
2个紫花苜蓿品种叶片对氮的光合响应
2018-01-16 07:44:20朱天琦刘晓静
草原与草坪 2017年6期
朱天琦,刘晓静,郝 凤
(甘肃农业大学 草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
刘晓静为通讯作者。
氮是作物体内叶绿素的重要组成元素之一[1],直接影响着作物的光合作用[2]。叶片是植物进行光合作用最主要器官,并且叶面积大小、叶绿素含量是决定植物能否将光能充分转化为化学能的重要因素,也是植物光合作用能力的主要限制因素[3-4]。据报道,植物叶片75%的氮素存在于叶绿体中,施氮会极显著的增加作物幼苗叶绿素的含量,并能显著提高其净光合速率、气孔导度[5-6]。在作物生长初期,真叶生长发育尚未健全,子叶光合产物难以满足其生长发育的迫切需求,在此时,及时补充外源氮素来缓解其前期营养供给,会直接影响作物的产量与品质[7]。在自然环境下,可供作物吸收的氮素形态有2种,不同的氮素形态对作物光合速率的影响不尽相同。研究表明,小麦对NH4+-N的吸收速率显著高于NO3--N,且NH4+-N处理较NO3--N处理显著提高了小麦的氮积累与光合速率[8]。
紫花苜蓿(Medicagosativa)因是畜牧业重要的植物性蛋白资源而被广泛推广种植。由于其对氮素独特的利用方式,可以通过生物固氮来补充体内氮营养缺乏,但高效生产仍需要配合氮肥,且苗期施氮可以促进其根系生长,为根瘤菌侵染和结瘤创造较好的条件[9]。由此可见,氮素对紫花苜蓿早期生长具有重要意义,但对其生长早期光合特性影响的研究报道较少。由于甘农3号紫花苜蓿抗旱、耐寒、抗冻害能力强,春季返青较早,再生能力强,广泛种植于西北内陆灌溉农业区和黄土高原等地区,陇东苜蓿主要分布于甘肃庆阳、平凉等陇东地区,是这些地区古老的地方品种,由于其抗旱性较强,在旱作条件下,被广泛种植。因此,选择甘农3号和陇东苜蓿,通过对比分析不同外源氮素形态对不同品种紫花苜蓿光合特性的影响,以期从光合特性的角度探究氮素促进紫花苜蓿高效生产的生理基础,为紫花苜蓿氮营养管理提供参考。
1 材料和方法
1.1 试验材料
供试紫花苜蓿品种甘农3号(Medicagosativacv.Gannong No.3), 陇东苜蓿(Medicagosativacv.Longdong),中华根瘤菌(12531),由甘肃农业大学草业学院提供。
选用高15 cm、长20 cm、宽12 cm的花盆,装入3 kg用蒸馏水清洗并灭菌的石英砂。
