
太湖食物网生态化学计量学特征空间差异
2018-01-09 07:18:06徐德琳林乃峰邹长新安树青环境保护部南京环境科学研究所江苏南京004南京大学生命科学院江苏南京003
中国环境科学 2017年12期
徐德琳 ,林乃峰 ,邹长新 ,王 燕 ,吴 丹 ,蔡 颖 ,安树青 ∗ (.环境保护部南京环境科学研究所,江苏南京 004;.南京大学生命科学院,江苏 南京 003)
食物网被认为是包含了生态系统中所有营养关系的网络结构[1],其内部的物质循环和能量流动过程一直是生态学研究的重点问题[2].而生态化学计量学,综合了生物学、化学和物理学的基本原理,对生态作用中的多种化学物质平衡关系进行研究,能够将食物网内不同营养级间生产者和消费者的物质变动连接起来[3].碳(C)、氮(N)和磷(P)是在生态化学计量学中被研究的最为广泛的 3种元素,其含量及比例特征能够反映有机体的生长,种群的动态,并最终反映到食物网结构[4],同时,食物网物种组成和营养级结构的改变也会引起C、N、P化学计量特征的改变.
生态化学计量能够反映生态系统的组分、相互关系以及过程[5].生物在长期的进化过程中,形成了特定的内稳态机制,即在外界变化的环境干扰中,具有保持自身化学组成相对恒定的能力[6].化学物质的内稳态是生态化学计量的核心与前提,这意味着高营养级的生物体体内物质组成比例通常比低营养级的生物要稳定.当其猎物的元素组成不能满足其捕食者的需求时,元素再循环的速率将降低[7].因此,食物网内生物的元素含量能够反映元素循环情况.元素的不平衡同样可以影响消费者的生长速率.当其猎物发生磷限制时,消费者的生长速率将受限,因为没有足够的磷元素产生蛋白质合成所需的 RNA[8].这种化学计量的不匹配同样影响其繁殖、种间竞争,并最终影响食物网的运行.
生态化学计量学的研究在国内发展迅速,主要集中在区域C、N、P化学计量特征及驱动因素方面,并以陆地的森林生态系统和草原生态系统为主[9-10].但是,随着水环境问题的不断凸显,在水生态系统开展生态化学计量学研究越来越得到重视[11].在富营养化湖泊中,人类活动显著改变了营养物质的输入和比例,并不可避免地改变了湖泊生态系统的健康.多种来源的污染物导致 N和 P含量增加,并致使湖泊食物网内营养盐元素比例的不平衡[12].已有研究表明水体富营养化可能导致食物网内生产者和消费者间的生态化学计量关系发生改变,但目前的研究主要集中在浮游或底栖食物网上[13-14],对湖泊食物网的生态化学计量特征的整体性研究较为缺乏.
对大型浅水湖泊食物网生态化学计量学特征进行研究十分必要,因为其空间异质性更高,湖水与沉积物间物质交换强烈,对污染物相应更为敏感,也更易受到外界干扰[15].在一些大型浅水湖泊中,可能会出现 2种稳态共存:水体清澈且水草密集的“草型稳态”,以及水体浑浊且浮游藻类占优势的“藻型稳态”[16].作为一个典型的大型浅水湖泊,太湖具有草藻 2种稳态共存:东南部的草型湖区和西北部的藻型湖区
[17].因此,我们利用元素分析方法对太湖不同富营养状态湖区内的食物网生态化学计量学特征的空间异质性进行分析,以揭示水体富营养化对食物网生态化学计量特征的影响.本研究致力于分析与比较不同湖区食物网组分元素含量(TOC,TN和TP)的空间异质性、食物网组分C、N、P元素间耦合关系(C:N,C:P和N:P)的空间异质性及食物网内C:N:P生态化学计量学特征的不平衡性.
1 研究区概况与研究方法
1.1 研究区概况
太湖位于长江中下游,是我国第三大淡水湖,具有重要的供水、航运、渔业、防洪、文化等服务功能.湖泊面积 2,428km2,全湖平均水深 1.9m,东太湖平均水深0.9m[18].作为典型的大型浅水湖泊,受水文地质、营养负荷等因素综合影响,太湖内部存在草藻 2种湖区共存:西北部湖区为藻型湖区,从上世纪 80年代后期开始,富营养化程度加剧,夏季时蓝藻水华爆发严重;而东南部湖区为草型湖区,水草生长茂盛[19].
1.2 样品采集与处理
所有的样品于2010年9月于太湖的6个采样点采集完成(见图1).样点1、2、3位于藻型湖区,样点4、5、6位于草型湖区,每个样点均有 3个重复,分别采集环境样品(包含水样和沉积物样),生物样品(包含底泥微生物、浮游植物、浮游动物、鱼类和沉水植物).
