京:科学出版社,1994. [6]夏鹏亮,王瑞,王昌军,等.恩施烟区无翅桃蚜在烤烟田空间动态的地统计学分析[J].生态学报,2014,35(5):1198-1204. [7]陈斌,李正跃,孙跃先,等.烟田烟蚜种群数量时空动态研究[J].云南农业大学学报,2000,15(4):311-316. [8]黄本荣.三明烟区烟蚜空间分布型的研究[D].长沙:湖南农业大学,2009:1-20. [9]巫厚长,徐光曙,房明慧,等.烟蚜及其捕食性天敌草间小黑蛛种群空间结构分析[J].应用生态学报,2004,15(6):1039-1042. [10]曾维爱,李密,谭琳,等.长沙烟区天敌昆虫物种多样性及主要害虫的消长动态[J].中国烟草科学,2016,37(5):63-67. [11]CASSIE R M.Frequency distribution modes in the ecology of plankton and other organisms[J].Journal Animal Ecology,1962,31:65-92. [12]DAVID F N,MOORE P G.Notes on contagious distribution in plant populations.Annals of Botany,1954,18:47-53. [13]LLOYD M.Mean crowding[J].Journal Animal Ecology,1967,36:1-30. [14]常亚文,何娅婷,沈媛,等.三叶斑潜蝇幼虫种群空间格局及抽样技术研究[J].环境昆虫学报,2016,38(4):866-873. [15]IWAO S.A new regress method for analyzing the aggregation patteren of animal populations[J].Researches on Population Ecology,1968,10(1):1-20. [16]TALOR L R.Aggregation,variance and the mean[J].Nature,1961,189:732-735. [17]BLACKITH R E. Nearest neighbour distance measurements for theestimation of animal populations[J].Ecology,1958,39:147-150. [18]唐雅文,李艺琼,金启安,等.椰子木蛾幼虫在椰子树上的空间分布型及抽样技术[J].植物保护,2017,43(2):107-111. [19]戈峰.昆虫生态学原理与方法[M].北京:高等教育出版社,2008. [20]洪波,张云慧,李超,等.马铃薯甲虫空间分布型及序贯抽样[J].植物保护学报,2010(3):206-210. [21]白树雄,张聪,闫占峰,等.玉米田蚜虫种群的空间动态[J].应用昆虫学报,2014,51(3):661-667. [22]张应,崔广林,李隆云.胡萝卜微管蚜在灰毡毛忍冬上的空间分布格局[J].植物保护,2012,38(6):77-79. [23]胡懿君,赵映书,陈龙,等.宁夏中宁县麦蚜种群空间分布与蚜茧蜂的跟踪作用[J].华中农业大学学报,2016,35(3):30-37. [24]兰新平.松毒蛾越冬蛹种群密度简易序贯抽样方法[J].应用昆虫学报,1993,30(4):234-236. [25]徐汝梅,刘来福,丁岩钦.改进的IWAO M*-M模型[J].生态学报,1984,4(2):111-117. [26]徐汝梅,李兆华,李祖荫,等.温室白粉虱成虫空间分布型的研究[J].昆虫学报,1980,23(3):265-275. [27]马占山,张执中.油松毛虫幼虫和蛹在油松各轮枝层分布的空间格局及其变化规律研究[J].林业科学,1990,26(6):519-526.

3 讨论
4 结论
猜你喜欢
玩具世界(2022年3期)2022-09-20
中学生数理化(高中版.高二数学)(2022年5期)2022-06-01
中学生数理化(高中版.高考数学)(2021年3期)2021-06-09
中学生数理化(高中版.高考数学)(2021年3期)2021-06-09
中学生数理化·高三版(2021年3期)2021-05-14
特产研究(2021年2期)2021-04-13
黑龙江水利科技(2020年8期)2021-01-21
甘肃教育(2020年2期)2020-09-11
小学生必读(低年级版)(2018年11期)2018-03-13
现代农业科技(2017年1期)2017-03-06