1.2 试验设计
设2种氮素形态(NH4+-N、NO3--N),5个浓度水平(0、105、210、315、420 mg/L),共9个处理,分别以CK(氮素水平为0)、NO3--105、NO3--210、NO3--315、NO3--420、NH4+-105、NH4+-210、NH4+-315、NH4+-420表示,每处理重复6次,共54盆,完全随机排列。试验在室外防雨网室内培养,采用营养液砂培法,以Fahraeus无氮营养液[10](Na2HPO4·12H2O 0.15 g,MgSO4·7H2O 0.12 g,EDTA-Fe 0.007 5 g,CaCl2·2H2O 0.1 g,KH2PO40.1 g,Gibson 微量元素1 mL,H2O 1 000 mL,pH 6.5~7.0)为基本营养液,以Ca(NO3)2和(NH4)2SO4为NO3--N、NH4+-N来源,按不同水平配置营养液。
2016年4月11日播种,每盆播种50粒灭菌种子,播种7 d后间苗,每盆留苗30株,然后开始浇入营养液,每周浇250 mL,为防止砂培中盐分积累,每7 d用蒸馏水淋洗1次,并浇入新配置营养液。待紫花苜蓿第1片真叶长出,每盆接种新培养的根瘤菌液25 mL,培养至40 d后取样测定各项指标。
1.3 测定指标与方法
采用GFS-3000便携式光合系统分析仪(Walz,Germany),于晴天上午9∶00~11∶00测定叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)。
叶面积:采用台式扫描仪进行扫描,通过分析软件对叶片的图像进行分析,每个处理重复6次。
叶绿素含量:采用丙酮浸提法[11]。称取新鲜叶片0.2 g,放入具塞试管中,加入乙醇-丙酮等体积混合液10 mL,置于培养箱中(无光),至叶片完全变白(以空白提取液为对照),测定D649 nm,D665 nm。
Ca(mg/g FW)=13.95D665 nm-6.88D649 nm
Cb(mg/g FW)=24.96D649 nm-7.32D665 nm
叶绿素总含量(mg/g FW)=色素水平×提取液体积×稀释倍数/样品鲜重
1.4 数据处理
采用Excel 2007软件进行数据的整理,应用SPSS 17.0对数据进行差异显著性分析。
2 结果与分析
2.1 不同品种紫花苜蓿叶面积对氮的响应