太湖水体较浅,故水样采集采用混合水柱.现场用有机玻璃采水器采集水面下30cm和50cm的水柱,混合后装入聚丙烯采样瓶中.浮游植物和动物分别用25号和13号浮游生物网采集,装入聚丙烯采样瓶中.用彼得森采泥器采集表层0~30cm 沉积物,分为 2部分,一部分用于沉积物C、N、P含量测试,一部分用于沉积物微生物C、N、P含量测试,分别放入密封袋中.用彼得森采泥器再次采集底栖动物(斧足类和腹足类),泥样经0.5mm套筛冲洗,在清水中排泄 24h,以去除肠含物影响,去壳取肌肉部分放入密封袋中.沉水植物用竹竿绞起后挑选鲜活植株茎叶,洗净,为避免腐烂装入布袋中保存.藻型湖区的鱼类由孔径1mm的拖网捕获,草型湖区的鱼类由湖区内的簖网捕获.对于大型鱼类(个体长度>10cm),取其背部肌肉作为样品,对于小型鱼类(个体长度<10cm),在去除内脏后由多条个体合并为一个样品.虾类伴随鱼类一同采集,去壳去头放入密封袋中.所有样品冷藏带回实验室立即处理.测定得出,水样的溶解性总碳(DTC)和底泥微生物的总有机碳(TOC)由总有机碳分析仪(vario TOC, Elementar Company, Germany)测定得出.沉积物样品送中国科学院南京土壤研究所检测其TN、TP和有机质含量.生物样品(浮游植物、浮游动物、底栖动物、鱼类和沉水植物)的TOC和TN 由 CHN-元素分析仪(Vario EL cube,Elementar Company, Germany)经 1800℃高温燃烧气化测定得出.所有生物样品的 TP由ICP-AES电感耦合等离子体发射光谱仪(Optima 5300DV, PE Company, USA)测定得出.
1.4 数据处理
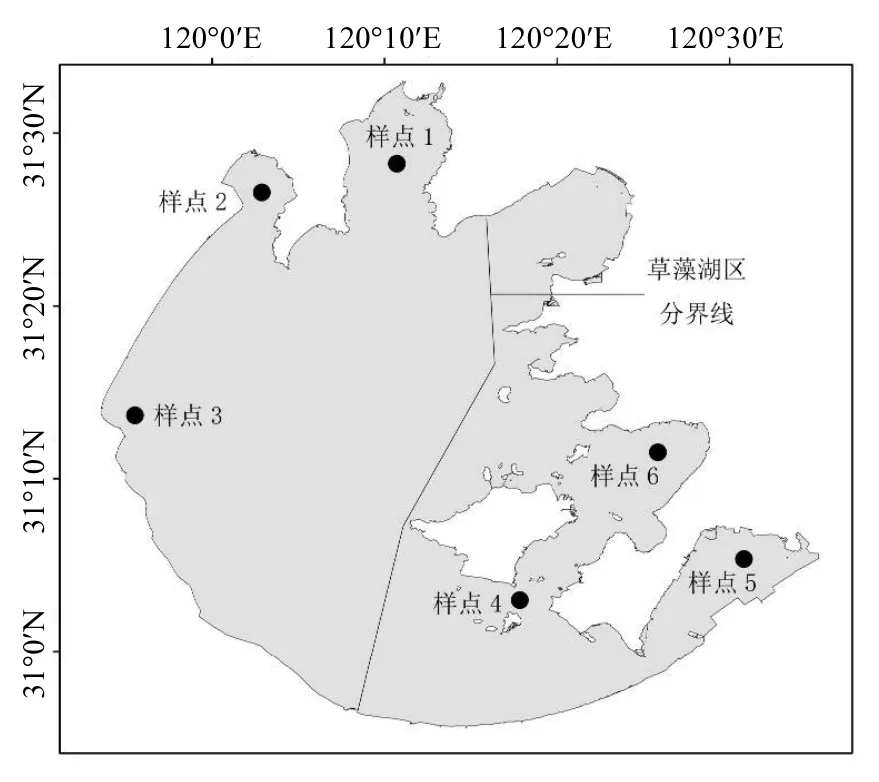
图1 样点分布示意Fig.1 Location of sampling sites
同一湖区内部及不同湖区间的环境样品和生物样品的元素含量(TOC、TN、TN),及元素比例(C:N、N:P)进行 ANOVAs分析.不能通过方差齐性检验的数据,进行 Mann-Whitney非参数检验.统计显著性α = 0.05.所有数据均在Origin 8.5进行统计分析.
