
长江口及邻近海域浮游动物生物量分布及季节变化
2018-01-15 10:04:13李晓慧刘镇盛
海洋学研究 2017年4期
李晓慧,刘镇盛*
(1. 国家海洋局 第二海洋研究所,浙江 杭州 310012;2. 国家海洋局 海洋生态系统与生物地球化学重点实验室,浙江 杭州 310012)
0 引言
浮游动物在海洋生态系统中扮演重要角色,它既是浮游植物和细菌的主要消耗者,又充当海洋的次级生产者,作为更高营养阶层海洋动物的饵料,浮游动物在海洋食物网中起关键作用[1-2]。浮游动物在昼夜垂直迁移生长代谢过程中,将有机碳从海表层迁移至海洋深处,有效削弱大气的温室效应,浮游动物既是海洋生态系统食物网结构与功能研究的关键环节,也是生物地球化学和生物海洋学研究的重点[3]。浮游动物生物量的分布特征,是相关海区次级生产力水平、渔业资源开发潜力和环境质量综合评价等的重要指标之一[4]。长江口及邻近海区受到陆缘径流和陆架水系物理和化学因素的共同作用,又是许多重要经济鱼类的产卵场和育幼场,这些人为和自然因素对浮游动物生物量的分布有重大影响[5]。1980年以来,对长江口及邻近海域浮游动物研究已有大量报道[6-14],但主要集中在长江河口区或东海陆架区,而从长江零盐度分界线开始延伸至长江冲淡水扩散海域(至127°E)中尺度海域的研究较薄弱[15]。本文报道了长江口及邻近海域浮游动物生物量的分布及季节变化,并结合同步观测的环境因子调查结果,探讨了长江口及邻近海域浮游动物生物量的季节变化及与环境的关系,为长江口及邻近海域海洋资源可持续利用和海洋生态系统研究提供科学依据。
1 材料和方法
1.1 研究海域
2006年7月—8月(夏季)、2006年12月—2007年2月(冬季)、2007年4月—5月(春季)、2007年10月—12月(秋季)对长江口及邻近海域海洋生物和水体环境进行了4个航次的综合调查。在调查海域,共布设150个观测站位。调查海域范围:29°30′N~32°30′N,120°E~127°30′E(图1)。4个调查航次均由“中国海监49船”执行。
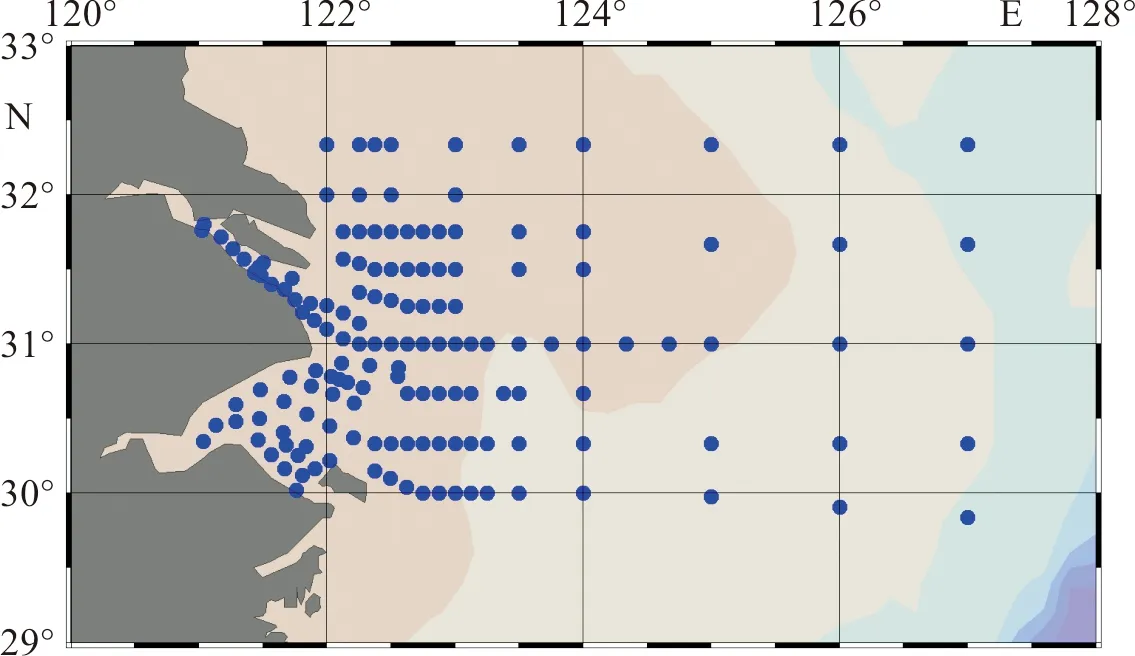
图1 长江口及邻近海域浮游动物采样站位图Fig.1 Sampling stations in the Changjiang Estuary and its adjacent waters
1.2 调查方法