不同氮素处理下甘农3号和陇东苜蓿叶面积变化趋势一致,供氮处理均显著高于CK(P<0.05),随供氮水平的增加呈先增加后降低的趋势,并在210 mg/L水平时达到峰值(表1)。在210 mg/L水平下,NO3--N培养下甘农3号和陇东苜蓿分别比对照增加了32.67%、24.00%,NH4+-N培养下分别增加了38.61%、40.00%。相同氮素水平下,整体上表现为NH4+-N处理高于NO3--N处理。对比2种基因型紫花苜蓿可知 ,甘农3号对氮营养的响应程度高于陇东苜蓿,在NO3--210 mg/L浓度时, 甘农3号叶面积为1.34 cm2,与陇东苜蓿相比增加了8.1%。因此,NH4+-N 210 mg/L处理是紫花苜蓿叶片生长发育最适宜的氮素形态及水平。
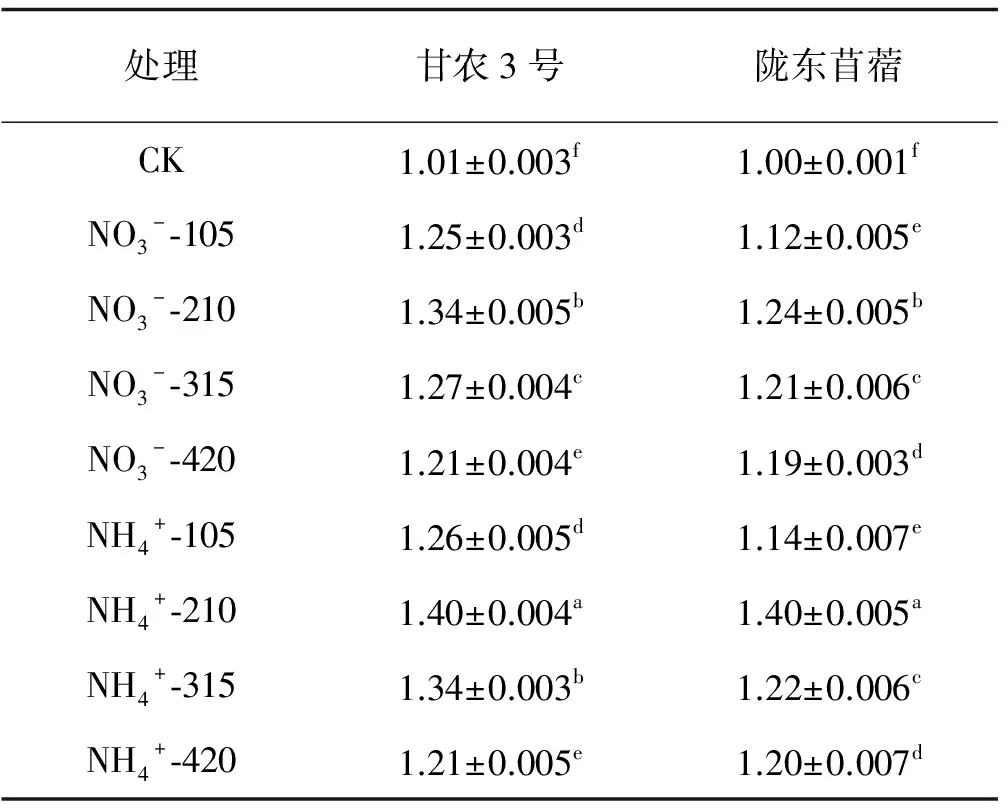
表1 不同品种紫花苜蓿叶面积对氮的响应
注:数据为平均值±标准误,同列不同小写字母表示差异显著(P<0.05),下同
2.2 不同品种紫花苜蓿幼苗叶绿素对氮的响应
不同氮素处理下甘农3号和陇东苜蓿的叶绿素含量变化趋势一致,供氮处理均显著高于CK(P<0.05),其含量均随供氮水平的增加呈先增加后降低的趋势,并表现在210 mg/L水平时达到峰值(表2)。在210 mg/L水平下,NO3--N培养下甘农3号和陇东苜蓿比对照分别增加了50.56%、73.33%,NH4+-N培养下分别增加了59.55%、84.00%。相同氮素水平下,整体上表现为NH4+-N处理高于NO3--N处理。对比2个紫花苜蓿品种可知,甘农3号对氮营养的响应程度高于陇东苜蓿,在210 mg/L浓度时,NO3--N培养下甘农3号叶绿素含量与陇东苜蓿相比增加了3.08%;NH4+-N培养下,甘农3号比陇东苜蓿增加了2.9%。因此,NH4+-N 210 mg/L处理是紫花苜蓿叶绿素合成适宜的氮素形态及水平。
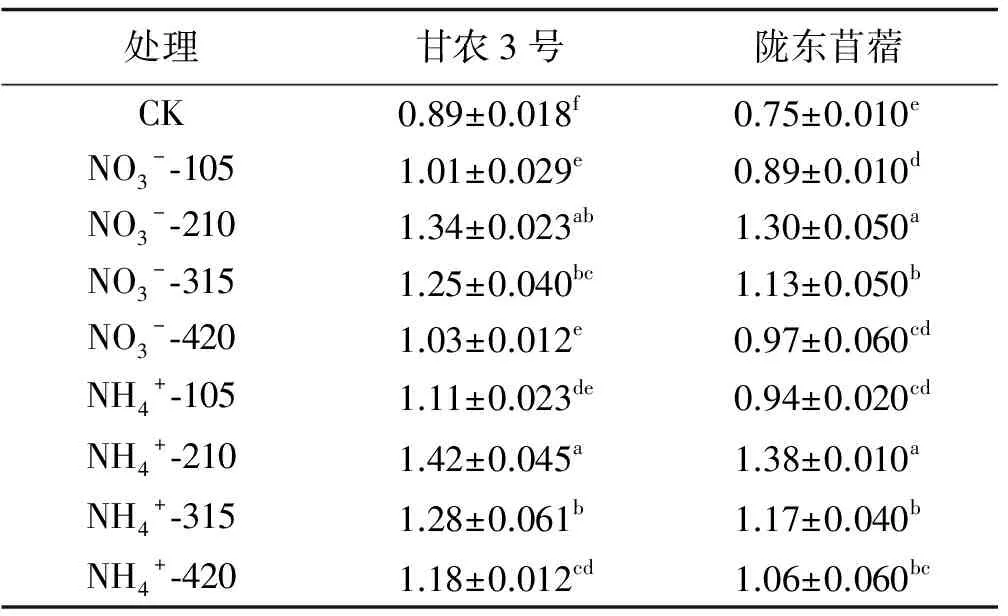
表2 不同品种紫花苜蓿叶绿素对氮的响应
2.3 不同品种紫花苜蓿幼苗光合速率对氮的响应
氮营养对紫花苜蓿幼叶净光合速率具有显著的调节作用。供氮处理均显著高于CK(P<0.05),随供氮水平的增加呈先增加后减小的趋势,均在210 mg/L处理水平下达到最大值(图1)。在210 mg/L处理水平下,NO3--N培养下甘农3号和陇东苜蓿净光合速率比对照分别增加了189.30%、210.10%,NH4+-N培养下分别增加了329.63%、250.65%。相同氮素水平下,整体上表现为NH4+-N处理高于NO3-N处理。对比2个紫花苜蓿品种可知,甘农3号对氮营养的响应程度明显高于陇东苜蓿,在210 mg/L浓度时,NO3--N培养下甘农3号净光合速率与陇东苜蓿相比增加了3.12%;NH4+-N培养下,甘农3号比陇东苜蓿增加了35.43%。因此,在NH4+-210处理下,幼叶净光合速率反应最为敏感。