2 结果
2.1 环境与生物样品的元素含量
为去除水体中颗粒悬浮物对水体C、N、P元素测定的影响,水样在测定之前,经 0.45µm醋酸纤维滤膜过滤备用.沉积物样品经酸化、风干及研磨后,用 100目网筛过筛.浮游植物样品通过离心法分离提纯,浮游动物样品通过体视显微镜分离后离心提纯.所有生物样品均经冻干后研磨成粉.底栖微生物由新鲜沉积物用氯仿熏制.
1.3 元素分析
水样的溶解性总氮(DTN)、溶解性总磷(DTP)和底泥微生物的总氮(TN)、总磷(TP)含量由连续流动分析仪(San++, Skala Company, Netherlands)
太湖藻型湖区环境样品(水样和沉积物样)具有相对较高的 C、N、P元素含量(表1和图2).ANOVA结果显示,太湖草藻湖区水样的DTC、DTN和DTP均具有显著性差异,但是草藻湖区沉积物样仅TP显示出显著性差异,而 TOC和TN均未显示出显著性差异(表2).
与环境样品相比,太湖草藻湖区生物样品元素含量(TOC,TN和 TP)的空间异质性较低(表2和图3).草藻湖区内高营养级功能组团在元素组成方面具有较高的稳定性,而低营养级功能组团的空间异质性相对更高.分析结果显示,草藻湖区底泥微生物的TOC,TN和TP均具有显著差异;草藻湖区浮游植物和浮游动物的TOC和TN具有显著性差异,但 TP无显著性差异;草藻湖区底栖动物的TOC和TN具有显著性差异,但是其TP无显著性差异;草藻湖区虾仅TOC具有显著性差异;而两个湖区的鱼类(包括滤食性鱼类、杂食性鱼类与肉食性鱼类)的TOC,TN和TP均无显著差异(表2).

表1 太湖草藻湖区环境样品与生物样品TOC、TN和TP含量均值Table 1 TOC, TN and TP (mean, with SD followed in parentheses) of the basal sources and consumers in the algae- and macrophyte-dominated zones of Lake Taihu
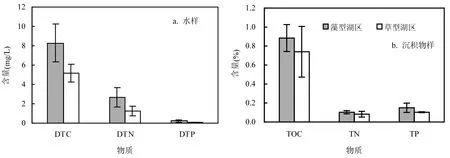
图2 太湖草藻湖区水样DTC、DTN和DTP含量(a)和沉积物样品TOC、TN和TP含量(b)Fig.2 The DTC DTN and DTP contents of water samples (a) and TOC, TN and TP content of sediment samples (b) in the algae- and macrophyte-dominated zones of Lake Taihu
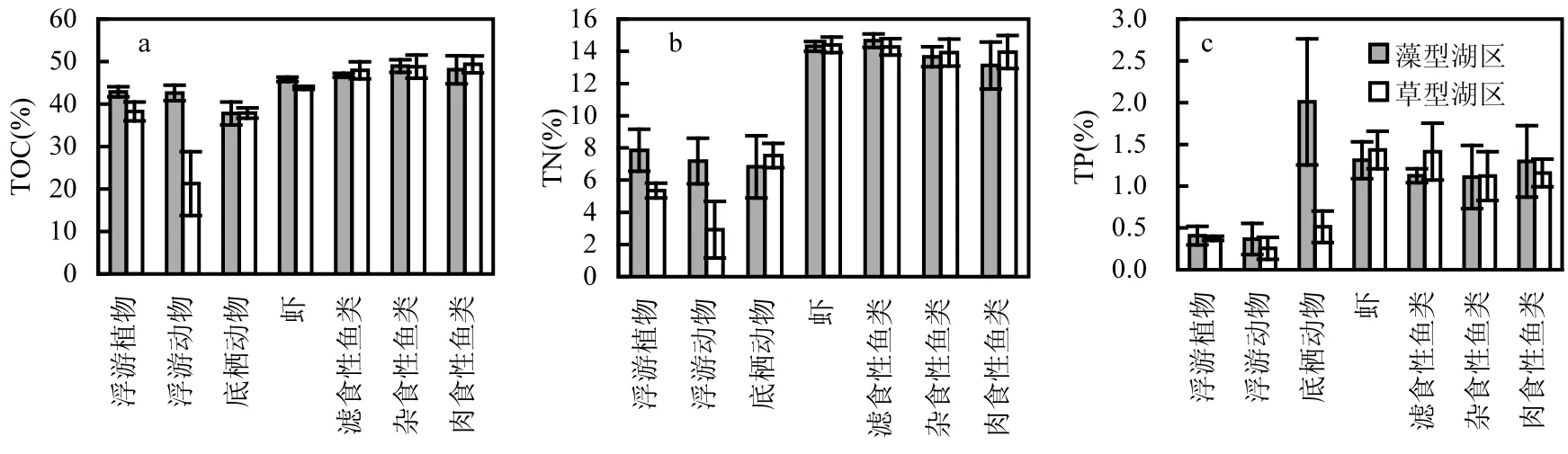
图3 太湖草藻湖区生物样品TOC(a)、TN(b)和TP(c)含量Fig.3 The TOC (a), TN (b) and TP (c) contents of biological samples in the algae-dominated zone and the macrophyte-dominated zone of Lake Taihu.

