
高油酸油菜脂肪酸代谢的蛋白质组学与转录组学关联分析
2018-01-12 01:07王晓丹张振乾官春云邬贤梦
华北农学报 2017年6期
王晓丹,肖 钢,常 涛,张振乾,官春云,邬贤梦
(湖南农业大学 农学院,南方粮油作物协同创新中心,湖南 长沙 410128)
含油量与脂肪酸组成是评价油菜品质的关键指标[1]。低芥酸、低α-亚麻酸与高油酸、高亚油酸一直是我们所追求的营养品质方面的重要育种目标之一,目前育成的高油酸油菜籽粒油酸含量高于75%,有些甚至高达90%以上[2-3]。高油酸菜籽油有较高的营养,且有利于人体心血管健康,因而近年来有较多高油酸油菜相关的研究[4-6],但其脂肪酸代谢分子生物学机理研究仍未完全清楚。
同位素标记相对和绝对定量(Isobaric tags for relative and absolute quantitation,iTRAQ)技术定量精确、有较高的鉴定率,可鉴定多达6 000种蛋白,是近年来定量蛋白质组学常用的高通量筛选技术[7-8]。转录组测序是一种高通量测序技术,精确性高,成本低,得到了广泛的应用[9-10]。蛋白质组与转录组联合分析可以从不同层面研究基因表达的情况,得到生物体更加完整的表达信息[11]。目前,利用蛋白质与转录组进行关联分析的研究在水稻、拟南芥、紫花苜蓿等许多植物中有了应用[12-14],但在高油酸油菜上仍未有相关应用。
为了更好地了解高油酸油菜的调控机理,本研究以一组高油酸近等基因系材料自交授粉后20~35 d新鲜、无病害的种子为材料,结合iTRAQ技术与转录组测序技术,进行蛋白质组与转录组的关联分析,以期找到其中显著差异表达的蛋白与基因,促进对其相关代谢路径的分子机理研究。
1 材料和方法
1.1 试验材料
高油酸油菜近等基因系材料由湖南农业大学油料作物研究中心提供,高低油酸品系油酸含量分别为81.4%,56.2%。2012年9月28日种植于湖南农业大学耘园基地,取自交套袋20~35 d的种子混合保存于超低温冰箱(-80 ℃)备用。
1.2 iTRAQ联合二维液相色谱-串联质谱(2D LC-MS/MS)分析差异表达蛋白
采用TCA/丙酮法提取蛋白后,测定蛋白质浓度(Bradford定量法),并进行电泳检测(12%的SDS聚丙烯酰胺凝胶)。37 ℃取等量蛋白加入胰蛋白酶酶解。再等量混合用iTRAQ试剂标记的酶解肽段,用强阳离子交换色谱(Strong cation exchange choematography,SCX)进行预分离。然后进行液相串联质谱(Liquid chromatography coupled with tandem mass spectrometry,LC-MS/MS)分析。另外,本研究所用数据库为白菜与甘蓝数据库,以Mascot 2.3.02对蛋白质进行鉴定分析,该试验由深圳华大公司完成。
1.3 转录组测序分析差异表达基因