浮游动物现场采样中,使用装有进口流量计(德国HYDRO-BIOS公司-浮游生物网口流量计E-Fiow)的浅I型浮游生物网(网长145 cm,网口内径50 cm,筛绢孔径0.505 mm,采集水深<30 m的浮游动物)或大型浮游生物网(网长280 cm,网口内径80 cm,筛绢孔径0.505 mm,采集水深>30 m的浮游动物)进行垂直拖网采样。海域水深<200 m时,从底层至表层进行垂直拖网;水深>200 m时,从200 m至表层垂直拖网采集样品。将每个测站采集的浮游生物样品装入600 cm3的塑料瓶中,加入5%(v/v)甲醛溶液固定保存,带回实验室分析鉴定。
同步观测的参数有水温、盐度、水深、溶解氧、硝酸盐、磷酸盐、硅酸盐、叶绿素a和初级生产力等。以上采样、观测和分析测定方法均参照GB/T 12763 6—2007海洋调查规范[16]相关规定执行。
1.3 浮游动物生物量分析和计算方法
在实验室内剔除浮游动物样品中的杂物,利用置于漏斗上的筛绢截留浮游动物样品,将筛绢放在滤纸上吸去水分后,用电子天平(Sartorius CP124S)称量浮游动物生物量,记录结果。将称量完成后的样品置于体视显微镜下,对样品进行种属鉴定和个体计数,分析方法按GB/T 12763 6—2007海洋调查规范[16]的规定执行。
浮游动物湿重生物量计算公式:
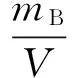
(1)
式中:PB为单位体积海水浮游动物的湿重含量,单位为mg/m3;mB为样品湿重含量,单位为mg;V为滤水量,单位为m3。
1.4 数据处理
本研究采用Ocean Date View 4绘图软件绘制调查站位图和浮游动物生物量平面分布图,使用SPSS 20.0软件对浮游动物生物量和环境因子进行Pearson相关分析。本研究的环境因子主要包括水温、盐度、溶解氧、硝酸盐、磷酸盐、硅酸盐、悬浮物和叶绿素a质量浓度。
2 结果
2.1 浮游动物生物量分布及季节变化
长江口及邻近海域浮游动物生物量有明显季节变化。浮游动物生物量季节变化特征为:春季>夏季>秋季>冬季(表1)。春季浮游动物生物量最高,平均值达368 mg/m3,冬季浮游动物生物量最低,平均值为65 mg/m3,夏季高于秋季,生物量平均值分别为334和180 mg/m3。
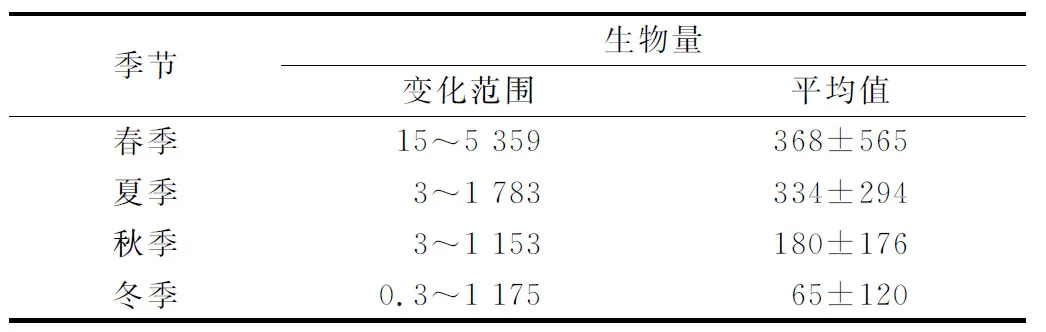