表2 太湖草藻湖区环境样品与生物样品TOC、TN和TP含量ANOVA分析结果Table 2 ANOVA results comparing differences of TOC, TN and TP contents in environmental and biological samples between the algae- and macrophyte-dominated lake zones of Lake Taihu
2.2 环境与生物样品的元素比例
藻型湖区环境样品的C:N,C:P和N:P值较草型湖区低,但是这种空间异质性并不非常明显(表3和图4).差异性分析结果显示,2个湖区的环境样品中,仅有水样的C:P和沉积物的C:N具有显著性差异(表3).水样的C:P比C:N和N:P值都高,而沉积物样的C:N比C:P和N:P的值高(图4).
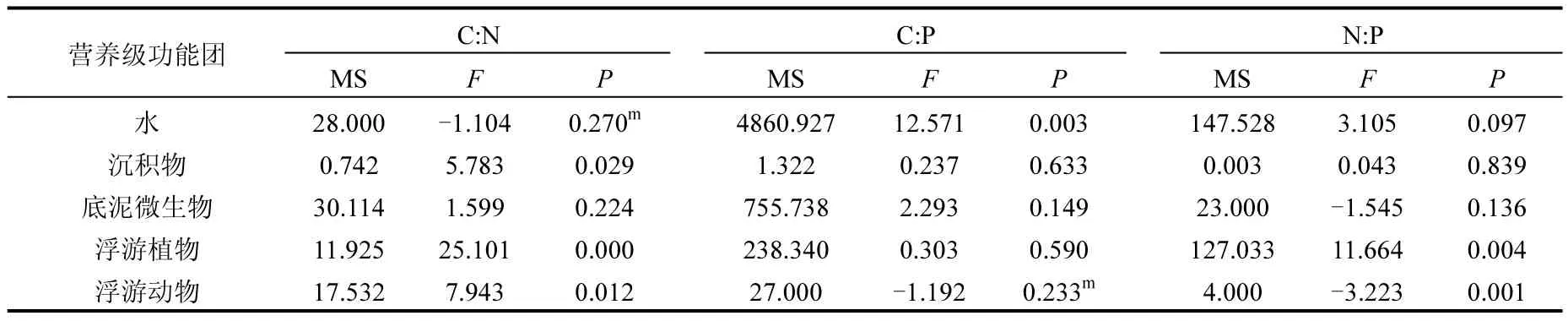
表3 太湖草藻湖区环境样品与生物样品C:N、C:P和N:P比值ANOVA分析结果Table 3 ANOVA results comparing differences of C:N, C: P and N:P ratios in environmental and biological samples between the algae- and macrophyte-dominated lake zones of Lake Taihu

