
3种楠属植物2年生苗生长规律及光合生理特性
2018-01-06 01:11马永征何舒怀唐巍王东费永俊
江苏农业科学 2017年22期
马永征+何舒怀+唐巍+王东+费永俊

摘要: 以闽楠[Phoebe bournei (Hemsl.) Yang]、浙江楠(Phoebe chekiangensis C.B.Shang)、桢楠(Phoebe zhennan)3种楠属植物2年生幼苗作为试材进行移栽,并测量其苗高、基径、春夏秋梢生长量、叶数和分枝数等生长指标,测定了其净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)等光合指标,以及可溶性蛋白、超氧化物歧化酶(SOD)、丙二醛(MDA)等生理指标。结果表明,桢楠的生长和生理指标都优于浙江楠、闽楠,桢楠生长状况最好,对环境的适应能力较强。
关键词: 楠属植物;生长规律;光合特性;生理特性
中图分类号: Q945.32;S792.240.5 文献标志码: A
文章编号:1002-1302(2017)22-0122-04
楠木为中国及南亚特有的珍贵树种,它包括樟科(Lauraceae)润楠属(Machilus Nees)和楠属(Phoebe Nees)植物。因其在家具制作、木雕工艺及园林绿化等方面具有广泛的应用价值和经济价值,楠木资源遭到了严重的掠夺,天然林资源濒临枯竭。楠属植物中桢楠(Phoebe zhennan)、浙江楠(Phoebe chekiangensis C.B.Shang)和闽楠[Phoebe bournei(Hemsl.) Yang]均为国家Ⅱ级重点保护野生植物 。现存的楠木资源多作为古树名木或风水林被保存,散生于村边或祠庙旁[3-4],因此,在学术界掀起了楠木研究热潮。范剑明等全面研究了楠木播种育苗技术[5-7];朱雁阐述了楠木容器苗芽苗移栽技术[8];王艺等研究了不同栽培基质对楠木根系发育的影响[9];钟灼坤指出,播种密度、苗期追肥以及病虫害的防治与楠木高效快速育苗具有相关性[10];刘刚等则对楠木进行了胁迫试验,研究其相应的抗逆生理特性[11-12]。本研究以桢楠、浙江楠、闽楠2年生苗为研究对象,系统研究楠木幼苗的生长规律、光合和生理指标,以期明确楠木的生长习性及应激反应,对楠木生产与珍稀树种的保育都具有重要的实践指导意义。
1 材料与方法
1.1 用地概况及试验材料
试材移栽后置于湖北省荆州市长江大学盆景园内,位于30°21′16″N、112°08′31″E,地处亚热带季风气候区,四季分明,热量丰富,光照适宜,年平均温度15.9~16.6 ℃,年降水量在1 100~1 300 mm,年无霜期250~267 d,太阳年总辐射约 435~460 kJ/cm2,平均年日照时数为1 800~2 000 h[13]。
2016年3月,从盆景园中选取生长状况一致的桢楠、浙江楠、闽楠2年生苗各60株(分别编号ZN 1~60号,ZJN 1~60号,MN 1~60号),移栽于聚乙烯塑料花盆中(上口径 15 cm,下口径12 cm,高13 cm),每盆栽种1株,统一采用黑土和木屑(3 ∶ 1)的混合基质,基质提前24 h用800倍多菌灵液消毒处理。
