
温盐交互胁迫下蒙古沙冬青生长及生理响应特征1)
2018-01-15 06:02李亦超秦玉梅韩蕊刘春艳郑天宇石莎
东北林业大学学报 2018年1期
李亦超 秦玉梅 韩蕊 刘春艳 郑天宇 石莎
(中央民族大学,北京,100081)
作为第三纪的遗迹,蒙古沙冬青适应潮湿和温暖的气候,其极端的性能与耐逆境胁迫的后续演化可以归因于渐进的气候变化和地质变化[1],因此,鲜明的特征使其成为研究逆境胁迫机理演变的很有价值的系统。
蒙古沙冬青因其超强的抗逆性而受生物学家的长期研究关注。从基因的克隆和转化[2-8]、鉴定及功能分析,蛋白的研究[9-12],到种子萌发抗逆性[13-16]的研究,再到对幼苗结构特性、抗逆生理、调节物质[17-18]的研究,种种方面均有涉及。在对于蒙古沙冬青幼苗的抗性研究方面,大多数文献体现的是用分子手段在分子层面进行研究,在宏观层面直接测定生理生态指标参数的试验往往只研究某一种胁迫的其中某些参数,由于蒙古沙冬青根系发达、根部具有根瘤和外观形态的其他特征[19]而适宜在沙砾质土壤或薄层覆沙的砾石质土壤地生长,因此,当前对于蒙古沙冬青抗旱性领域的研究较多,植株在适应气候变化过程中形成了明显的旱生结构,栅栏组织发达、海绵组织退化、气孔小而单位面积气孔数目多、有表皮毛、气孔高度下凹等特点[20-21]是其耐干旱的主要原因,除了超旱生结构,蒙古沙冬青体内的可溶性糖、游离氨基酸、保护酶等系统在抗旱方面也发挥了一定的作用。
随着研究的深入,蒙古沙冬青在高温和高盐胁迫方面也表现出了较强的抗性,并且当前盐渍化土地问题日益严重,因此,对于蒙古沙冬青在逆境胁迫,特别是多种逆境交互胁迫下生理生态响应特征的研究显得格外重要,目前,在幼苗抗性领域所取得主要研究进展包括,在高温胁迫方面,由于蒙古沙冬青本身就生长在温度较高的地区,故其对高温环境已经产生了良好的生态适应性,一般认为外界环境超过40 ℃时其光合反应中心的活性就会受到影响,但恢复到正常温度后蒙古沙冬青仍能正常生长[22],而超过50 ℃植株会出现萎蔫状况,无法正常生长;在盐胁迫方面,蒙古沙冬青幼苗对单盐胁迫具有很强的抗性[23],可以耐受的最高盐渍度约为1.3%,但对于两种及以上复盐同时存在的胁迫抗性较差,一般在0.6%左右就已达到其耐受限度,并且其体内可溶性糖和脯氨酸质量分数也会随着盐质量分数的升高而发生变化以适应盐胁迫。本试验通过分析温盐交互胁迫下蒙古沙冬青幼苗生理生态响应的差异与联系,为西北水土流失区提供防风固沙林树种和改造荒漠的优良灌木提供理论依据。
1 材料与方法
1.1 试验材料及胁迫条件设置
本试验所用种子取自内蒙古鄂尔多斯市鄂托克旗蒙西镇。萌发种子,集体栽培至长出3~4片真叶时,选取生长状况相同的幼苗分栽到花盆中用蛭石培养,每盆培养20株幼苗,每三盆进行相同处理。
温度胁迫处理方法:利用恒温培养箱模拟温度胁迫,在其他条件均相同情况下,分别在20 、30 、40 ℃ 3个温度下恒温处理30 d,各温度均为黑暗12 h,光照12 h(光照强度为6 400 lx)[24]。
盐胁迫处理方法:在其他条件均相同情况下,共设6个盐分处理。
处理1:空白处理,每隔2 d定量浇水,保持蒸发平衡和蛭石含水量稳定。
处理2:用盐质量分数为0.3%(指NaCl占土壤干质量的千分比)、V(盐)∶V(蛭石)=3∶1的溶液从幼苗基部蛭石层均匀浇灌,花盆下垫塑料小盘,目的是将浇水时流出的溶液及时倒回花盆内,避免盐分流失,每隔2 d定量浇水,保持蒸发平衡和蛭石含水量稳定[25],并且在盐胁迫结束后测定蛭石含盐量,盐胁迫处理为30 d。