续表3
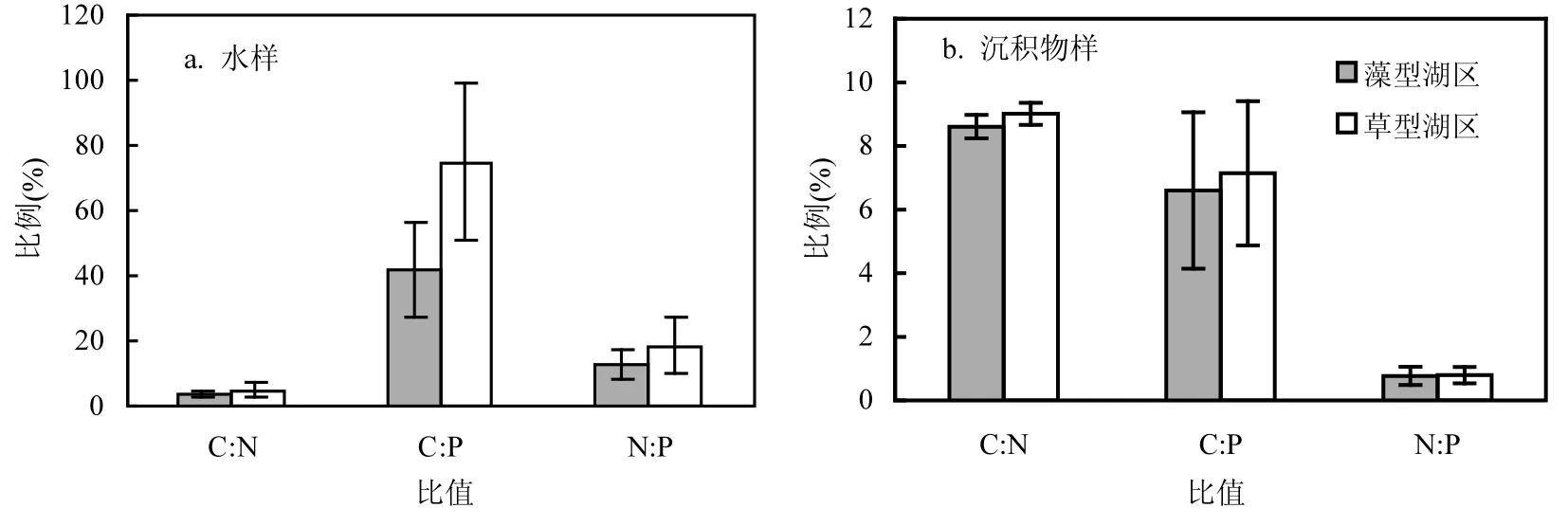
图4 太湖藻型和草型湖区水样C:N、C:P和N:P比值(a)和沉积物样C:N、C:P和N:P比值(b)Fig.4 The C:N, C:P and N:P ratios of water samples (a) and sediment samples (b) in the algae-dominated zone and the macrophyte-dominated zone
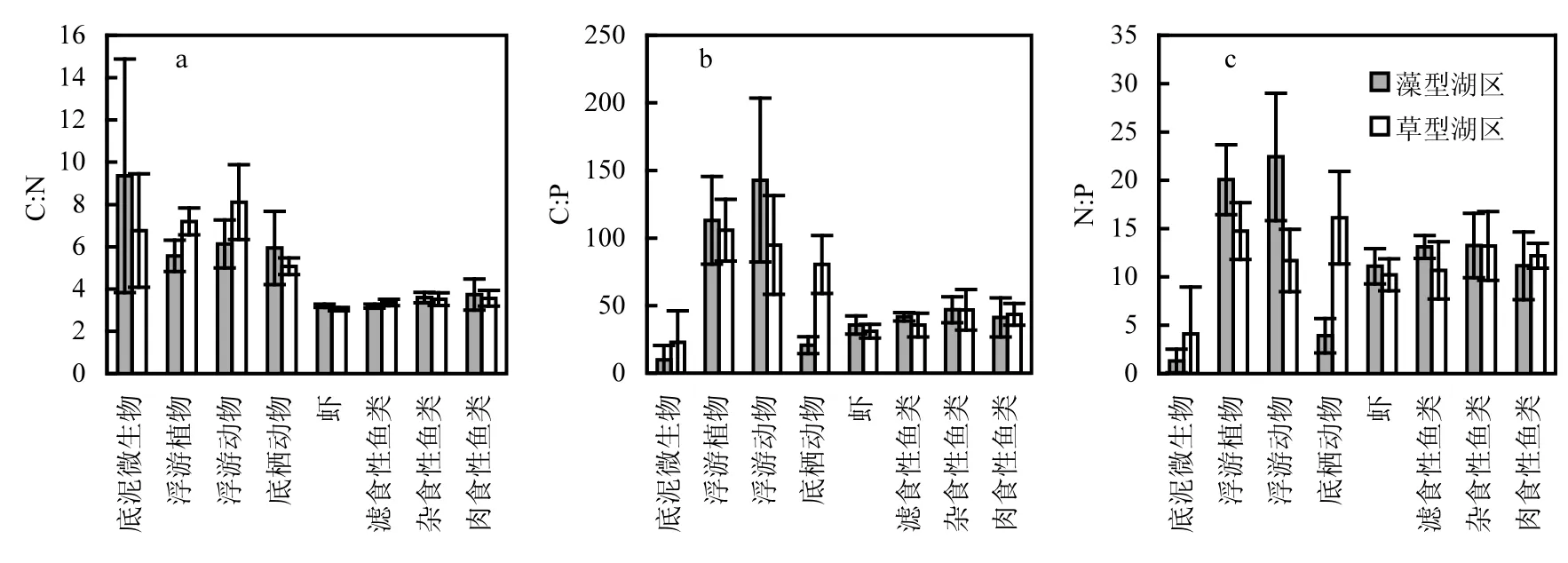
图5 太湖藻型和草型湖区生物样品C:N(a)、C:P(b)和N:P(c)比值Fig.5 The C:N (a), C:P (b) and N:P (c) ratios of biological samples in the algae-dominated zone and the macrophyte-dominated zone of Lake Taihu
两个湖区生物样品元素比例的空间异质性相对复杂(表3和图5).低营养级的功能团在元素组成上较高营养级的功能团具有更高的变动性.差异性分析结果显示,仅有浮游植物的 C:N 和N:P、浮游动物的 C:N 和 N:P、底栖动物的 C:P和 N:P具有显著性差异,其他生物样品的元素比例均为显示出显著性差异.