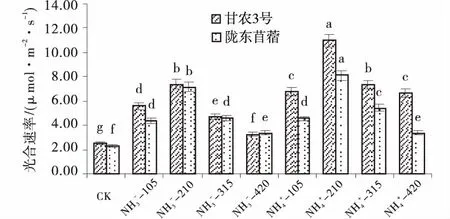
图1 不同品种紫花苜蓿光合速率对氮的响应Fig.1 Effects of nitrogen application on photosynthetic rate注:图中不同小写字母表示同一品种各水平之间差异显著(P<0.05)
2.4 不同品种紫花苜蓿幼苗蒸腾速率对氮的响应
氮营养调控下2个紫花苜蓿品种幼叶的蒸腾速率显著增加。供氮处理均显著高于CK(P<0.05),随供氮水平的增加呈先增加后减小的趋势,均在210 mg/L处理水平下达到最大值(图2)。在210 mg/L处理水平下,NO3--N培养下甘农3号和陇东苜蓿蒸腾速率比对照分别增加了51.6%、47.29%,NH4+-N培养下分别增加了88.06%、29.93%。相同氮素水平下,整体上表现为NH4+-N处理高于NO3--N处理。甘农3号对氮营养的敏感程度明显高于陇东苜蓿,在210 mg/L浓度时,NO3--N培养下甘农3号蒸腾速率与陇东苜蓿相比增加了16.25%;NH4+-N培养下,甘农3号比陇东苜蓿增加了27.68%。
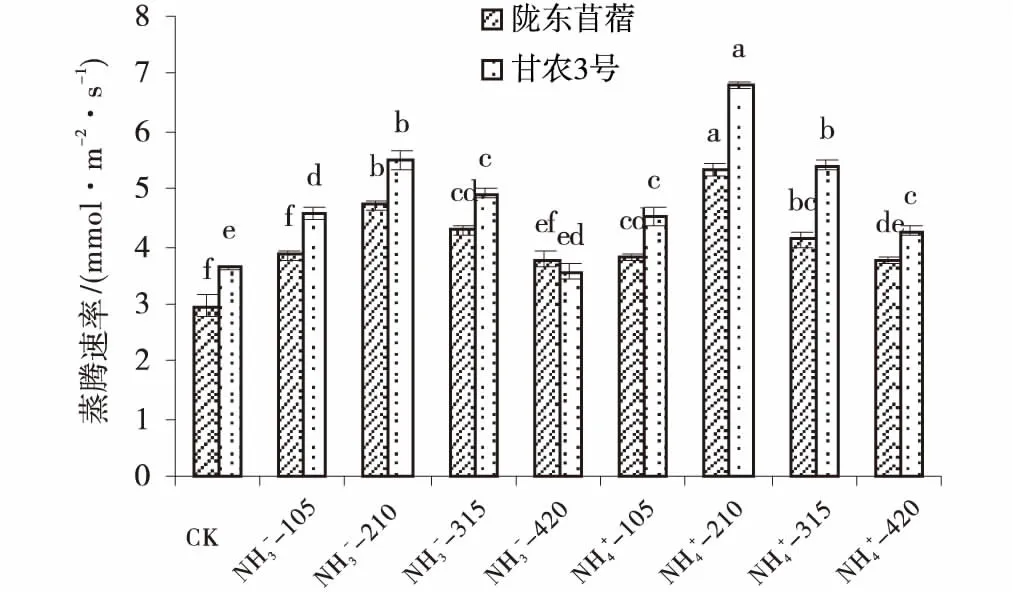
图2 不同品种紫花苜蓿蒸腾速率对氮的响应Fig.2 Effects of nitrogen application on transpiration rate注:图中不同小写字母表示同一品种各水平之间差异显著(P<0.05)
2.5 不同品种紫花苜蓿幼苗气孔导度对氮的响应
供氮处理的气孔导度均显著高于CK(P<0.05),并随供氮水平的增加呈先增加后减小的趋势,均在210 mg/L处理水平下达到最大值(图3)。在210 mg/L处理水平下,NO3--N培养下甘农3号和陇东苜蓿气孔导度比对照分别增加了86.3%、73.38%,NH4+-N培养下分别增加了227.03%、192.40%。相同氮素水平下,整体上表现为NH4+-N处理高于NO3--N处理。对比2个紫花苜蓿品种可知,甘农3号对氮营养的响应程度明显高于陇东苜蓿,在210 mg/L浓度时,NO3--N培养下甘农3号气孔导度比陇东苜蓿增加了10.3%;NH4+-N培养下,甘农3号比陇东苜蓿增加了14.82%。因此,NH4+-210 mg/L处理最有利于紫花苜蓿叶片气孔导度增大,从而提高植株体内与外界气体交换的通量。