1.2 试验方法
1.2.1 育苗管理
栽种后2个月内将所有试验苗进行遮阴处理,以保证其成活率。育苗期间进行合理的除草、常规浇水及适当的病虫害防治等措施,保证所有试验苗正常生长;于2016年8—9月用含腐殖酸50%的肥料对部分苗木进行多次追肥处理。夏季高温时适当遮阴处理。
1.2.2 测定指标
标号1~30的楠木用于测量株高、基径等生长指标,研究3种楠木的生长特性;对标号31~60的楠木进行追肥处理,并测定光合及生理指标,研究3种楠木的光合及生理特性。
1.2.2.1 生长指标
2016年3—11月,每月进行3次抽样测量,每次抽样5株,3次重复;使用游标卡尺(精度 0.02 mm)和直尺(精度1 mm)测量基径、株高,并记录叶片数和分枝数。要求基径测量植株茎与土壤接触处的直径;株高测量植株顶梢处到土壤表面的垂直距离;分枝数计数从植株茎上发出的一级分枝。
1.2.2.2 光合指标
2016年8月16日09:00—11:00(天气晴朗情况下),采用美国Li-6400便携式光合分析仪对植株进行测定,每个处理测3次重复,随机抽取3株,每株检测3张叶片,每次测定都选取植株冠层中部的成熟功能叶片的同一位置。待数据稳定后,记录其净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)指标值[14]。
1.2.2.3 叶绿素含量
2016年9月,采用SPAD-502Plus便携式叶绿素测定仪对植株进行测定。每个处理测3次重复,记录其叶绿素相对含量。
1.2.2.4 生理指标
2016年11月,至试验苗停止生长后,分别从3种楠木中随机采集足够量的当年生叶片,用透明自封袋袋装,贴上标签,用冰盒带回实验室,于超低温冰箱中保存,进行试验操作,测量其可溶性糖含量、可溶性蛋白含量、丙二醛含量、超氧化物歧化酶活性以及根活性,均進行3次重复。
1.3 数据处理
利用SAS8.0统计软件对观测数据进行差异性分析,图表制作和数据计算均采用Excel 2007软件完成。
2 结果与分析
2.1 幼苗生长特性
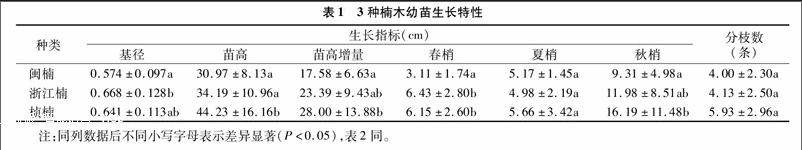
闽楠、浙江楠、桢楠幼苗移栽后的生长状况存在差异(表1),但都具特有的生长规律。(1)桢楠生长状况好且生长速率快,苗高、基径增长迅速;新生叶片数量多、鲜红色,老叶脱落速率快;分枝数量多,梢长而细。(2)浙江楠生长状况一般,苗高、基径增长较快;新生叶片和老叶的更换速率缓慢;分枝数量也较少,秋梢较春梢和夏梢明显增长。(3)闽楠生长状况最缓慢,苗高、基径生长慢;叶片小而多;分枝少且梢短。
2.1.1 基径和苗高