2.3 食物网生态化学计量特征
如图6所示,藻型湖区水样的C:P和N:P均比草型湖区的低,但是藻型湖区浮游生物(浮游植物和浮游动物)的C:P和N:P较草型湖区的高.在藻型湖区,浮游动物的C:P和N:P比浮游植物的高.藻型湖区的沉积物和底栖生物(底泥微生物和底栖动物)的 C:P和 N:P较草型湖区的低.在两个湖区的游泳组团(虾类、滤食性鱼类、杂食性鱼类和肉食性鱼类)中,杂食性鱼类均显示出最高的C:P和N:P.除了肉食性鱼类外,藻型湖区游泳组团的C:P和N:P均比草型湖区的高.

图6 太湖草型湖区(a)与藻型湖区(b)食物网生态化学计量比值(C/N/P)Fig.6 The stoichiometric characteristics ratios (C/N/P) of food webs in the algae-dominated zone (a) and the macrophyte-dominated zone (b) of Lake Taihu
3 讨论
太湖草藻湖区环境样品(水样和沉积物样)的元素含量(C,N和P)显示出了较高的空间异质性.总体而言,藻型湖区环境样品的C、N、P含量较草型湖区的高,这可能与藻型湖区环湖的污染物输入量较高有关[20].研究表明,藻型湖区沉积物中 P组分以无机磷(IP)为主,外源输入是无机磷的重要来源;而草型湖区沉积物中的P组分以水生植物降解的有机磷(OP)为主[21].藻型湖区位于太湖北部重工业污染区和重要农产品区,同时有多条入湖河流将陆源的无机磷带入湖体,造成环境样品营养盐含量高[22-23].与草藻湖区水样的元素含量(C,N和P)差异显著相比,2个湖区沉积物样仅有TP显示出显著性差异,这可能与P元素的相对稳定性以及P循环的特征有关[24-25].由于此次采样时间为 9月,藻型湖区水体藻类尤其是蓝藻大量繁殖,沉积物表面有大量的藻类死亡堆积,湖区易形成厌氧环境,利于反硝化作用,故环境样品的TN含量会较其他季节降低,而TP含量相对高[26-27].因此,季节因素对草藻湖区环境样品的元素含量空间异质性也会带来一定的影响.
对于生物样品而言,低营养级物种元素含量上(C,N和P)比高营养级物种体现出较高的空间差异性.低营养级物种(底泥微生物、浮游植物、浮游动物)的元素含量对环境变化较高营养级物种更为敏感[13].草、藻湖区的虾类和鱼类的元素含量差异性不大,这与高营养级生物生态化学计量的内稳态特征相吻合[28].保持自身营养元素的稳态是生命体的本质特征,目前认为维持化学计量的内稳态有助于物种在面临短暂的外界压力后迅速恢复,但也有研究指出弱稳态物种更适应环境条件的变化,因其可以贮存多余的营养物质,以备在资源匮乏时使用,进而保证生长速率[29].
本研究太湖表层沉积物TN和TP含量与于佳佳等[30]研究结果基本一致,但是与王健等[31]调查的中国东部100个浅水湖泊的表层沉积物TN(0.0479%~0.5574%)和 TP(0.0248%~0.1000%)的范围含量相比,太湖藻型湖区表层沉积物 TP的含量偏高,体现出较高的内源污染负荷.藻型湖区底栖动物和底泥微生物的TP含量均较草型湖区的高,这与沉积物 TP含量的分布一致.湖泊水环境氮素和磷素在化学计量上的变化,与湖泊富营养化的空间分布特性存在密切关系[32-33].
与草型湖区相比,藻型湖区环境样品的C:N、C:P、N:P都相对较低.C:N是有机质降解速率的一个重要指标,通常与有机质降解率成反比[34].在藻型湖区中,沉积物的低C:N显示了该区域可能较高的有机质降解速率,而浮游植物较低的C:N与藻型湖区浮游植物中蓝藻占比较大有关[35].藻型湖区浮游植物的低C:N也通过食物链传递,降低了浮游动物的C:N[36].藻型湖区底栖动物较低的 C:P和 N:P可能与该区域沉积物的 P含量较高有关[37].2个湖区高营养级生物显示出较为稳定的元素比例,这也反映了化学计量的内稳态特征.