表1 长江口及邻近海域浮游动物生物量的季节变化(2006年7月—2007年12月)Tab.1 Seasonal variation of zooplankton biomass in the Changjiang Estuary and its adjacent waters(Jul. 2006 to Dec. 2007) mg/m3
长江口及邻近海域浮游动物生物量分布有明显区域特征。
春季,舟山群岛邻近海域,浮游动物生物量形成高值区(>1 000 mg/m3),高值区中心的浮游动物生物量达5 000 mg/m3以上。浮游动物生物量最高值出现在舟山群岛东侧海域(29.0°N~30.0°N,122°E~124°E),达5 359 mg/m3。长江口及杭州湾海域,出现浮游动物生物量低值区。从舟山群岛东侧海域向北部及陆架外海,浮游动物生物量呈逐渐下降趋势(图2a)。
夏季,在长江口门区海域,出现浮游动物生物量高值区(>600 mg/m3)。浮游动物生物量最高值出现在31.0°N~32.0°N,123°E~124°E海域,达1 783 mg/m3。东海陆架外海海域,生物量普遍较低(<100 mg/m3),从长江口高值区往陆架外海方向,浮游动物生物量呈梯度下降趋势(图2b)。
秋季,在长江口北部与黄海毗邻海域(31.0°N~33.0°N,125°E~126°E),出现浮游动物生物量高值区(>500 mg/m3),高值区中心浮游动物生物量达1 000 mg/m3以上。长江口及邻近海域以该生物量高值区为中心,其周边水域生物量逐渐下降,杭州湾和东海外海陆架水域生物量普遍较低(图2c)。
冬季,浮游动物生物量明显下降,为四季中最低。在舟山群岛东北海域(29.5°N~31.0°N,124°E~125°E)形成两个生物量高值区(>250 mg/m3),高值区中心浮游动物生物量达500 mg/m3以上。以生物量高值区为中心,其周围的浮游动物生物量呈逐渐递减趋势(图2d)。
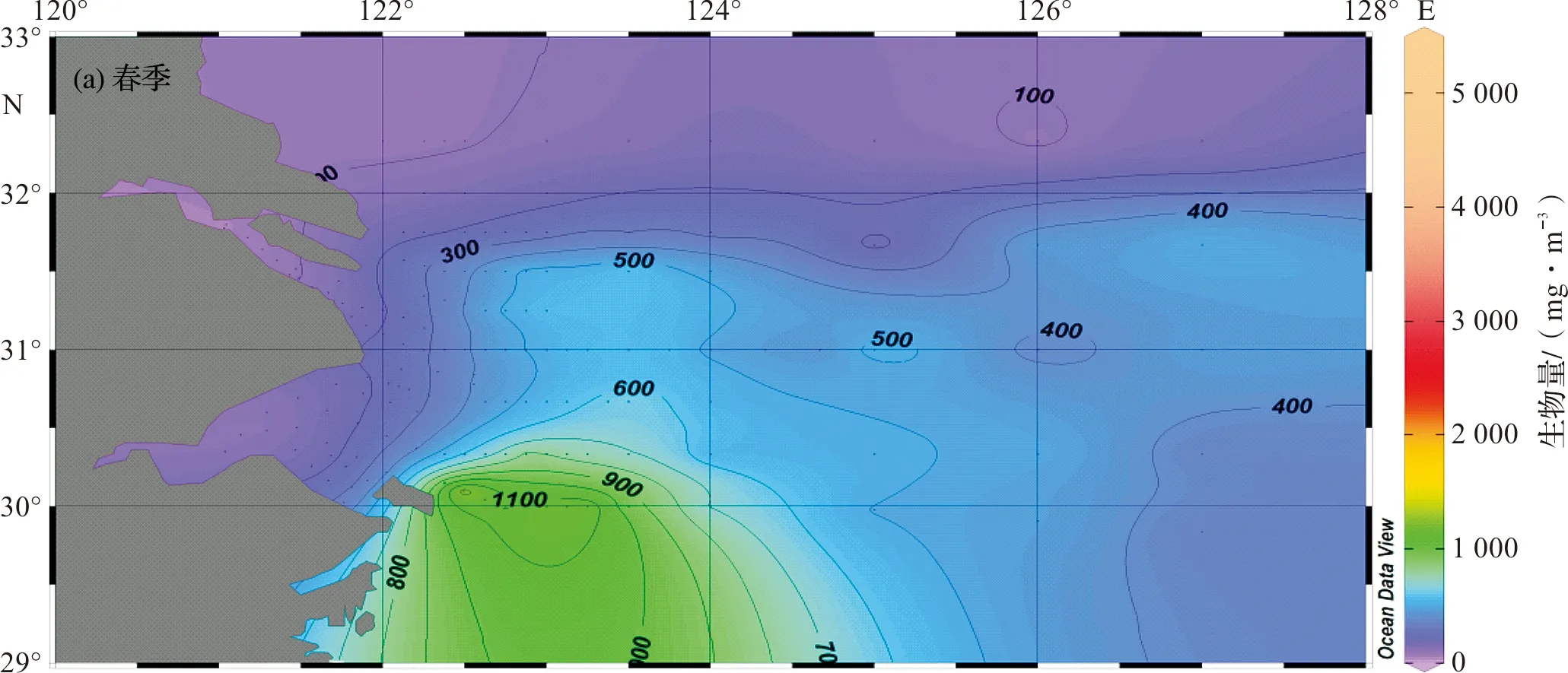
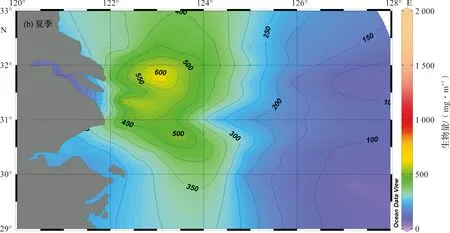
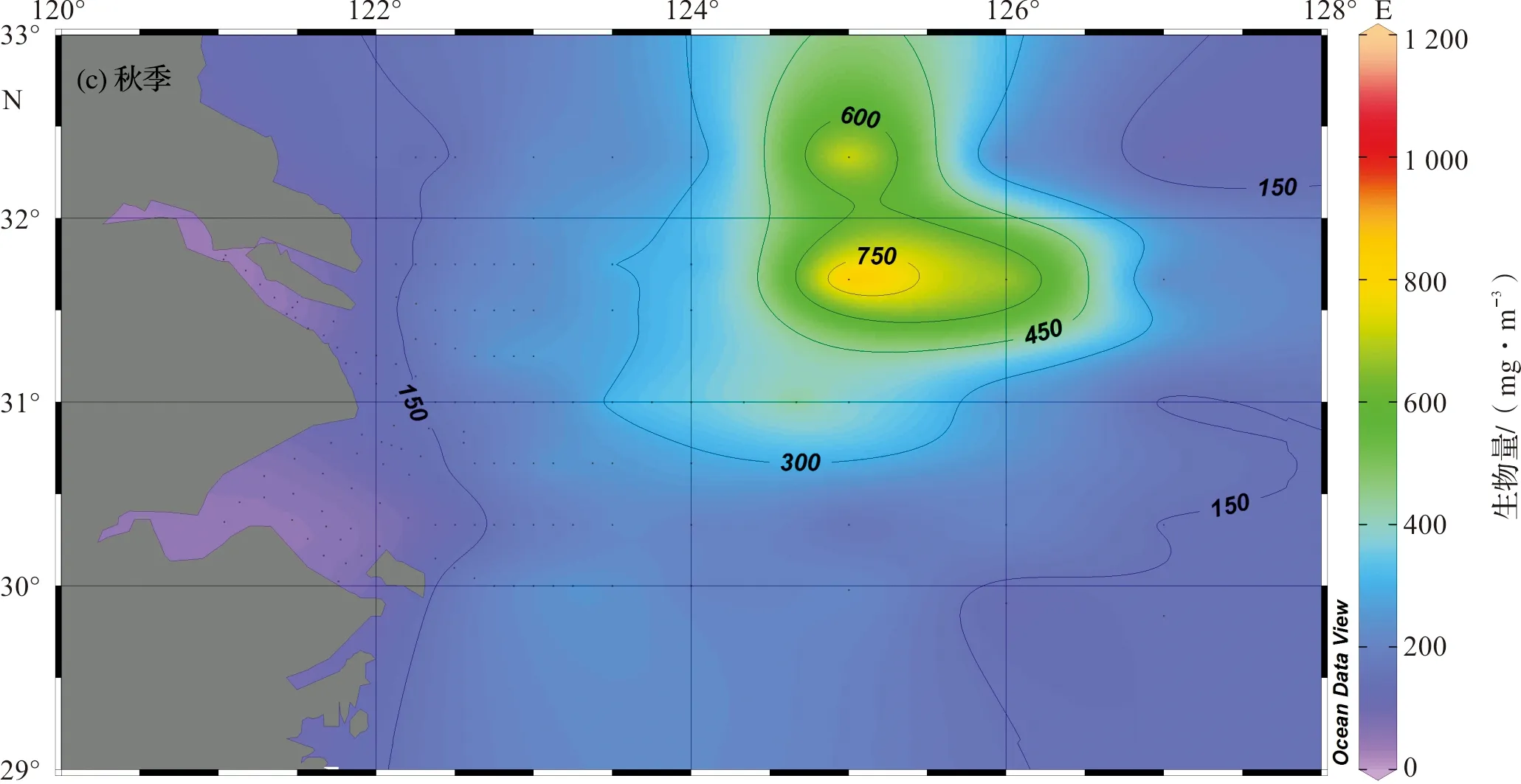
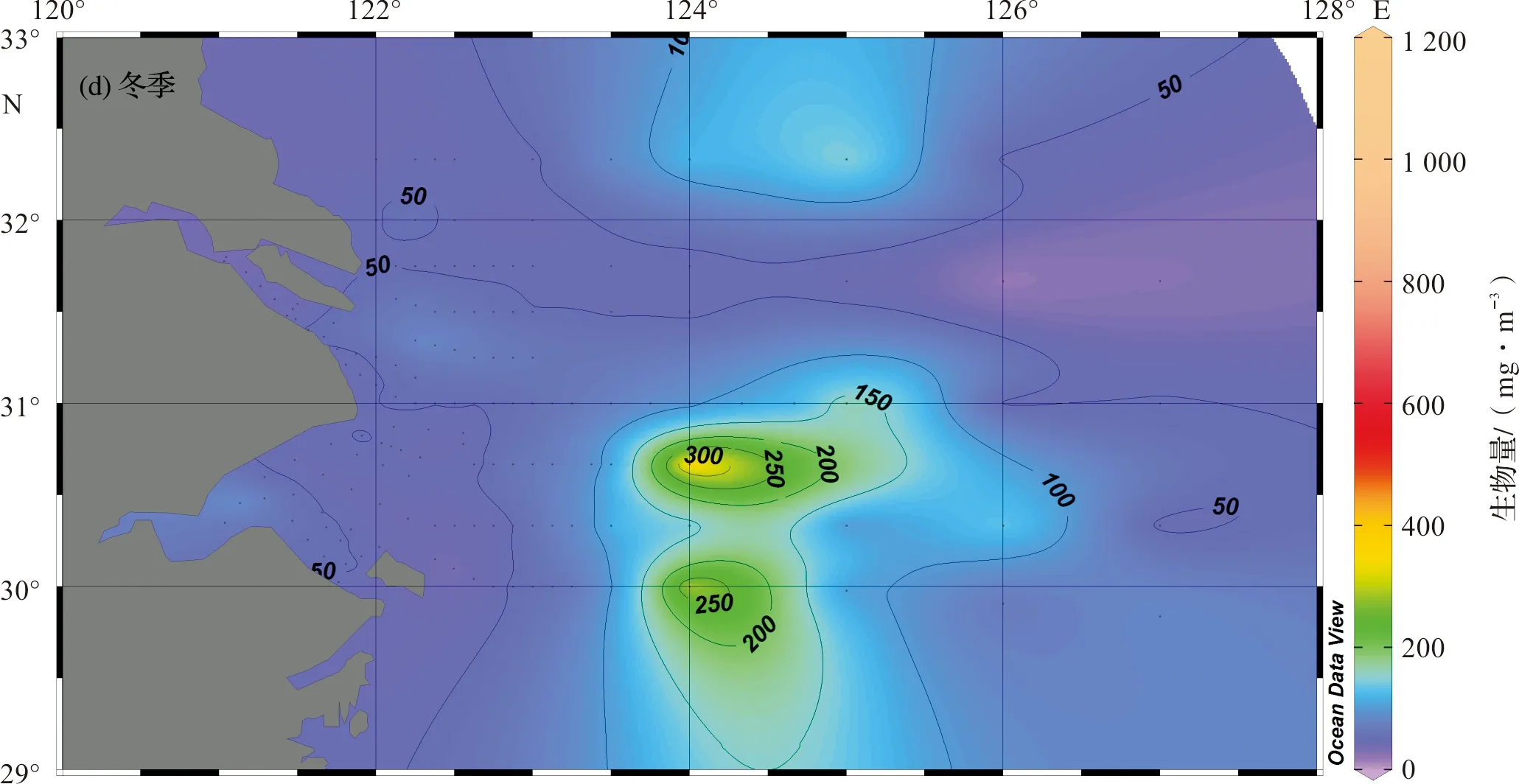
图2 长江口及邻近海域浮游动物生物量水平分布Fig.2 Horizonal distribution of zooplankton biomass in the Changjiang Estuary and its adjacent waters