处理3~处理6以此类推,盐质量分数分别为0.6%、0.9%、1.2%和1.5%。
待幼苗长到7~8片真叶时,按照上述温度与盐胁迫处理的方法进行交互胁迫处理,即将盐质量分数为0、0.3%、0.6%、0.9%、1.2%和1.5%的处理(每种处理有3个平行,共18盆),分别置于20 、30 、40 ℃ 3个温度下(一个温度下放置不同盐质量分数处理下的三盆)共54盆,各处理梯度均为预试验所得。
1.2 测量方法及数据分析
株高伸长量:利用直接测量法测植株株高,株高伸长量为受到胁迫后植株高度与胁迫前植株高度之差。
比叶面积:利用直接测量法测叶面积和叶片干质量,比叶面积为叶面积与干质量的比值。
光合速率:利用Li-6400光合仪测定光合速率,先将光合仪预热,待仪器稳定后再进行测定,测定相同部位的叶片,记录试验数据以及测定区域的叶面积。
叶绿素质量分数:丙酮法测定叶绿素质量分数,将所取叶片擦净称量后,剪碎加入1~2 mL 80%丙酮研磨后提取叶绿素,倒入刻度试管中,定容至10 mL,4 000转·min-1离心5 min,将色素提取液倒入比色杯中,用分光光度计在波长为652 nm(OD652)下测定吸光度值。
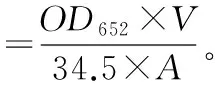
式中:V为提取液体积;A为叶片鲜质量。
叶片相对含水量:饱和称质量法计算叶片相对含水量,取鲜叶立刻称其鲜质量,后放入盛有蒸馏水的容器中浸泡24 h,取出后称饱和鲜质量,后在105 ℃下烘24 h,至恒质量后称取干质量。

水势:小叶流法测定水势,首先配制一系列不同浓度的蔗糖溶液,分别是 0.05、0.10、0.20、0.30、0.40、0.50、0.60、0.80 mol·L-1各10 mL,共8管。注入试管中,各管都塞上棉花并编号,作为空白对照组,另取8支试管,与前一组试管对应编号,作为试验组,然后由对照组各试管分别取溶液约4 mL,移入相同编号组的各试管中,再向试验组中投入同等数量的叶片加塞并经常摇动,30 min后,向试验组的每支试管中各加一滴甲烯兰溶液,摇动试管,使溶液均匀着色,用滴管(或小量程移液管)吸取浸有材料的蓝色溶液,然后插入对照组相同编号的试管,在液体的中部轻轻挤出一滴蓝色试验溶液,仔细观察液滴移动的方向,并记录。经过预试验后,蔗糖溶液质量分数梯度再逐渐缩小,以得到较精确结果,由标准压力下溶液的水势(渗透势的相反数)φs=-iCRT(式中:i为解离系数;C为摩尔浓度;R为气体常数(8.32 J·mol-1·K-1);T为热力学温度),计算可得水势。
可溶性糖质量分数:蒽酮比色法测定可溶性糖质量分数,将沙冬青叶片剪碎后加入蒸馏水沸水浴以提取可溶性糖,取适量可溶性糖提取液加入浓硫酸和蒽酮试剂后沸水浴,冷却后在630 nm处比色,记录光密度值。

脯氨酸质量分数:茚三酮比色法测定脯氨酸质量分数,3%的磺基水杨酸提取脯氨酸后加入冰醋酸和酸性茚三酮,再进行沸水浴,加入甲苯后3 000 r·s-1离心5 min,吸取上清液在波长520 nm处比色,获得吸光度值。
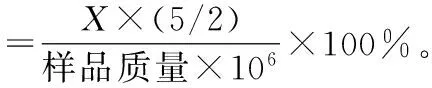