用TRNzol法提取授粉后20~35 d种子的RNA,检测合格后,将该种子样品送往深圳华大公司测序。
1.4 关联分析
以低油酸材料为对照,通过上述结果,以蛋白表达差异倍数≥1.5,P值≤0.05:基因表达差异倍数≥2,P值≤0.001为标准,筛选差异表达的基因和蛋白质,进行关联分析,计算Person相关系数。
2 结果与分析
2.1 蛋白质组学与转录组学分析
2.1.1 iTRAQ定量蛋白质组数据 通过iTRAQ技术共得到谱图340 528张,Mascot 2.3.02分析匹配到67 334张,共鉴定到4 726个蛋白。按1.4筛选差异表达蛋白的标准,共得到334个差异蛋白,其中148个上调表达,186个下调表达(表1)。
2.1.2 转录组测序数据 通过Illumina HiseqTM2000测序及除杂处理后,有24 345个基因被注释,按照筛选差异基因的标准,高低油酸材料间,有2 043个基因表达量发生了改变,其中256个上调表达,53个下调表达(表1)。
2.2 蛋白质组与转录组关联分析
2.2.1 蛋白质组与转录组关联数量关系 将经鉴定的蛋白与转录组结果进行关联(当某一个蛋白质在转录组水平有表达量时,被认为两者相关联)。得到在鉴定、定量、显著差异3个范围中,能关联到的蛋白质和基因数量关系如表2。有2 604个基因在mRNA和蛋白质水平被鉴定,其中在mRNA和蛋白质水平均有表达的基因2 019个,而在334个显著差异表达蛋白中与基因关联的有87个。
2.2.2 蛋白质组与转录组的相关性 本研究中,依据mRNA水平和蛋白质水平的表达结果,从所有定量蛋白和基因的关联表达、蛋白质与基因变化趋势相同的关联表达与蛋白质和基因变化趋势相反的关联表达3个方面,分别对蛋白质组与转录组的相关性进行分析,结果如图1,定量蛋白和基因关联系数为-0.326 9;变化趋势相同差异蛋白和基因的表达关联系数为0.714 3;变化趋势相反差异蛋白和基因的表达关联系数为-0.735 8,蛋白质组和转录组相关性并不高。

表2 关联到的蛋白质组和转录组数量Tab.2 Number of associated proteomics and transcriptomics
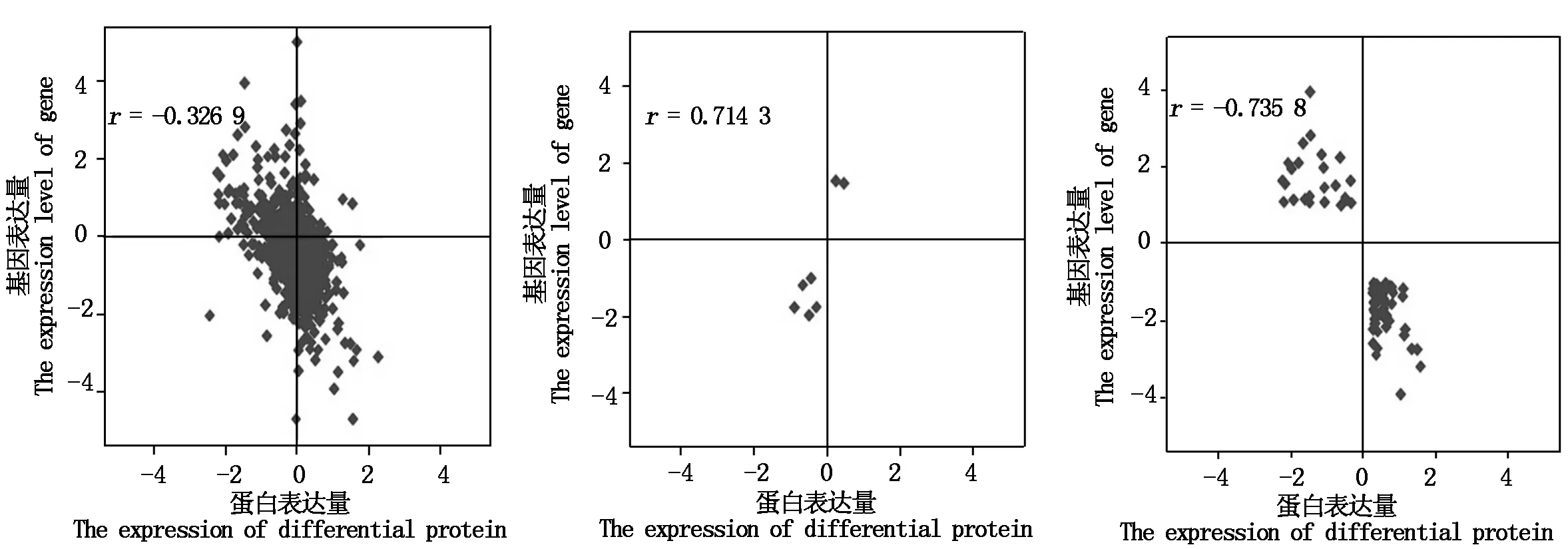
从左到右依次为:所有定量蛋白质和基因的关联表达;变化趋势相同的差异蛋白和基因的关联表达;变化趋势相反的差异蛋白和基因的关联表达。The pictures from left to right are the expression of all quantitative proteins and genes;The same expression of differential proteins and genes;The opposite expression of differential protein and gene.