2.2 浮游动物生物量与环境因子的关系
浮游动物生物量与环境因子的Pearson相关分析结果表明(表2):春季,浮游动物生物量与水温和盐度呈现极显著正相关关系(P<0.01),而与硝酸盐、磷酸盐和硅酸盐存在极显著负相关关系(P<0.01)。夏季,浮游动物生物量与水温呈现极显著负相关关系(P<0.01),与硝酸盐、磷酸盐和硅酸盐存在显著负相关关系(P<0.05),与溶解氧存在显著正相关关系(P<0.05),与叶绿素a呈现极显著的正相关关系(P<0.01)。秋季,浮游动物生物量与盐度呈现极显著正相关关系(P<0.01),而与硝酸盐、磷酸盐、硅酸盐、溶解氧和悬浮颗粒物呈现极显著负相关关系(P<0.01)。冬季,浮游动物的生物量与其它环境因子不存在明显相关关系。
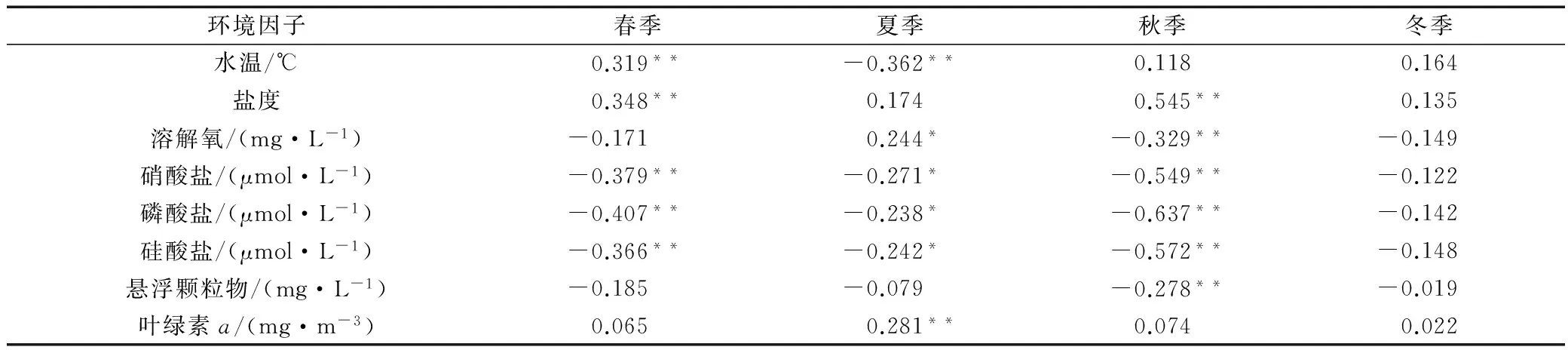
表2 长江口及邻近海域浮游动物生物量与环境因子的相关性(2006年7月—2007年12月)Tab.2 Relative coefficient among biomass of zooplankton and environment factors in the Changjiang Estuary and its adjacent waters (Jul. 2006 to Dec. 2007)
注:**表示极显著相关,P=0.01;*表示显著相关,P=0.05
3 讨论
3.1 长江口及邻近海域浮游动物生物量时空变化
由于长江口及邻近海域四季水温、盐度等环境因子波动幅度大,对浮游动物的生物量分布产生重要影响[17-18],因此,长江口及邻近海域浮游动物生物量有明显的时空变化。本研究表明,影响浮游动物生物量的主要是季节变化。
春季,中华哲水蚤(Calanussinicus)、百陶带箭虫(Zonosagittabedoti)、双生水母(Diphyeschamissonis)和中华假磷虾(Pseudeuphausiasinica)是浮游动物生物量的主要贡献者,其种群丰度的分布及变动直接影响着长江口及邻近海域浮游动物生物量的变化。在舟山群岛东南部邻近海域浮游动物生物量高值区,浮游动物主要由中华哲水蚤和双生水母构成。