从表1可以看出,浙江楠的平均基径最粗,桢楠其次,闽楠最细;桢楠的基径与浙江楠差异不明显,而浙江楠的基径与闽楠差异显著,且前者比后者高16.38%。相对于苗高而言,桢楠>浙江楠>闽楠;闽楠的苗高与浙江楠差异不显著,而桢楠苗高与闽楠、浙江楠苗高差异显著,且比闽楠高42.82%,比浙江楠高29.37%。表明3种楠木中,桢楠幼苗高大挺拔,浙江楠幼苗基径壮实,桢楠与浙江楠的幼苗长势较好,而闽楠的幼苗则较为弱小。
2.1.2 苗高、春梢、夏梢、秋梢比较
从表1可以看出,桢楠的苗高增量最大,闽楠最小;桢楠的苗高增量与闽楠差异显著,桢楠比闽楠苗高增量高59.27%,而浙江楠的苗高增量与闽楠、桢楠差异不显著。3种楠木不同季节的树梢生长量也不尽相同,春梢生长量处于3.11~6.43 cm:浙江楠>桢楠>闽楠,浙江楠、桢楠的春梢生长量与闽楠差异显著,分别高于闽楠106.75%、97.75%,浙江楠与桢楠差异不显著。夏梢生长量在4.98~5.66 cm:桢楠>闽楠>浙江楠,三者之间差异不显著。秋梢生长量为9.31~16.19 cm:桢楠>浙江楠>闽楠,桢楠的秋梢生长量与闽楠差异显著,桢楠高于闽楠 73.90%,而浙江楠与桢楠、闽楠则差异不显著。分析结果,秋季是楠木幼苗苗高生长的高峰期,显著高于春、夏2季,桢楠幼苗纵向生长速率最快。
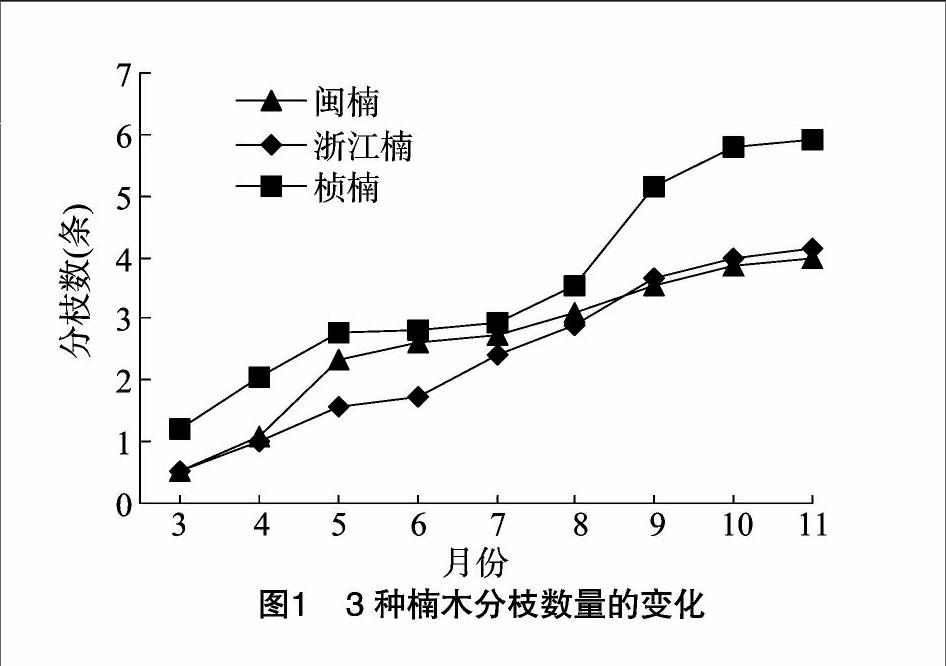
2.1.3 分枝数量的变化
从图1可以看出,3种楠木2年生幼苗的一级分枝数量在4~6条:桢楠>浙江楠>闽楠,且三者的分枝数差异不显著;3种楠木分枝数的生长峰值期都为3—5月,其他时间平缓增长,与时间呈一定的正相关,而桢楠在8—9月又进行了一次较为迅速的生长期。
2.1.4 新老叶数量的变化
从图2可以看出,测量初始期闽楠和浙江楠的老叶数多于桢楠,新叶数则低于桢楠;随着老叶的脱落和新叶的生长,测量结束时桢楠的新、老叶数都据首位;老叶的脱落速度为闽楠>浙江楠>桢楠,主要集中在3—5月脱落;新叶的增长速度为桢楠>闽楠>浙江楠,叶片在7—9月间达到生长峰值;桢楠老叶脱落速度明显较低,但其新叶生长速度最快。
2.2 幼苗光合特性
2.2.1 净光合速率
从图3-A可以看出,比较3种楠木的净光合速率,净光合率为桢楠>浙江楠>闽楠,桢楠、浙江楠的净光合速率与闽楠差异显著,桢楠与浙江楠差异不显著;追 肥处理结果相同,但相应提高了净光合速率,桢楠、浙江楠、闽楠净光合速率追肥处理分别高于未追肥处理6.20%、20.00%、6.58%,但差异不显著。
2.2.2 气孔导度
从图3-B可以看出,追肥处理的3种楠木气孔导度都比未追肥处理的高,未追肥处理的楠木中,闽楠的气孔导度最高,桢楠最低,桢楠的气孔导度与闽楠、浙江楠差异显著,气孔导度桢楠比闽楠、浙江楠分别低35.44%、3289%,闽楠与浙江楠差异不显著,前者比后者高3.68%;追肥处理的楠木中,浙江楠的气孔导度最高,桢楠最低,桢楠与浙江楠差异显著,桢楠低于浙江楠35.58%,而闽楠与桢楠、浙江楠差异不显著。
2.2.3 胞间CO2浓度