2.2.3 脂肪酸代谢相关蛋白与基因的关联分析 与差异蛋白变化趋势相同的基因:本研究中,将转录组与蛋白质组数据关联分析,发现有80个与差异蛋白变化趋势相同的差异基因,其中有54个上调表达,26个下调表达。GO注释表明这些基因涉及代谢、信号转导、防御与胁迫应答、氧化还原、转录等方面功能。其中与脂肪酸代谢过程相关的有23个,16个上调表达,7个下调表达(表3)。
酰基辅酶A家族在脂肪酸合成过程发挥着相当重要的作用,Fae1基因编码控制3-酮酰-辅酶合成酶(KCS),是长链脂肪酸合成的限速酶,半胱氨酸是KCS催化中心不可或缺的氨基酸[15-16]。而酰基-ACP硫脂酶是脂肪酸合成酶(Fatty acid symthase,FAS)体系的组成部分[17],本研究关联到与脂肪酸合成相关的差异基因8个,其中上调表达6个,下调表达2个(表3),除了验证了前人所做的工作之外,gi|297330358(庚二酰ACP甲基酯的羧酸酯酶)、gi|297321940(磷酸化酶)、gi|297314818(乙酰胺、甲酰胺酶家族)3个推定蛋白有待于后续研究。
种子发育过程,蔗糖是合成脂肪酸的主要碳源,通过卡尔文循环转为丙酮酸,又氧化形成脂肪酸合成的前体物乙酰CoA[18]。植物体脂肪酸合成位点主要在质体,那么脂肪酸到细胞其他位点发挥作用必然有信号传递与运输的过程,溶血磷脂酰胆碱酰基转移酶是脂肪酸修饰作用的关键酶[19]。此外,能量代谢是脂肪酸代谢必不可少的一环,无论是脂肪酸的生物合成、分解以及运输过程都涉及能量的转化。本研究关联到的与糖代谢相关的基因有4个,均上调表达;关联到的与膜与转导有关的差异表达基因有4个,其中3个上调表达,1个下调表达;关联到7个与能量转化相关的差异表达基因,其中3个上调表达,4个下调表达(表3)。这些基因可能对脂肪酸整个代谢过程有重要意义。
蛋白显著差异表达,mRNA无显著变化:将转录组与蛋白质组数据关联分析,发现有247个在蛋白水平有表达变化而转录水平无显著变化的基因,其中92个上调表达,155个下调表达。与脂肪酸代谢过程相关的有25个,其中10个上调表达,15个下调表达(表4)。
mRNA显著差异表达,蛋白无显著变化:关联分析发现有231个在mRNA水平有表达变化,在蛋白水平无显著变化,其中106个上调表达,25个下调表达。与脂肪酸代谢过程相关的有24个,其中18个上调表达,6个下调表达(表5)。

表3 关联到与差异蛋白变化趋势相同的脂肪酸代谢相关的差异基因Tab.3 Differentially expressed genes associated with fatty acid metabolism
注:+.上调;-.下调。表4-5同。
Note:+.Up;-.Down.The same as Tab.4-5.