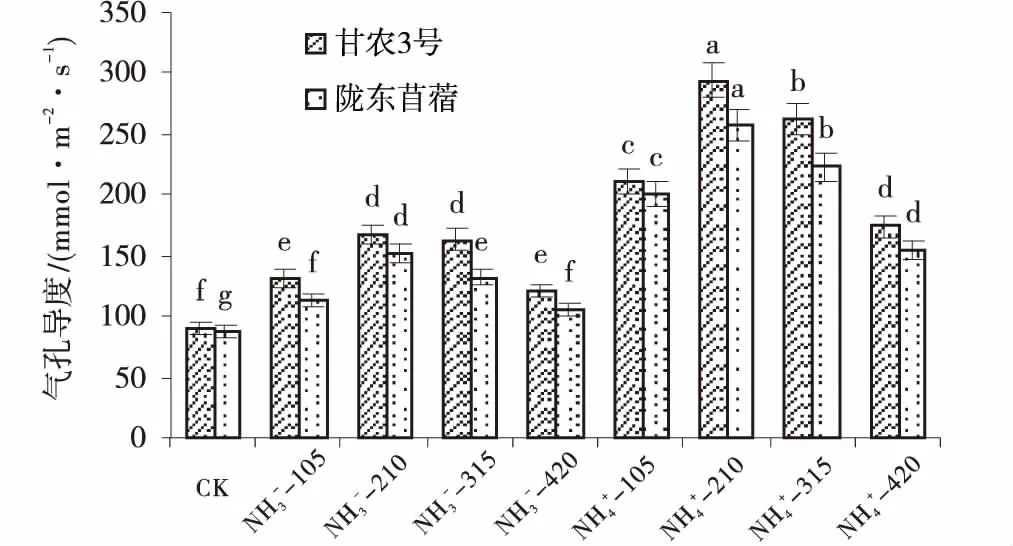
图3 不同品种紫花苜蓿气孔导度对氮的响应Fig.3 Effects of nitrogen application on stomatal conductance注:图中不同小写字母表示各水平之间差异显著(P<0.05)
2.6 不同品种紫花苜蓿幼苗胞间CO2浓度对氮的响应
供氮处理的胞间CO2浓度比CK显著减小(P<0.05),随供氮水平的增加呈现递减趋势。同一氮素水平下,整体上表现为NH4+-N处理下降的幅度显著大于NO3--N处理(P<0.05)(图4)。对比2种基因型紫花苜蓿可知,相同氮营养调控下陇东苜蓿胞间CO2浓度高于甘农3号。
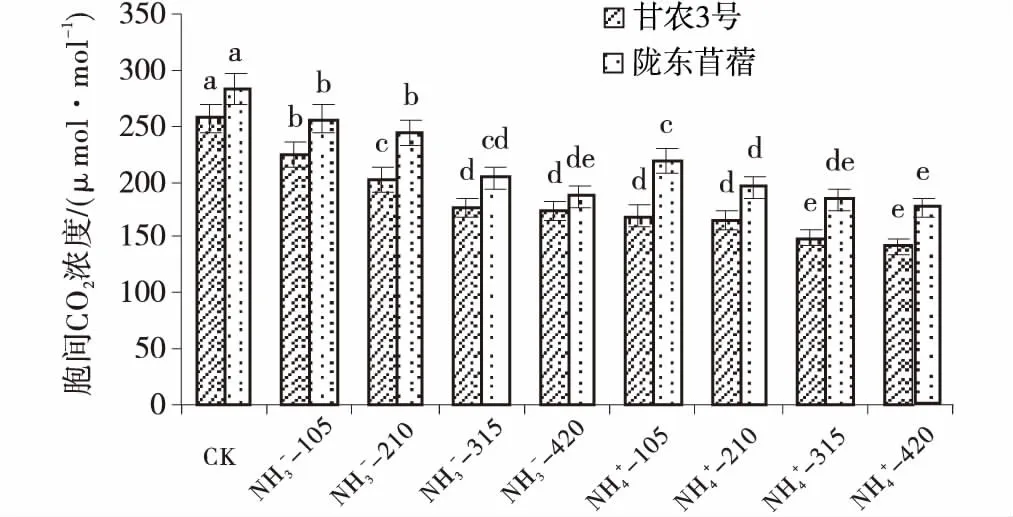
图4 不同品种紫花苜蓿胞间CO2浓度对氮的响应Fig.4 Effects of nitrogen application on intercellular CO2 concentration注:图中不同小写字母表示同一品种各水平之间差异显著(P<0.05)
3 讨论
光合能力是作物品质与产量形成的根本保证,叶面积、叶绿素含量是影响其光合能力的重要因素[12]。张生杰等[13]在氮素对不同品种烤烟叶片光合特性及产量品质的研究中发现,烤烟叶片叶绿素随施氮量的增加而升高,并可以显著的缓解烤烟叶片叶绿素的降解,增强叶片光合能力。此次研究表明,施氮能显著促进2个紫花苜蓿品种叶面积及叶绿素含量(P<0.05),并且两者均在NH4+-210 mg/L处理达到最大值,NH4+-N和NO3--N培养下叶面积与叶绿素均呈现先增加后降低的趋势,说明氮素添加能有效促进紫花苜蓿叶的生长发育,并促进叶绿素的合成,氮浓度过高与过低均不利于叶的生长发育。在同一氮素水平下,NH4+-N培养要好于NO3--N,究其原因可能是因为NH4+-N可显著提高紫花苜蓿体内Fe的积累[14](P<0.05),Fe是叶绿素合成的必须因子和铁氧蛋白的组分,Fe的大量积累,有利于植株铁氧蛋白的合成,继而稳定了叶绿体的结构,促进了叶绿素的合成[15]。这一点也在叶莉莎等[16]氮素形态及配比对雷竹光合特性的研究中得到了证实。2个不同品种紫花苜蓿相比,相同氮水平处理下甘农3号叶面积大小、叶绿素含量均高于陇东苜蓿,说明甘农3号叶的生长发育对氮素较为敏感,对氮素的转化及利用率较陇东苜蓿高。