采用Excel 2003软件进行数据统计及图表绘制。SPSS 17.0软件对幼苗生长的各项生理生态指标进行单因素方差分析(One-way ANOVA)和Duncan多重比较,在置信水平95%采用LSD多重比较法检验其差异显著性[26],以平均值±标准差表示试验结果中所有数据。
2 结果与分析
2.1 温盐交互胁迫对蒙古沙冬青幼苗表观指标的影响
幼苗在经历30 d的交互胁迫后,幼苗死亡率均在正常范围内,可以进行正常生理生态指标的测定。
处理1到处理6分别代表盐胁迫质量分数从低到高(表1)。3个温度胁迫下,幼苗株高伸长量随盐质量分数的升高整体呈现下降趋势,且在处理2和处理4之间变化幅度不明显,在处理5和处理6下发生明显变化,在处理1下,40 ℃下幼苗株高伸长比30 ℃下伸长量少0.50 cm左右;在不同盐质量分数处理下,30 ℃ 胁迫时幼苗植株始终最高,40 ℃时其植株始终最矮,当温度在30 ℃时,处理5条件下幼苗株高伸长比处理1条件下伸长量少1.00 cm左右。与30 ℃处理1时相比,40 ℃处理3条件下其株高伸长量显著减小(P<0.05),在处理5和处理6条件下产生极显著变化(P<0.01)。由此可见,高温与盐交互胁迫显著抑制蒙古沙冬青幼苗的生长,且1.2%的盐质量分数相比于40 ℃的温度对蒙古沙冬青幼苗正常生长的抑制作用更明显。
2.2 温盐交互胁迫对蒙古沙冬青幼苗光合生理指标的影响
在不同质量分数盐胁迫处理条件下,蒙古沙冬青幼苗均在30 ℃下获得最适生长,光合生理生态指标达到最优,具体体现在比叶面积最小而光合速率及叶绿素质量分数最大,这说明在本次试验条件下30 ℃是蒙古沙冬青幼苗生长的最适温度,而过高和过低的温度均会对其各项光合生理指标造成不利影响,并且40 ℃相比于20 ℃对蒙古沙冬青幼苗光合生理指标的影响更大;在各温度处理下,比叶面积随着盐质量分数的升高而增大,与对照组相比,一般在处理4条件下达到显著差异(P<0.05),在处理5和处理6条件下达到极显著(P<0.01),而光合速率及叶绿素质量分数随着盐质量分数的升高而降低,且与对照组相比,在处理5条件下产生显著差异(P<0.05),在处理6条件下均产生极显著差异(P<0.01),这说明在本次试验条件下蒙古沙冬青幼苗光合生理指标在低盐质量分数中表现出较强的抗性,而高盐质量分数对其光合作用产生很大抑制。
在处理1条件下,40 ℃下幼苗比叶面积比30 ℃下比叶面积高230.000 0 cm2·g-1左右、光合速率低2.400 0 μmol·m-2·s-1左右、叶绿素质量分数少0.500 0 mg·g-1左右;当温度在30 ℃时,处理6条件下幼苗比叶面积比处理1条件下比叶面积高430.000 0 cm2·g-1左右、光合速率低3.200 0 μmol·m-2·s-1左右、叶绿素质量分数少1.000 0 mg·g-1左右。
2.3 温盐交互胁迫对蒙古沙冬青幼苗水分生理指标的影响
在不同质量分数盐胁迫处理条件下,蒙古沙冬青幼苗均在30 ℃下获得最适生长,水分生理生态指标达到最优,具体体现在叶片相对含水量和叶片水势最高而可溶性糖质量分数及脯氨酸质量分数最低,这再次说明在本次试验条件下30 ℃是蒙古沙冬青幼苗生长的最适温度,而过高和过低的温度均会对其各项水分生理指标造成不利影响,并且40 ℃相比于20 ℃对蒙古沙冬青幼苗水分生理指标的影响更大;在各温度处理下,叶片相对含水量和叶片水势随着盐质量分数的升高而降低,与对照组相比,一般在处理5条件下达到显著差异,在处理6条件下均达到极显著差异,而可溶性糖质量分数及脯氨酸质量分数随着盐质量分数的升高而增加,且与对照组相比,脯氨酸质量分数在处理5条件下产生显著差异,在处理6条件下均产生极显著差异,而可溶性糖质量分数无显著差异发生,这说明在本次试验条件下蒙古沙冬青幼苗水分生理指标在低盐质量分数中表现出较强的抗性,而高盐质量分数促进了其渗透调节作用的发生,并且渗透调节物质主要是脯氨酸。