夏季,随着水温上升,浮游动物生长代谢速率增大,东海黑潮分支流和台湾暖流对长江口及邻近海域也产生重要影响。在长江口门浮游动物生物量高值区,主要由背针胸刺水蚤(Centropagesdorsispinatus)和短额超刺糠虾(Hyperacanthomysisbrevirostris)等构成。长江口及邻近海域,除了背针胸刺水蚤和短额超刺糠虾之外,中华哲水蚤和百陶带箭虫也是影响浮游动物生物量的主要种类,这与刘守海 等[13]报道的结果基本一致。
秋季,随着水温的下降,浮游动物生长繁殖速率降低,生物量逐渐下降。长江口与黄海毗邻陆架海域浮游动物生物量高值区,主要由中华哲水蚤和肥胖软箭虫(Flaccisagittaenflata)等组成。此外,亚强次真哲水蚤(Subeucalanussubcrassus)、百陶带箭虫也是影响长江口及邻近海域浮游动物生物量的主要贡献种[15]。
冬季,长江口及邻近海域受北方冷空气和寒流的影响,水温明显下降,多种浮游动物暖水种消失,浮游动物生物量为四季最低值。在舟山群岛东北海域生物量高值区,主要由中华哲水蚤和百陶带箭虫等构成。此外,真刺唇角水蚤(Labidoceraeuchaeta)和中华假磷虾等也是影响冬季长江口及邻近海域浮游动物生物量的主要种类。
近50 a历史资料及文献反映了本研究海域浮游动物生物量的变动趋势(表3)。20世纪50s末期,长江口及邻近海域浮游动物生物量较低,至1980s中期,该海域生物量没有明显变化[8]。1990s末期以来,长江口海域浮游动物生物量出现升高趋势,且生物量波动幅度大[9-11]。本研究表明,长江口及邻近海域浮游动物生物量的季节变化趋势与同期调查的东海和黄海的变动趋势一致[6],但与徐兆礼 等[10]报道的结果存在一些差异。因本研究范围覆盖长江内河段至长江冲淡水扩散区陆架外海至127°E海域,而1980s以来,有关长江口及邻近海域浮游动物文献报道的研究海域主要集中在124°E以西海域,其调查海域范围相对较小且离岸更近,与本研究的调查海域范围存在空间差异,因而,统计分析结果有所不同。另外,春季,本研究海域少量测站出现大量聚集性的尾索动物,导致该季节部分测站生物量出现异常高值,对长江口及邻近海域平均生物量统计也可能产生影响。
3.2 环境因子对长江口及邻近海域浮游动物生物量的影响
长江口及邻近海域受长江冲淡水、台湾暖流、江浙近岸流、黑潮分支流等多个水系的共同作用和影响,形成独特的海洋环境,浮游动物生物量的分布与水温、盐度、营养盐等环境因子密切相关[19],在不同季节,影响浮游动物生物量的环境因子有明显变化。
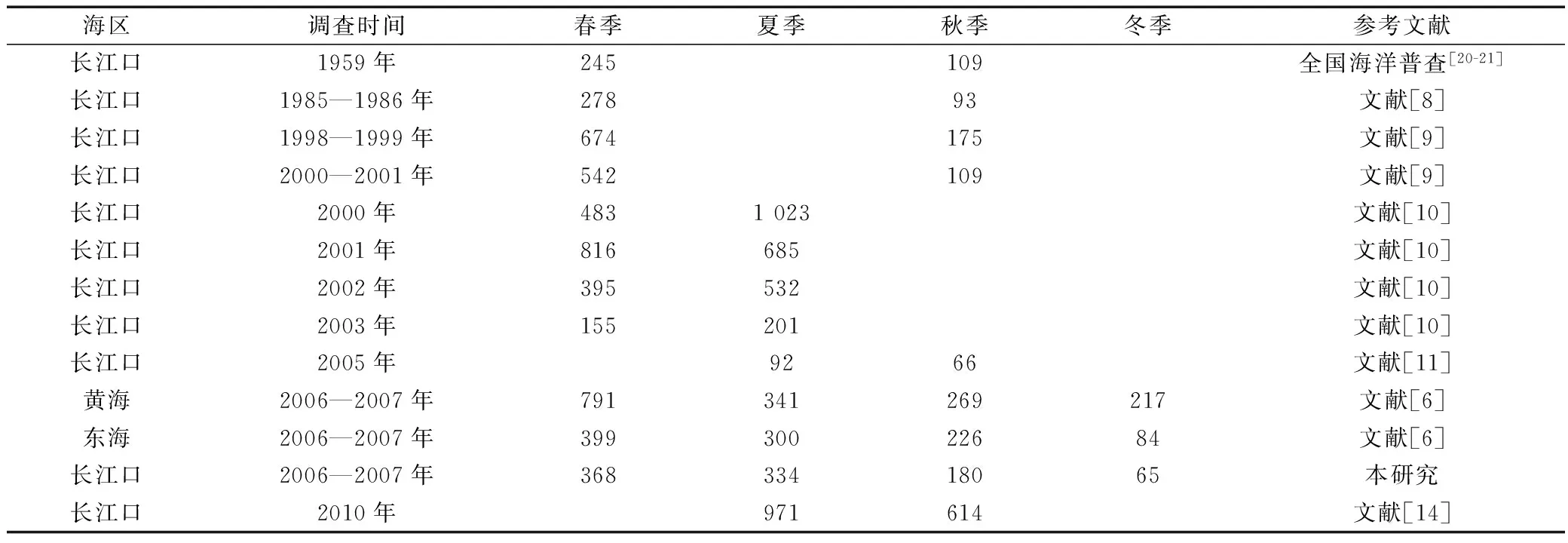