从图3-C可以看出,闽楠胞间CO2浓度最高,桢楠最低,追肥处理得到相同的结果,且追肥处理可以适当增加楠木幼苗的胞间CO2浓度,但闽楠追肥处理效果不明显;未追肥处理的闽楠、浙江楠、桢楠幼苗胞间CO2浓度间差异显著,且闽楠比浙江楠高28.91%,比桢楠高7828%,浙江楠比桢楠高38.30%;追肥处理闽楠、浙江楠、桢楠胞间CO2浓度间差异显著,且闽楠比浙江楠高14.43%,比桢楠高 44.36%,浙江楠比桢楠高26.15%。桢楠追肥处理和未追肥处理幼苗间差异显著,追肥处理比未追肥处理高17.86%。
2.2.4 蒸腾速率
从图3-D可以看出,未追肥处理3种楠木的蒸腾速率都低于追肥处理的,未追肥处理的闽楠叶片蒸腾速率最高,桢楠最低;追肥处理的浙江楠蒸腾速率最高,桢楠最低。未追肥处理的幼苗中,闽楠、浙江楠蒸腾速率间差异不显著;追肥处理幼苗中,闽楠与浙江楠蒸腾速率差异不显著,但浙江楠与桢楠间差异显著,浙江楠比桢楠高 32.49%。浙江楠中追肥处理和未追肥处理间差异显著,追肥处理比未追肥处理高32.39%。
2.3 幼苗生理特性
2.3.1 叶绿素相对含量
从表2可以看出,桢楠的叶绿素相对含量最高,闽楠其次,浙江楠最低,追肥处理可以提高楠木幼苗叶绿素相对含量,但闽楠却呈下降趋势;未追肥处理的3种幼苗中,桢楠、闽楠与浙江楠差异显著,分别高于浙江楠1463%、14.33%,桢楠与闽楠间差异不显著;追肥处理的闽楠与浙江楠、桢楠间差异显著,追肥处理的浙江楠与桢楠间差异不显著;追肥处理与未追肥处理的同种幼苗间差异显著。
2.3.2 营养物质含量
从表2可以看出,3种楠木幼苗的可溶性糖含量为浙江楠>闽楠>桢楠,且浙江楠的可溶性糖含量与闽楠、桢楠差异显著,闽楠與桢楠间则差异不显著;追肥处理后,可适当提高幼苗内可溶性糖的含量,与未追肥处理幼苗差异不显著,追肥处理导致桢楠幼苗可溶性糖含量下降。
3种幼苗的可溶性蛋白含量由高到低依次为浙江楠>桢楠>闽楠。追肥处理可以提高楠木幼苗可溶性蛋白含量,但对闽楠效果不佳,且与未追肥处理的幼苗之间差异不明显;追肥处理后,浙江楠的可溶性蛋白含量急剧增高,与闽楠差异显著。
2.3.3 超氧化物歧化酶活性
从表2可以看出,桢楠幼苗的SOD活性最强,闽楠最低,桢楠与浙江楠差异显著,闽楠与浙江楠间差异不显著;追肥处理会降低楠木幼苗的SOD活性,追肥处理与未追肥处理浙江楠的SOD活性差异显著,追肥有效降低浙江楠幼苗的SOD活性,但增强了闽楠幼苗的SOD活性。
2.3.4 丙二醛含量
由表2可知,浙江楠幼苗的MDA含量最高,桢楠其次,闽楠最低;追肥处理提高浙江楠、桢楠幼苗叶片中MDA含量,分别高出未处理幼苗8.56%、7.94%,但差异不显著,追肥处理却使闽楠幼苗叶片中MDA含量迅速下降,下降了41.55%,追肥处理闽楠的MDA含量与浙江楠、桢楠差异显著。
2.3.5 根系活力
由表2可知,追肥处理桢楠与浙江楠的根系活力下降,分别下降20.89%、11.36%,但闽楠的根系活力上升12.02%,追肥仅对桢楠幼苗的根系活力有显著影响。
3 结论与讨论