在处理1条件下,40 ℃下幼苗叶片相对含水量比30 ℃下相对含水量少0.020 0%左右、水势低0.500 0 MPa左右、脯氨酸质量分数高75.000 0 mg·g-1左右;当温度在30 ℃时,处理6条件下叶片相对含水量比处理1条件下相对含水量少0.040 0%左右、水势低0.600 0 MPa左右、脯氨酸质量分数高125.000 0 mg·g-1左右。
2.4 温度、盐胁迫和温盐交互胁迫比较分析
温度胁迫和盐胁迫单一胁迫对蒙古沙冬青幼苗的株高、比叶面积、光合速率、叶绿素质量分数、叶片相对含水量、水势、可溶性糖质量分数及脯氨酸质量分数均有极显著影响(P<0.01),温度与盐的交互胁迫作用对蒙古沙冬青幼苗植株叶绿素质量分数仅有显著影响,而对可溶性糖质量分数并未出现影响,对其他生理生态特征则与单一胁迫相似,呈极显著影响。

表1 温度和盐交互胁迫下蒙古沙冬青幼苗的各项生理指标
注:表中数据为平均值±标准差;同列不同字母表示相同温度下不同质量分数盐处理间差异显著(P<0.05)。

表2 温度、盐与温盐交互作用对蒙古沙冬青幼苗生理生态特征的影响
注:表中数据为双因素方差分析的F值;*代表P<0.05,** 代表P<0.01。
3 结论与讨论
在预试验中,45 ℃下恒温处理30 d会使绝大部分幼苗萎蔫直至死亡,说明45 ℃恒温条件在本次试验中已超过蒙古沙冬青幼苗的耐受限度,由试验结果可知,在本次试验条件下,蒙古沙冬青幼苗的最适生长温度是30 ℃,低于或高于这一温度10 ℃接受处理的幼苗都会受到不同程度的胁迫,并在表观、生理和代谢物质指标的各个方面得以体现,最终影响其光合生理生态和水分生理生态,这与蒙古沙冬青的实际生长情况很相似,蒙古沙冬青的花期是5月份,而种子采集地5月份的气温一般在30 ℃左右[27],适宜的气温促使蒙古沙冬青开花结果;由参考文献可知,蒙古沙冬青幼苗一般能适应的最高NaCl单盐胁迫质量分数在1.2%~1.3% ,而在本次试验中处理6条件下,即用1.5%盐质量分数的NaCl溶液处理30 d幼苗还有一定的存活率,说明在本次试验条件下,1.5%的NaCl质量分数没有超过蒙古沙冬青的耐受限度,与实地情况相结合,蒙古沙冬青自然分布区的土壤盐碱度较高[28],植株对于单盐胁迫的耐受性比较强[29],但总的来说,随着盐质量分数的不断升高,幼苗的光合及水分各项生理生态指标均受到不同程度的影响,尤其当幼苗从受到低盐质量分数(0.3%)胁迫的开始和受到高盐质量分数(1.2%)胁迫的开始这两个过程中,会在表观、生理和代谢物质指标方面表现出较明显的变化,蒙古沙冬青幼苗在低盐质量分数下表现出较强的抗性,在高盐质量分数下正常生长受到很大抑制。
蒙古沙冬青幼苗在受到温度胁迫时,叶绿素质量分数降低,从而导致其光合速率的下降以及叶片干物质积累的减少,叶片接受光照的能力也降低,光合作用受阻,影响植株正常生命活动,使其表观形态发生变化,具体表现在植株伸长量的减少和比叶面积的增加,说明在蒙古沙冬青受到高温胁迫时,蒙古沙冬青的光系统Ⅱ受到抑制或受损,低温时光系统Ⅱ也会受到抑制从而导致光合速率降低[30];蒙古沙冬青幼苗在受到盐胁迫时,盐质量分数越大,叶绿素质量分数越低,光合速率越低,叶片受光能力也越弱,光合作用受阻,影响植株正常生命活动,使其表观形态发生变化,具体表现在植株伸长量的减少和比叶面积的增加,说明NaCl可以增强叶绿素分解酶活性[31]。