表3 长江口及邻近海域浮游动物生物量的变化Tab.3 Changes in biomass of zooplankton in the Changjiang Estuary and its adjacent waters mg/m3
注:空白处表示无数据
温度是影响春、夏季浮游动物生物量的主要环境因子。相关分析表明(表2),春季研究海域浮游动物生物量与水温和盐度呈显著正相关,这与王荣 等[22]报道的研究结果一致。水温的回升,有利于浮游动物生长代谢和幼体发育,营养盐为浮游植物生长繁殖提供物质基础,而浮游动物是浮游植物的主要消费者,与浮游植物形成动态平衡关系[23-24]。夏季,随着水温的升高,长江口及邻近海域的浮游植物现存量出现峰值,浮游植物生长速率增强导致溶解氧的浓度上升,为浮游动物生长繁殖提供丰富饵料和良好的生存环境,有利于浮游动物生长代谢,浮游动物生物量与叶绿素a、溶解氧呈显著正相关。然而,水温的大幅升高也影响浮游动物群落重要种群(如:中华哲水蚤)的生长繁殖[25-26],因此,夏季浮游动物生物量与水温呈显著负相关。
秋季,长江等陆源入海径流量减少,水体营养盐浓度下降,台湾暖流及黑潮分支流对长江口及邻近海域的控制势力增强,浮游动物群落中高温高盐种比例上升,浮游动物生长代谢消耗浮游植物现存量,浮游植物的生长繁殖空间变大。而营养盐的供给对浮游植物的生长繁殖起主要调控作用,浮游植物的生长消耗营养盐。这可能是导致浮游动物与营养盐呈显著负相关的原因。浮游植物在光合作用过程中吸收营养盐,释放氧气,使水体中溶解氧浓度上升,而浮游植物死亡时,有机质降解则会消耗水体中的溶解氧[27]。秋季,长江口及邻近海域悬浮颗粒物质量浓度较高,透明度降低,不利于浮游植物生长繁殖,导致叶绿素a质量浓度下降,溶解氧的浓度下降。受台湾暖流及黑潮分支流的影响,浮游动物群落中高温高盐种比例上升,使浮游动物生物量与溶解氧的浓度呈显著负相关,这与崔彦萍 等[28]的研究结果一致。
冬季,受北方冷空气和寒流影响,研究海域浮游动物群落中高温高盐种消失,浮游动物生长代谢速率下降,浮游动物生物量与环境因子不存在明显相关关系。
致谢章菁和夏平参加了本项目采样和分析工作;国家海洋局第二海洋研究所陈建芳研究员提供了长江口海域水化学数据,黄大吉研究员提供了温、盐数据,郝强博士提供了叶绿素a数据;“海监49号”科学考察船全体船员在海上采样过程中给予了帮助;孙栋博士对本文写作提出宝贵建议,在此一并致以谢忱!