表4 蛋白显著差异表达,mRNA无显著变化的基因Tab.4 The differential genes with significantly change in protein express and indistinctively in mRNA

表5 mRNA显著差异表达,蛋白无显著变化Tab. 5 The differential genes with significantly change in mRNA express and indistinctively in protein
3 结论与讨论
通过对高低油酸材料进行蛋白质组与转录组分析,得到定量蛋白和基因关联系数为-0.326 9;变化趋势相同差异蛋白和基因的表达关联系数为0.714 3;变化趋势相反差异蛋白和基因的表达关联系数为-0.735 8,蛋白质组和转录组相关性并不高。
将转录组与蛋白质组数据关联分析,发现有80个与差异蛋白变化趋势相同的差异基因,GO注释表明这些基因涉及代谢、信号转导、防御与胁迫应答、氧化还原、转录等方面功能。其中与脂肪酸代谢过程相关的有23个,其中16个上调表达,7个下调表达。蛋白水平有表达变化而转录水平无显著变化的247个基因中,与脂肪酸代谢过程相关的有25个,其中10个上调表达,15个下调表达。在mRNA水平有表达变化而蛋白水平无显著变化的231个基因中,与脂肪酸代谢过程相关的有24个,其中18个上调表达,6个下调表达。
转录组学与蛋白质组学是后基因组学发展的热点领域,两者关联分析可以更全面的了解基因表达情况,通过相互间的补充与对比,从而得到更完整的生物信息,以便为后续研究做准备[20-21]。
但目前研究发现,两者之间相关性比较低[22]。李茂峰[23]采用基因芯片与iTRAQ技术,比较了纤维初始发育期4个纤维正常发育的陆地棉品种(系)之间的RNA和蛋白质表达差异,结果发现有240个基因可以找到270个对应的或高度同源的蛋白质,其中仅有7个基因表现一致,绝大多数不一致,相关系数仅为0.029。Kubala等[24]使用微阵列技术与2-DE双向凝胶电泳技术对PEG处理甘蓝型油菜种子萌发过程关键基因进行转录组与蛋白质组分析,差异表达蛋白78个,12个蛋白在转录和蛋白质组水平差异表达一致,相关系数也仅有0.15。本研究发现,高油酸材料定量蛋白和基因的表达呈现较低的负相关关系,可能是由于转录与翻译水平并不一致所致[22]。此外,差异基因与蛋白的表达趋势相反,呈负相关关系表明,转录后的翻译修饰等调控有重要作用[25]。
本研究利用蛋白质组与转录组关联分析,挖掘高油酸材料参与脂肪酸代谢的相关基因,发现有23个基因在蛋白质组与转录组均有显著差异,这些基因可能为解释高油酸高脂肪酸代谢机理做出解答,其中gi|297330358(庚二酰ACP甲基酯的羧酸酯酶)、gi|297321940(磷酸化酶)、gi|297314818(乙酰胺、甲酰胺酶家族)3个推定蛋白有待于后续研究。
[1] 刘后利. 油菜遗传育种学[M]. 北京:中国农业大学出版社,2000.
[2] 涂金星,张冬晓,张 毅,等. 我国油菜育种目标及品种审定问题的商榷[J]. 中国油料作物学报,2007,29(3):350-352.
[3] 高建芹,浦惠明,戚存扣,等. 高含油量油菜种子和果皮油分积累及主要脂肪酸的动态变化[J]. 中国油料作物学报,2009,31(2):173-179.
[4] Nicolosir J,Woolfrey B,Wilson T A,et al. Decreased aortic early atherosclerosis and associated risk factors in hypercholesterolemic hamsters fed a high-or mid-oleic acid oil compared to a high-linoleic acid oil[J]. The Journal of Nutritional Biochemistry,2004,15(9):540-547.
[5] Rudkowska I,Roynette C E,Nakhasi D K,et al. Phytosterols mixed with medium-chain triglycerides and high-oleic canola oil decrease plasma lipids in overweight men[J]. Metabolism-Clinical and Experimental,2006,55(3):391-395.
[6] 张振乾,胡庆一,官春云. 高油酸油菜研究现状、存在的问题及发展建议[J]. 作物研究,2016,30(4):462-474.