尽管本研究中食物网的功能团分类并不是十分精细,但在一定程度上展示了太湖的生态系统空间异质性.草型湖区和藻型湖区所受的不同的人类影响和生态系统特征,大大影响了这 2个湖区食物网的生态化学计量特征.生态化学计量特征也对食物网结构和湖泊生物化学循环产生重要影响[11].本研究静态展现了太湖生态系统元素组成和比例,下一步研究将探索利用元素循环模型、元素添加实验、稳定同位素分析方法来更加系统地研究食物网生态化学计量特征的空间异质性.此外,食物网的生态化学计量特征研究成果可为湖泊生态系统管理提供理论支撑,因为单一元素的控制策略可能对于水生生态系统达不到预期效果[38].
4 结论
4.1 与草型湖区相比,太湖藻型湖区环境样品(水样和沉积物样)具有相对较高的 C、N、P元素含量,除沉积物 TOC和 TN外,其余环境样品C、N、P元素含量有显著性差异.受营养盐含量较高影响,太湖藻型湖区环境样品的 C:N,C:P和N:P值较草型湖区的低,但两湖区间环境样品C、N、P元素比例的空间异质性不高.
4.2 太湖草藻湖区的食物网内低营养级物种的C、N、P元素含量和比例显示出较大的空间差异,而高营养级物种并未受到食物源元素含量和比例差异的太大影响,能够更好地维持自身的C、N、P元素含量内稳态.
4.3 草、藻湖区食物网均存在营养级间化学计量不平衡现象,食物网组分的C、N、P含量和比例的空间分布特征,反映出草藻湖区生态系统不同层次组分的生态过程本身和抵抗外界干扰能力的差异,低营养级物种更易受到外界环境变化的影响.
[1] 张 波,吴 强,金显仕.1959-2011年莱州湾渔业资源群落食物网结构的变化 [J]. 中国水产科学, 2015,22(2):278-287.
[2] Banasek-Richter C, Bersier L F, Cattin M F, et al. Complexity in quantitative food webs [J]. Ecology, 2009,90(6):1470-1477.
[3] 贺金生,韩兴国.生态化学计量学:探索从个体到生态系统的统一化理论 [J]. 植物生态学报, 2010,34(1):2-6.
[4] 邬 畏,何兴东,周启星.生态系统氮磷比化学计量特征研究进展 [J]. 中国沙漠, 2010,30(2):296-302.
[5] Bradshaw C, Kautsky U, Kumblad L. Ecological stoichiometry and multi-element transfer in a coastal ecosystem [J]. Ecosystems,2012,15(4):591-603.
[6] Sterner R W, Elser J J. Ecological Stoichiometry: the Biology of Elements from Molecules to the Biosphere [M]. Princeton,Princeton University Press, 2002:221-224.
[7] Persson J, Fink P, Goto A, et al. To be or not to be what you eat:regulation of stoichiometric homeostasis among autotrophs and heterotrophs [J]. Oikos, 2010,119(5):741-751.
[8] 程 滨,赵永军,张文广,等.生态化学计量学研究进展 [J]. 生态学报, 2010,30(6):1628-1637.
[9] 曾冬萍,蒋利玲,曾从盛,等.生态化学计量学特征及其应用研究进展 [J]. 生态学报, 2013,33(18):5484-5492.
[10] 谢 锦,常顺利,张毓涛,等.天山北坡植物土壤生态化学计量特征的垂直地带性 [J]. 生态学报, 2016,36(14):4363-4372.
[11] 邢 伟,刘 寒,刘贵华.生态化学计量学在水生态系统中的研究与应用 [J]. 植物科学学报, 2015,33(5):608-619.
[12] Smith V H, Schindler D W. Eutrophication science: where do we go from here? [J]. Trends in Ecology & Evolution, 2009,24(4):201-207.
[13] 陈 蕾,李超伦.海洋浮游生物的生态化学计量学研究进展 [J].应用生态学报, 2014,25(10):3047-3055.
[14] 蔡永久,薛庆举,陆永军,等.长江中下游浅水湖泊 5种常见底栖动物碳、氮、磷化学计量特征 [J]. 湖泊科学, 2015,27(1):76-85.
[15] Rawcliffe R, Sayer C D, Woodward G, et al. Back to the future:using palaeolimnology to infer long-term changes in shallow lake food webs [J]. Freshwater Biology, 2010,55(3):600-613.
[16] McCann M J. Evidence of alternative states in freshwater lakes:A spatially-explicit model of submerged and floating plants [J].Ecological Modelling, 2016,337(c):298-309.
[17] Zhao D H, Jiang H, Cai Y, et al. Artificial regulation of water level and its effect on aquatic macrophyte distribution in Taihu Lake [J]. PLoS ONE, 2012,7(9):e44836.