[1]LIShao-jing,XUZhen-zu,HUANGJia-qi,etal.StudiesonbiologyofmarinezooplanktoninChina[J].JournalofXiamenUniversity:NaturalScience,2001,40(2):574-585.
李少菁,许振祖,黄加祺,等.海洋浮游动物学研究[J].厦门大学学报:自然科学版,2001,40(2):574-585.
[2]XUZhao-li.Advancesandfutureofourstudyonmarinezooplankton[J].JournalofXiamenUniversity:NaturalScience,2006,45(Sup.2):16-23.
徐兆礼.中国海洋浮游动物研究的新进展[J].厦门大学学报:自然科学版,2006,45(增刊2):16-23.
[3]HAYSG.Areviewoftheadaptivesignificanceandecosystemconsequencesofzooplanktondielverticalmigration[J].Hydrobiologica,2003,503:163-170.
[4]DUFei-yan,LIChun-hou,LIAOXiu-li,etal.ThevarietyofzooplanktonbiomassinDayabay[J].MarineEnvironmentalScience,2006,25(1):37-39,43.
杜飞雁,李纯厚,廖秀丽,等.大亚湾海域浮游动物生物量变化特征[J].海洋环境科学,2006,25(1):37-39,43.
[5]MORGANCA,ROBERTISAD,ZABELRW.ColumbiaRiverplumefronts.I.Hydrography,zooplanktondistribution,andcommunitycomposition[J].MarEcolProgSer,2005,299:19-31.
[6]WANGChun-sheng,CHENXing-qun,LIUZhen-sheng.Chinaoffshoresea:Marinebiologyandecology[M].Beijing:ChinaOceanPress,2012.
王春生,陈兴群,刘镇盛.中国近海海洋:海洋生物与生态[M].北京:海洋出版社,2012.
[7]TANGQi-sheng.BiologicalresourcesandhabitatenvironmentofseaareainChina’sexclusiveeconomiczone[M].Beijing:SciencePress,2006:307-363.
唐启升.中国专属经济区海域生物资源与栖息环境[M].北京:科学出版社,2006:307-363.
[8]GAOShang-wu,ZHANGHe-qing.StudiesonzooplanktonecologyintheChangjiangRiverEstuary[J].StudiaMarinaSinica,1992,33:201-216.
高尚武,张河清.长江口区浮游动物生态研究[J].海洋科学集刊,1992,33:201-216.
[9]WANGKe,WANGRong,ZUOTao,etal.AnalysisofzooplanktonbiomassintheChangjiangRiverEstuaryandadjacentwaters[J].OceanologiaetLimnologiaSinica,2004,35(6):568-576.
王克,王荣,左涛,等.长江口及邻近海区浮游动物总生物量分析[J].海洋与湖沼,2004,35(6):568-576.
[10]XUZhao-li,SHENXin-qiang.ZooplanktonbiomassanditsvariationinwaternearChangjiangEstuary[J].ResourcesandEnvironmentintheYangtzeBasin,2005,14(3):282-286.
徐兆礼,沈新强.长江口水域浮游动物生物量及其年间变化[J].长江流域资源与环境,2005,14(3):282-286.
[11]ZHANGFei-yan,TANGJing-liang,LIDao-ji,etal.ZooplanktondistributionandvariationintheYangtzeEstuaryanditsadjacentwatersinsummerandautumn[J].ActaHydrobiologicaSinica,2009,33(6):1 219-1 215.
章飞燕,唐静亮,李道季,等.夏、秋季长江口及其毗邻海域浮游动物的分布与变化[J].水生生物学报,2009,33(6):1 219-1 215.
[12]ZHUYan-zhong,LIULu-san,ZHENGBing-hui,etal.RelationshipbetweenspatialdistributionofzooplanktonandenvironmentalfactorsintheChangjiangEstuaryanditsadjacentwatersinspring[J].MarineSciences,2011,35(1):59-65.
朱延忠,刘录三,郑丙辉,等.春季长江口及毗邻海域浮游动物空间分布及与环境因子的关系[J].海洋科学,2011,35(1):59-65.
[13]LIUShou-hai,XIANGLing-yun,LIUCai-cai,etal.EcologicaldistributioncharacteristicsofzooplanktoninYangtzeRiverEstuaryinspringandsummerduring2007—2008[J].MarineScienceBulletin,2013,32(2):184-190.
刘守海,项凌云,刘材材,等.2007—2008年春夏季长江口水域浮游动物生态分布特征研究[J].海洋通报,2013,32(2):184-190.
[14]WANGLi,WANGBao-dong,CHENQiu-wen,etal.CharacteristicsofthezooplanktoncommunityandimpactfactorsintheYangtzeEstuarycoastalareaafterthirdstageimpoundmentoftheThreeGorgesDam[J].ActaEcologicaSinica,2016,36(9):2 505-2 512.
王丽,王保栋,陈求稳,等.三峡三期蓄水后长江口海域浮游动物群落特征及影响因子[J].生态学报,2016,36(9):2 505-2 512.
[15]SHAOQian-wen,LIUZhen-sheng,ZHANGJing,etal.SeasonalvariationinzooplanktoncommunitystructureintheChangjiangEstuaryanditsadjacentwaters[J].ActaEcologicaSinica,2017,37(2):1-9.