罗雪华等[17]在氮水平及形态对巴西橡胶树花药苗生长的研究中发现,随着NH4+-N含量的增加,巴西橡胶树幼苗净光合速率均呈现先增加后减小的趋势。研究发现,供氮处理均显著高于CK(P<0.05),随供氮水平的增加呈先增后减的变化趋势,并均在210 mg/L处理水平下达到最大值。说明氮营养对紫花苜蓿幼叶净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)具有显著的调节作用,在一定的范围中增加氮营养有利于叶片净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)的增加。胞间CO2浓度(Ci)的影响与光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)变化相反,呈现出递减趋势。Ci的大小决定于叶片周围空气CO2的浓度、气孔导度、叶肉导度和叶肉细胞的光合活性4种因素,其与Pn、Tr、Gs表现出相反的趋势,究其原因可能是因为光合速率随着施氮处理后光合色素增加,光合有效面积增大,继而使得叶肉细胞的CO2羧化能力增加,胞间CO2急剧消耗导致其浓度降低,从而表现出与光合速率相反的趋势。胡海非等[18]在油麦菜幼苗的研究上也得出了相同的结论。在同一氮素水平下,NH4+-N培养要好于NO3--N,可能因为NH4+-N提高了水的光解和电子传递的速率,提高了希尔反应活性及光合磷酸化能力,从而提高了植株的光合能力[19]。Raab等[20]在甜菜的研究中也证实了这一点。2个不同紫花苜蓿品种相比,甘农3号净光合速率、蒸腾速率、气孔导度均高于陇东苜蓿,胞间CO2浓度表现相反,说明甘农3号对氮营养响应程度及光合能力较陇东苜蓿更为敏感。陈水红等[21]和刘玉华等[22]的研究也表明,不同紫花苜蓿品种光合特性存在显著差异,光合性能较高的品种对营养物质的吸收及利用更为敏感。
4 结论
施氮能显著增大紫花苜蓿叶面积、增加叶绿素的含量,提高其光合性能;2种氮素形态在同一氮水平下,NH4+-N更有利于植株光合性能的提高,且均在210 mg/L水平时达到峰值;甘农3号的叶面积、叶绿素及光合性能对氮的响应程度及氮的转化与利用均高于陇东苜蓿。
[1] 曹翠玲,李生秀.氮素形态对小麦中后期的生理效应[J].作物学报,2003,29(2):258-262.