植物株高、基径以及叶枝数量的变化是植物生长最直观的表现,研究楠木幼苗生长的基本指标有助于了解楠木幼苗生长特性的规律[15]。本研究结果,3种楠木幼苗的基径随时间均匀增长,与时间呈一定的线性关系;苗高增长高峰期出现在7—9月,由于此时雨水增多、温度适宜,促进楠木幼苗生长发育,这与黄志玲等的研究结果[16-17]一致;分枝数量在3—5月迅速增长,新叶增长速率的高峰期在7—9月出现,而老叶在3—5月集中脱落,这是由于楠木幼苗经过寒冬后,对开春温度上升的反应,逐渐进入生长高峰期;10月之后,荆州天气开始降温,3种楠木幼苗的生长都趋于平缓,至11月接近停止生长,而闽楠的新叶出现部分脱落现象,这与闽楠喜阴而不耐寒的特性相吻合[18]。比较3种楠木幼苗的生长性状可知,桢楠的苗高、苗高增量、分枝数量等生长指标都高于浙江楠与闽楠,为移栽后表现最好的幼苗,闽楠则表现最差。
植物通过光合作用进行营养物质的积累,光合特性是检验植株生长状况的有效指标[19]。分析表明,叶绿素含量与光合速率呈显著或极显著正相关[20-22]。本研究结果表明,3种楠木幼苗中叶绿素相对含量与净光合速率呈正相关,气孔导度、胞间CO2浓度、蒸腾速率与净光合速率呈负相关;影响桢楠净光合速率的主要因子是叶绿素含量。追肥处理对3种楠木幼苗的光合生理具有明显作用,可提高净光合速率 6.20%~20.00%,桢楠反应最为显著,相关研究表明,随施肥浓度的增加净光合速率呈先上升后下降的趋势[23],3种幼苗对相同浓度的肥料感应敏感程度不同,可适当增加闽楠和浙江楠的肥料浓度;追肥处理后,浙江楠的气孔导度与蒸腾速率急剧上升,由于其胞间CO2浓度未随之上升,因此不影响其净光合速率。比较3种楠木幼苗的光合指标,桢楠的光合作用能力最强,与其生长指标优于浙江楠和闽楠相一致。
生理指标可以反映植物生长状况,同时也反映了植物的生长能力[24]。本试验研究表明,在营养物质含量上,桢楠和浙江楠在一定程度上都高于闽楠,与三者的生长差异相符合,由于可溶性糖和蛋白类似于信号分子的存在,参与调控苗木的生长、发育、成熟和衰老等过程[25];SOD活性和MDA含量桢楠和浙江楠也相对优于闽楠,表明桢楠和浙江楠在适应环境、抵抗外界干扰的能力上要强于闽楠;桢楠的根活力最强,说明桢楠根吸收水和养分的能力更强,与其生长指标相对应。追肥处理后,有助于提升楠木可溶性糖和可溶性蛋白及MDA含量,而降低SOD含量与根系活力,但闽楠表现异常,这与其生长状况相联系。
在3种楠木幼苗中桢楠生长状况最好,对环境的适应能力较强,生长也相对较快,适于作为园林应用树种进行推广,同时也可作为保护楠木的典型树种进行大量繁殖栽培。本试验研究范围有限,楠属的其他几种楠木,如紫楠、滇楠、白楠等未进行研究,还有待今后进一步研究完善。
参考文献:
[1] 葛继稳,吴金清,朱兆泉,等. 湖北省珍稀濒危植物现状及其就地保护[J]. 生物多样性,1998,6(3):220-228.
[2]李冬林,丁彦芬,向其柏. 浙江楠引种育苗技术[J]. 林业科技开发,2003,17(3):43-45.