邵倩文,刘镇盛,章菁,等.长江口及邻近海域浮游动物群落结构及季节变化[J].生态学报,2017,37(2):1-9.
[16]GB/T12763 6—2007SpecificationsforoceanographicsurveyPart6:Marinebiologicalsurvey[S].2007.
GB/T12763 6—2007海洋调查规范第6部分:海洋生物调查[S].2007.
[17]WUFeng-run,TONGChun-fu.EvolutionoftheNorthBranchofYangtzeEstuaryinlast30yearsandcorrespondingeffectsonspeciesdiversity[J].HaiyangXuebao,2017,39(2):72-85.
吴逢润,童春富.近30年长江口北支演变及其对物种多样性的影响[J].海洋学报,2017,39(2):72-85.
[18]LUWu-yang,MAZeng-ling,XUZhao-li,etal.VariationofzooplanktoncommunityinChineseestuariesalonglatitudesinspring[J].HaiyangXuebao,2016,38(10):83-93.
卢伍阳,马增岭,徐兆礼,等.春季我国不同纬度河口浮游动物群落变化趋势[J].海洋学报,2016,38(10):83-93.
[19]WUXiao-dan,SONGJin-ming,LIXue-gang.SeasonalvariationofwatermasscharacteristicandinfluenceareaintheYangtzeEstuaryanditsadjacentwaters[J].MarineSciences,2014,38(12):110-119.
吴晓丹,宋金明,李学刚.长江口邻近海域水团特征与影响范围的季节变化[J].海洋科学,2014,38(12):110-119.
[20]Nationalmarinecomprehensivesurveyreport:Volumeeight[R].Tianjin:MarineGeneralInvestigationOffice,1977.
全国海洋综合调查报告:第八册[R].天津:海洋综合调查办公室,1977.
[21]Nationalmarinecomprehensivesurveyreport:Volumeone[R].Tianjin:MarineGeneralInvestigationOffice,1961.
全国海洋综合调查资料报告:第一册[R].天津:海洋综合调查办公室,1961.
[22]WANGRong,CHENYa-qu,WANGKe.EstimationoftheannualproductionofCalanussinicusBrodsky(Copepoda:Calanoida)intheEastChinaSea[J].ActaEcologicaSinica,2003,23(6):1 212-1 215.
王荣,陈亚瞿,王克,等.东海中华哲水蚤的年产量估算[J].生态学报,2003,23(6):1 212-1 215.
[23]ZHOUWei-hua,WANGHan-kui,DONGJun-de,etal.PhytoplanktonandbacterialbiomassandtheirrelationshipwiththeenvironmentalfactorsinautumnandwinterintheSanyaBay,China[J].ActaEcologicaSinica,2006,26(8):2 633-2 639.
周伟华,王汉奎,董俊德,等.三亚湾秋、冬季浮游植物和细菌的生物量分布特征及其与环境因子的关系[J].生态学报,2006,26(8):2 633-2 639.
[24]ZENGXiang-bo,HUANGBang-qin.Seasonalvariationsinabundance,biomassandestimatedproductionratesofmicrozooplanktonatWestofXiamenwaters[J].JournalofXiamenUniversity:NaturalScience,2010,49(1):109-115.
曾祥波,黄邦钦.厦门西海域微型浮游动物的丰度、生物量及其生产力的季节变动[J].厦门大学学报:自然科学版,2010,49(1):109-115.
[25]SUNSong.Preliminarystudyontheover-summermechanismofCalanussinicusintheYellowSea[J].OceanologiaetLimnologiaSinica:ZooplanktonResearchAlbums, 2005,36(Supplement):92-99.
孙松.黄海中华哲水蚤度夏机制初探[J].海洋与湖沼:浮游动物研究专辑,2005,36(增刊):92-99.
[26]WANGR,ZOUT,WANGK.TheYellowSeacoldbottomwateranoversummeringsiteforCalanussinicus(Copepods,Crustacea)[J].JPlanktonRes,2003,25(2):169-183.
[27]ZHANGYing-ying,ZHANGJing,WUYing,etal.CharacteristicsofdissolvedoxygenanditsaffectingfactorsintheYangtzeEstuary[J].EnvironmentalSciences,2007,28(8):1 649-1 654.
张莹莹,张经,吴莹,等.长江口溶解氧的分布特征及影响因素研究[J].环境科学,2007,28(8):1 649-1 654.
[28]CUIYan-ping,WANGBao-dong,CHENQiu-wen.Thespatial-temporaldynamicsofchlorophyllaandDOintheYangtzeestuaryafternormalimpoundmentoftheThreeGorgesReservoir[J].ActaEcologicaSinica,2014,34(21):6 309-6 316.
崔彦萍,王保栋,陈求稳.三峡正常蓄水后长江口叶绿素a和溶解氧变化及其成因[J].生态学报,2014,34(21):6 309-6 316.
猜你喜欢
现代农村科技(2022年9期)2022-08-16 02:25:34
中国麻业科学(2021年5期)2021-12-02 02:08:52
生物学通报(2021年9期)2021-07-01 03:24:44
辽宁化工(2021年5期)2021-06-03 05:15:50
——以“不同咖啡因饮料对水蚤心率的影响”为例
中学生物学(2021年2期)2021-04-30 12:04:06
海洋渔业(2016年6期)2016-04-16 03:00:35
海洋渔业(2016年6期)2016-04-16 03:00:32
中国资源综合利用(2016年6期)2016-01-22 07:28:51
湖南农业(2015年7期)2015-03-20 01:48:37
华东师范大学学报(自然科学版)(2014年3期)2014-03-11 16:18:19