在温盐交互胁迫时,高温和高盐的交互作用会严重影响蒙古沙冬青幼苗的光合生理生态作用,并可以预测40 ℃处理6条件下的植株已经濒临死亡,猜想盐胁迫存在时温度对光系统的破坏也会加剧,通过影响植株光合作用的场所进而对其光合速率产生影响;同时,叶绿素分解酶的最适作用温度可能也比较高,高温条件下该酶活性增强,又受到NaCl存在的促进作用,二者相互叠加,叶绿素分解速率加快而无法积累,进而对植株光合速率和叶片接受光照能力产生影响。
蒙古沙冬青幼苗在受到温度胁迫时,叶片脱水,叶片相对含水量下降,具体表现在水势降低,可溶性糖和脯氨酸质量分数升高以维持植株正常的渗透势和水分生理状态;蒙古沙冬青幼苗在受到盐胁迫时,盐质量分数越大,叶片脱水越严重,叶片相对含水量越低,具体表现在水势降低,可溶性糖和脯氨酸质量分数升高以维持植株正常的渗透势和水分生理状态。脯氨酸作为水溶性很强的氨基酸,对水分有束缚作用,通过提高脯氨酸的质量分数,植物可以减少甚至避免蛋白质变性情况发生在细胞脱水时,所以,提高脯氨酸质量分数是很多植物逆境条件下的自卫反应之一,脯氨酸也是细胞内的重要渗透调节物质,在有效平衡液泡渗透势的同时还可以防止生物大分子被破坏[32]。
在温盐交互胁迫时,高温和高盐的交互作用会严重影响蒙古沙冬青幼苗的水分生理生态作用,蒙古沙冬青处于交互胁迫环境时,其细胞内脯氨酸质量分数随着盐质量分数的升高而迅速升高,表明蒙古沙冬青在应对胁迫时是通过增加脯氨酸质量分数来维持自身细胞渗透压,从而起到应对逆境并且保护自身的作用,虽然可溶性糖也可作为细胞内的渗透调节剂,但试验结果表明,可溶性糖质量分数不如脯氨酸质量分数变化得迅速,同时蒙古沙冬青幼苗叶片中的可溶性糖也未出现大量积累现象,猜测这种现象出现的原因是脯氨酸与可溶性糖发生积累跟蒙古沙冬青所受胁迫程度有关,或者是与蒙古沙冬青受胁迫时期以及所处生长状况有关,蒙古沙冬青不断调节渗透物质类型是根据受胁迫的程度[33],在不同时期甚至不同地点受胁迫也会有相应渗透调节物质积累,这种现象也可能是因为脯氨酸的积累抑制了可溶性糖的积累,脯氨酸和可溶性糖两种物质在调节细胞渗透压时可能会相互补偿[34]。
交互胁迫中,保证植株死亡率较低并可以正常存活的情况下,当控制温度或盐浓度其中一个指标,比较另一指标可得出结论,即当温度处于30 ℃时,处理6与处理1条件下光合和水分各项生理生态指标变化量的差值,大于处理1条件下,40 ℃与30 ℃下光合和水分各项生理生态指标变化量的差值,因此,在本次试验条件下高盐胁迫比高温胁迫对蒙古沙冬青幼苗生长的抑制作用更明显。
中国西北地区地域辽阔,高温区域与盐碱地分布广泛,通过本次探究可为林业生态工程在西北地区播种蒙古沙冬青改善生态环境提供理论依据。但本次试验只探究了温度和盐胁迫对蒙古沙冬青幼苗交互的影响,其他逆境交互胁迫作用还有待进一步研究,关于蒙古沙冬青幼苗在面对逆境胁迫时的调节机制同样需要更深入全面系统地研究和探讨。
[1] 夏恩龙,施海,彭祚登.沙冬青抗逆性与培育技术研究进展[J].中国水土保持科学,2006,4(4):109-113.