[3]李冬林,金雅琴,向其柏. 我国楠木属植物资源的地理分布、研究现状和开发利用前景[J]. 福建林业科技,2004,31(1):5-9.
[4]葛永金,王军峰,方 伟,等. 闽楠地理分布格局及其气候特征研究[J]. 江西农业大学学报,2012,34(4):749-753,761.
[5]范剑明,谢金兰. 楠木栽培技术[J]. 广东林业科技,2007,23(6):94-96.
[6]林朝楷,黄世贵. 浅谈楠木播种育苗技术[J]. 中国园艺文摘,2010,26(1):91.
[7]王伙琴. 楠木容器播种育苗技术研究[J]. 现代农业科技,2013(10):150-151.
[8]朱 雁. 珍贵树种楠木容器苗芽苗移栽技术[J]. 中国林副特产,2010(3):54-55.
[9]王 艺,王秀花,张丽珍,等. 不同栽培基质对浙江楠和闽楠容器苗生长和根系发育的影响[J]. 植物资源与环境学报,2013,22(3):81-87.
[10] 钟灼坤. 楠木高效快速育苗试验分析[J]. 现代园艺,2016(3):10-11.
[11]刘 刚,涂 铭,朱 丽,等. 不同浓度盐胁迫下闽楠和楠木光合生理研究[J]. 湖北林业科技,2014,43(6):9-12,20.
[12]黄晓蓉,李玮婷,刘 刚,等. 水分胁迫对楠木幼苗抗逆生理特性的影响[J]. 北方园艺,2015(7):68-71.
[13]李晓明. 荆州市休闲观光农业发展研究[D]. 荆州:长江大学,2013.
[14]麻文俊,张守攻,王军辉,等. 1年生楸树无性系苗期生长特性[J]. 林业科学研究,2012,25(5):657-663.
[15]肖 遥,楚秀丽,尹增芳,等. 不同产地南方红豆杉各家系幼苗生长、光合生理与株高生长节律的差异分析[J]. 植物资源与环境学报,2016,25(1):34-42.
[16]黄志玲,郝海坤,庞世龙,等. 红锥种源苗期生长节律研究[J]. 林业科技开发,2012,26(3):24-28.[HJ1.72mm]
[17]周 诚,廖海红,王丽艳,等. 毛红椿播种苗苗期生长规律分析[J]. 林业科技开发,2013,27(1):18-21.
[18]李玮婷,宗 卫,费永俊. 水杨酸和氯化钙对低温条件下闽楠生理指标的影响[J]. 林业科技开发,2015,29(5):23-26.
[19]Tuittila E S,Vasander H,Laine J. Sensitivity of C sequestration in reintroduced sphagnum,to water-level variation in a cutaway peatland[J]. Restoration Ecology,2004,12(4):483-493.
[20]娄义龙,刘振兴. 烟草光合作用的遗传研究 Ⅳ. 叶绿素含量的遗传及其与产量的关系[J]. 山地农业生物学报,1994,13(2):1-7.
[21]徐 坤,鄒 琦. 生姜光合特性研究再探[J]. 山东农业大学学报(自然科学版),2000,31(2):147-150.
[22]魏书銮,于继洲,宣有林,等. 核桃叶片的叶绿素含量与光合速率的研究[J]. 北京农业科学,1994,12(5):31-33.
[23]贺 维,胡庭兴,王 锐,等. 施肥对桢楠幼苗光合生理及生长特性的影响[J]. 西北植物学报,2014,34(6):1187-1197.
[24]常冀原. 施肥对江南油杉幼苗生理特性的影响[D]. 长沙:中南林业科技大学,2015.
[25]Geigenberger P,Stitt M. Sucrose synthase catalyses a readily reversible reaction in vivo in developing potato tubers and other plant tissues[J]. Planta,1993,189(3):329-339.
猜你喜欢
热带农业科学(2016年11期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年12期)2017-01-21
山东农业科学(2016年11期)2016-12-17
现代园艺(2016年9期)2016-07-16
现代园艺(2016年2期)2016-03-15