[2] 罗雪华,邹碧霞,吴菊群,等.氮水平和形态配比对巴西橡胶树花药苗生长及氮代谢、光合作用的影响[J].植物营养与肥料学报,2011,17(3):693-701.
[3] 赵长星,马东辉,王月福,等.施氮量和花后土壤含水量对小麦旗叶衰老及粒重的影响[J].应用生态学报,2008,19(11):2388-2393.
[4] 马富举,李丹丹,蔡剑,等.干旱胁迫对小麦幼苗根系生长和叶片光合作用的影响[J].应用生态学报,2012,23(3):724-730.
[5] 吴楚,王政权,范志强,等.不同氮浓度和形态比例对水曲柳幼苗叶绿素合成、光合作用以及生物量分配的影响[J].植物生态学报,2003,27(6):771-779.
[6] 陈静,刘连涛,孙红春,等.氮素水平对棉花幼苗生长和光合特性的影响[J].棉花学报,2013,25(5):403-409.
[7] 孙进武,刘金祥,陈三有,等.不同构树光合作用的光响应比较研究[J].草原与草坪,2010,30(3):47-51.
[8] 戴廷波,曹卫星,荆奇.氮形态对不同小麦基因型氮素吸收和光合作用的影响[J].应用生态学报,2001,12(6):849-852.
[9] 蒯佳林,刘晓静,李文卿.不同氮素水平对接种根瘤菌紫花苜蓿生长特性的影响[J].草原与草坪,2011,31(3):56-59.
[10] 孙建光,张燕春,徐晶,等.高效固氮芽孢杆菌筛选及其生物学特性[J].中国农业科学,2009,42(6):20-45.
[11] 王磊,白由路.不同氮处理春玉米叶片光谱反射率与叶片全氮和叶绿素含量的相关研究[J].中国农业科学,2005,38(11):2269-2270.
[12] 段巍巍,赵红梅,郭程瑾,等.夏玉米光合特性对氮素用量的反应[J].作物学报,2007,33(6):949-954.
[13] 张生杰,黄元炯,任庆成,等.氮素对不同品种烤烟叶片衰老、光合特性及产量和品质的影响[J].应用生态学报,2010,21(3):668-674.
[14] 于铁峰,刘晓静,郝凤,等.NO3--N/NH4+-N配比对紫花苜蓿营养品质及饲用价值的影响研究[J].草业学报,2016,25(12):102-110.
[15] 李唯.植物生理学[M].北京: 高等教育出版社,2012:48-75.
[16] 叶莉莎,陈双林.氮素形态及配比对雷竹光合特性和氮代谢酶活性的影响[J].生态学杂志,2016,35(9):2355-2360.
[17] 罗雪华,邹碧霞,吴菊群,等.氮水平和形态配比对巴西橡胶树花药苗生长及氮代谢、光合作用的影响[J].植物营养与肥料学报,2011,17(3):693-701.
[18] 胡海非,张巧柔,吴卫东,等.氮素形态对油麦菜幼苗光合作用及其荧光特性的影响[J].热带作物学报,2016,37(1):7-14.
[19] 郭培国,陈建军,郑燕玲.氮素形态对烤烟光合特性影响的研究[J].植物学报,1999,16(3):262-267.
[20] Raab K T,Terry N.Nitrogen source regulation of growth and photosynthesis in Beta vulgaris[J].Plant Physiology,1994,105:1159-1166.
[21] 陈水红,高军,张剑云,等.不同紫花苜蓿品种光合性能比较研究[J].湖北农业科学,2009,48(12):3100-3102.
[22] 刘玉华,贾志宽,史纪安,等.旱作条件下不同苜蓿品种光合作用的日变化[J].生态学报,2006,26(5):1468-1477.
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
机械研究与应用(2022年3期)2022-07-25 05:52:40
中国饲料(2021年17期)2021-11-02 08:15:24
现代畜牧科技(2021年9期)2021-10-13 06:38:52
现代畜牧科技(2021年4期)2021-07-21 06:13:04
生态学报(2021年6期)2021-04-25 08:24:42
农业工程学报(2021年3期)2021-04-15 10:03:14
节水灌溉(2020年9期)2020-09-16 00:32:12
现代园艺(2018年3期)2018-02-10 05:18:23
新商务周刊(2017年6期)2017-10-17 05:01:31