[2] 庞涛,郭丽丽,夏新莉,等.CBL1基因5'非翻译区内含子在旱生植物沙冬青中的作用[J].北京林业大学学报,2011,33(6):157-165.
[3] 聂利珍,郭九峰,刘红葵,等.沙冬青脱水素基因转化紫花苜蓿的耐寒性研究[J].西北植物学报,2014,34(9):1727-1734.
[4] 李章磊,高飞,曹玉震,等.蒙古沙冬青AmDREB2.1基因的克隆及表达分析[J].生物技术通报,2015,31(3):108-114.
[5] 郭婷,王茅雁,董博,等.蒙古沙冬青AmDREB2C基因的克隆及表达分析[J].植物遗传资源学报,2015,16(2):344-348.
[6] SUN J, NIE L Z, SUNG Q, et al. Cloning and characterization of dehydrin gene fromAmmopiptanthusmongolicus[J]. MolBiol Rep,2013,10(40):2281-2291.
[7] GULJ, ZHANG H, GUO H M, et al. Cloning ofAmmopiptanthusmongolicusC-repeat-binding factor gene and Its cold-induced tolerance in transgenic tobacco[J] . Brazilian Archives of Biology and Technology,2013,56(3):1516-8913.
[8] SHI J, LIU M Q, SHI J N, et al. Reference gene selection for qPCR inAmmopiptanthusmongolicusunder abiotic stresses and expression analysis of seven ROS-scavenging enzyme genes[J]. Plant Cell Rep,2012,31(7):1245-1254.
[9] 丁志强,尚桂军,李娜,等.沙冬青CBL1蛋白纯化及性质研究[J].中国生物工程杂志,2011,31(2):23-29.
[10] GUL J, CHENG H M. Isolation, molecular cloning and characterization of a cold-responsive gene, Am DUF1517, fromAmmopiptanthusmongolicus[J]. Plant Cell Tiss Organ Cult,2014,117(11):201-211.
[11] YUAN W J, YU Y, YUE Y D, et al. Use of infrared thermal imaging to diagnose health ofAmmopiptanthusmongolicusin northwestern China[J]. For. Res,2015,26(3):605-612.
[12] GUO H M, LI Z C, ZHOU M L, et al. cDNA-AFLP analysis reveals heat shock proteins play important roles in mediating cold, heat, and drought tolerance inAmmopiptanthusmongolicus[J]. FunctIntegr Genomics,2014,14(1):127-133.
[13] 于军,焦培培.聚乙二醇(PEG6000)模拟干旱胁迫抑制矮沙冬青种子的萌发[J].基因组学与应用生物学,2010,29(2):355-360.
[14] 段慧荣,李毅,马彦军.PEG胁迫对沙冬青种子萌发过程的影响[J].水土保持研究,2011,18(3):221-225.
[15] 张娇,张大治,马艳,等.氯化钠模拟盐胁迫对沙冬青种子萌发和幼苗生长的影响[J].北方园艺,2014(14):65-72.