[18] 王银平,谷孝鸿,曾庆飞,等.太湖不同生态型湖区湖鲚(Coilia ectenes taihuensis)食物组成及其季节变化 [J]. 湖泊科学,2016,28(5):1078-1085.
[19] 陈立侨,刘 影,杨再福,等.太湖生态系统的演变与可持续发展[J]. 华东师范大学学报(自然科学版), 2003,4:99-106.
[20] 王 倩,吴亚东,丁庆玲,等.西太湖入湖河流水系污染时空分异特征及解析 [J]. 中国环境科学, 2017,37(7):2699-2707.
[21] 章婷曦,王晓蓉,金相灿.太湖不同营养水平湖区沉积物中磷形态的分布特征 [J]. 农业环境科学学报, 2007,26(4):1207-1213.
[22] 赵海霞,王 梅,段学军.水环境容量约束下的太湖流域产业集聚空间优化 [J]. 中国环境科学, 2012,32(8):1530-1536.
[23] 王 佩,卢少勇,王殿武,等.太湖湖滨带底泥氮、磷、有机质分布与污染评价 [J]. 中国环境科学, 2012,32(4):703-709.
[24] Liu W Z, Zhang Q F, Liu G H. Effects of Watershed Land Use and Lake Morphometry on the Trophic State of Chinese Lakes:Implications for Eutrophication Control [J]. Clean-soil, air,water, 2013,39(1):35-42.
[25] 王永平,朱广伟,洪大林,等.太湖草、藻型湖区沉积物-水界面厚度及环境效应研究 [J]. 中国环境科学, 2013,33(1):132-137.
[26] 王秋娟,李永峰,姜 霞,等.太湖北部三个湖区各形态氮的空间分布特征 [J]. 中国环境科学, 2010,30(11):1537-1542.
[27] 周 莉,冯 胜,李忠玉,等.夏季太湖浊度分布特征及其在水-沉积物界面识别中的应用 [J]. 中国环境科学, 2015,35(10):3108-3116.
[28] 王 芳,国先涛,董双林.水域生态系统生态化学计量学研究进展 [J]. 中国海洋大学学报(自然科学版), 2015,45(12):16-23.
[29] Persson J, Fink P, Goto A, et al. To be or not to be what you eat:regulation of stoichiometric homeostasis among autotrophs and heterotrophs [J]. Oikos, 2010,119(5):741-751.
[30] 于佳佳,尹洪斌,高永年,等.太湖流域沉积物营养盐和重金属污染特征研究 [J]. 中国环境科学, 2017,37(6):2287-2294.
[31] 王 健,张靖天,昝逢宇,等.中国东部浅水湖泊沉积物总氮总磷基准阈值研究 [J]. 生态环境学报, 2014,23(6):992-999.
[32] 蔡龙炎,李 颖,郑子航.我国湖泊系统氮磷时空变化及对富营养化影响研究 [J]. 地球与环境, 2010,38(2):235-241.
[33] 张明礼,杨 浩,林振山,等.太湖竺山湾底泥中有害物质含量与环境污染评价 [J]. 中国环境科学, 2011,31(5):852-857.
[34] 王 娜,徐德琳,郭 璇,等.太湖沉积物微生物生物量及其与碳、氮、磷的相关性 [J]. 应用生态学报, 2012,23(7):1921-1926.
[35] 周笑白,张宁红,张 咏,等.太湖蓝藻的时空变化规律及治理方法 [J]. 生态环境学报, 2013,22(12):1930-1935.
[36] Elser J J, Acharya K, Kyle M, et al. Growth rate-stoichiometry couplings in diverse biota [J]. Ecology Letters, 2003,6(10):936-943.
[37] Liu E, Shen J, Yuan H, et al. The spatio-temporal variations of sedimentary phosphorus in Taihu Lake and the implications for internal loading change and recent eutrophication [J].Hydrobiologia, 2013,711(1):87-98.
[38] Glibert P M. Ecological stoichiometry and its implications for aquatic ecosystem sustainability [J]. Current Opinion in Environmental Sustainability, 2012,4(3):272-277.
猜你喜欢
海洋通报(2022年2期)2022-06-30 06:07:04
海洋石油(2021年3期)2021-11-05 07:43:12
当代水产(2021年8期)2021-11-04 08:49:06
河北环境工程学院学报(2021年1期)2021-03-19 08:43:00
海峡旅游(2018年4期)2018-06-01 11:20:00
江西建材(2018年4期)2018-04-10 12:37:24
玩具世界(2017年4期)2017-07-21 13:27:24
宝藏(2017年2期)2017-03-20 13:16:42
中亚信息(2016年3期)2016-12-01 06:08:24
环境科技(2015年4期)2015-11-08 11:10:44