[7] 喻娟娟,戴绍军. 植物蛋白质组学研究若干重要进展[J]. 植物学报,2009,44(4):410-425.
[8] 张 杨,孙 明. iTRAQ技术在植物蛋白质组学中的应用[J]. 分子植物育种,2013,11(1):1047-1051.
[9] 陈 勇,柳亦松,曾建国. 植物基因组测序的研究进展[J]. 生命科学研究,2014,18(1):66-74.
[10] 许 波,张伟强,冯晓曦,等. 转录组测序技术在玉米中的应用研究进展[J]. 玉米科学,2014,22(1):67-72.
[11] Lan P,Li W F,Schmidt W. Complementary proteome and transcriptome profiling in phosphate-deficientArabidopsisroots reveals multiple levels of gene regulation[J]. Molecular & Cellular Proteomics,2012,11(11):1156-1166.
[12] Li M,Wang K,Li S,et al. Exploration of rice pistil responses during early post-pollination through a combined proteomic and transcriptomic analysis[J]. Journal of Proteomics,2016,131(131):214-226.
[13] Sinha R,Kumar D,Datta R,et al. Integrated transcriptomic and proteomic analysis ofArabidopsisthalianaexposed to glutathione unravels its role in plant defense[J]. Plant Cell Tissue and Organ Culture,2015,120(3):975-988.
[14] 张振亚,裴翠明,马 进.基于转录组和蛋白质组关联研究技术筛选紫花苜蓿耐盐相关候选基因[J]. 植物生理学报,2016(3):317-324.
[15] 李晓丹. 油料作物种子脂肪酸累积模式及相关基因的克隆与序列比较研究[D]. 北京:中国农业科学院,2007.
[16] Ohlrogge J,Browse J. Lipid biosynthesis[J]. The Plant Cell,1995,7(7):957.
[17] 王 倩. 拟南芥脂肪酸代谢相关基因的克隆与功能分析[D]. 杨凌:西北农林科技大学,2014.
[18] 王镜岩,朱圣庚,徐长法. 生物化学 (下册)[M].北京:高等教育教育出版社,2002.
[19] 官玲亮. 红花(CarthamustinctoriusL.)不同组织多不饱和脂肪酸积累模式及调控机制[D]. 雅安:四川农业大学,2011.
[20] Dyhrman S T,Jenkins B D,Rynearson T A,et al. The transcriptome and proteome of the diatomThalassiosirapseudonanareveal a diverse phosphorus stress response[J]. PLoS One,2012,7(3):1643-1646.
[21] Gonzalez L M G,ElKayal W,Ju C J T,et al. Integrated transcriptomic and proteomic profiling of white spruce stems during thetransition from active growth to dormancy [J].Plant Cell Environ,2012:35(4):682-701.
[22] 吴松峰,朱云平,贺福初.转录组与蛋白质组比较研究进展[J].生物化学与生物物理进展,2005,32(2):99-105.
[23] 李茂峰. 不同农艺性状棉花品种(系)纤维初始发育时期转录组和蛋白质组的比较分析[D].南京:南京农业大学,2011.
[24] Kubala S,Garnczarska M,Wojtyla,et al,Deciphering priming-induced improvement of rapeseed (BrassicanapusL.) germination through an integrated transcriptomic and proteomic approach[J].Plant Science,2015,231:94-113.
[25] Galland M,Huguet R,Arc E,et al.Dynamic proteomics emphasizes the importance of selective mRNA translation and protein turnover duringArabidopsisseed germination[J]. Mol Cell Proteomics,2014,13 (1):252-268.
猜你喜欢
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2021年4期)2021-07-23
智慧健康(2021年33期)2021-03-16
中国生殖健康(2020年5期)2021-01-18
新农业(2020年18期)2021-01-07
中成药(2018年12期)2018-12-29
中国生殖健康(2018年5期)2018-11-06
天然产物研究与开发(2018年2期)2018-04-04
中国民族医药杂志(2016年4期)2016-05-09
中国洗涤用品工业(2015年8期)2015-02-28