[16] ZHOU Y J, GAO F, LIU R, et al. De novo sequencing and analysis of root transcriptome using 454 pyrosequencing to discover putative genes associated with drought tolerance inAmmopiptanthusmongolicus[J]. BMC Genomics,2012,13(1):266.
[17] 张谧,王慧娟,于长青.超旱生植物沙冬青高温胁迫下的快速叶绿素荧光动力学特征[J].生态环境学报,2009,18(6):2272-2277.
[18] WEI Q, HU P, KUAI B K. Ectopic expression of anAmmopiptanthusmongolicusH+-pyrophosphatase gene enhances drought and salt tolerance inArabidopsis[J]. Plant Cell Tiss Organ Cult,2012,110(1):359-369.
[19] 曹小曙,李平,颜廷真,等.近百年来西辽河流域土地开垦及其对环境的影响[J].地理研究,2005,24(6):889-898.
[20] 张莹,何炎红,田有亮.强光和水分胁迫对沙冬青光呼吸的影响[J].安徽农业科学,2013,41(24):10027-10030.
[21] 蒋志荣.沙冬青抗旱机理的探讨[J].中国沙漠,2000,20(1):71-74.
[22] 张党权,宋志丹,田晔林.抗逆模式灌木沙冬青的研究进展[J].中南林业科技大学学报.2012,32(2):16-20.
[23] 李婧男,刘强,李升.生长素和氯化钙对盐胁迫下沙冬青幼苗的缓解作用[J].植物研究,2010,30(1):27-31.
[24] 刘敏,厉悦,毛子军,等.盐分和温度对万寿菊种子发芽的影响[J].草业科学,2014,31(6):1104-1109.
[25] 胡月楠,贺康宁,王占林,等.盐胁迫对霸王水势的影响[J].中国沙漠,2009,29(5):905-910.
[26] 蔺吉祥,李晓宇,张兆军,等.温度与盐碱胁迫交互作用对羊草种子萌发与幼苗生长的影响[J].草地学报,2011,19(6):1005-1009.
[27] 刘生龙.在干旱沙区适宜引种的野生花卉[J].植物杂志,1992,2(2):17.
[28] 麦尔哈巴·阿布拉.沙冬青植被特征与干旱环境的关系[D].北京:中央民族大学,2015.
[29] 夏恩龙.沙冬青地理分布与在北京地区引种栽培技术的研究[D].北京:北京林业大学,2006.
[30] 宋祥春,赵惠新,苗玉,等.低温对两种沙冬青幼苗光合生理指标的影响[J].新疆大学学报(自然科学版),2009,26(3):342-346.
[31] 李婧男,刘强,贾志宽,等.盐胁迫对沙冬青幼苗生长与生理特性的影响[J].植物研究,2009,29(5):553-558.
[32] 赵峰侠,尹林克,牟书勇.新疆沙冬青(Ammopiptanthusnanus)渗透调节物质的季节变化与环境因子的关系[J].干旱区地理,2008,121(5):665-672.
[33] 王华,贾桂霞,丁琼.沙冬青抗逆性研究进展与应用前景[J].中国农学通报,2005,21(12):121-125.
[34] 郭华军.水分胁迫过程中的渗透调节物质及其研究进展[J].安徽农业科学,2010,38(15):7750-7753,7760.
猜你喜欢
中老年保健(2022年3期)2022-11-21
中华建设(2020年5期)2020-07-24
科学与财富(2019年15期)2019-10-21
阅读与作文(小学高年级版)(2019年8期)2019-10-16
蒙古学问题与争论(2019年0期)2019-03-29
西域历史语言研究集刊(2018年0期)2018-11-09
小学生必读(低年级版)(2018年6期)2018-09-08
发酵科技通讯(2018年2期)2018-07-06
草原歌声(2017年1期)2017-04-23
——致坚守奋斗的你
音乐天地(音乐创作版)(2017年1期)2017-02-12
