
春季杉木球果与叶片的光合特征比较
2018-01-05 02:38左丹丹曹世江林思祖
四川农业大学学报 2017年4期
左丹丹 ,曹世江 ,李 佳 ,熊 升 ,林思祖 ,陈 宇 *
(1.福建农林大学林学院;2.林木逆境生理生态及分子生物学福建省高校重点实验室;3.国家林业局杉木工程技术研究中心,福州 350002))
春季杉木球果与叶片的光合特征比较
左丹丹1,2,曹世江2,3,李 佳2,3,熊 升2,3,林思祖1,3,陈 宇2,3*
(1.福建农林大学林学院;2.林木逆境生理生态及分子生物学福建省高校重点实验室;3.国家林业局杉木工程技术研究中心,福州 350002))
【目的】明确杉木球果在春季生长旺盛期的光合生理特性。【方法】以杉木球果为材料,以同一枝条的叶片为参照,对其光合色素含量、叶绿素荧光参数和光合作用参数进行比较研究。【结果】球果中的叶绿素a(Ca)、叶绿素b(Cb)、叶绿素总量(CT)和类胡萝卜素(Cx.c)均低于叶片,但在春季的生长过程中,均伴随时间推移而呈增加趋势;初始荧光(F0)、最大荧光(Fm)和可变荧光(Fv)均高于叶片且呈上升趋势,非光化学淬灭(NPQ)和光化学淬灭(qp)均低于叶片且呈下降趋势;净光合速率(Pn)最初为负值,而后逐渐增强,于4月下旬表现为正值,表明球果光合作用强度由弱变强,于4月下旬开始强于呼吸作用;气孔导度(Gs)随时间小幅上升,胞间CO2浓度(Ci)明显大于叶片。【结论】春季杉木球果具备一定的光合作用能力,且在春季生长过程中呈逐渐增强的趋势,但明显弱于叶片。
杉木;光合作用;球果;叶片;春季
光合作用是植物最为重要的生理活动之一。植物体通过吸收、传递、分配和利用环境光能,将之转化成可供体内所有代谢活动消耗的化学能,并最终影响植物的生长、发育、抗性和生殖等一系列生理过程[1]。植物光合作用能力特征评价体系主要有光合速率指标体系和叶绿素荧光参数体系,目前已广泛应用于植物的抗性生理测定[2-4]、生长发育评价[5]、种质资源评估[6-7]以及引种驯化等领域[8]。在裸子植物中,植株个体中可进行光合作用的组织器官主要为叶片,除此之外,多数裸子植物的球果中也含有一定浓度的叶绿素,可进行光合作用。已有文献报道,球果的光合作用与球果生物量之间存在显著的相关性,并可进一步影响种子质量[9]。可见,球果的光合生理特征对于种子发育有重要的作用。
作为我国最为重要的用材林树种之一,杉木(Cunninghamia lanceolata(Hook)Lamb.)肩负着全国约37.4%的木材产量,地位举足轻重。目前,杉木在光合作用方面的研究主要以叶片为研究对象,探讨其抗性表现[10];或是对不同叶龄、叶位,乃至不同种源间的光合能力进行评价[8,11-12]。而有关杉木球果的光合作用能力强弱和光合生理特性评价的研究尚未见报道。因此,本研究以春季杉木生长旺盛期的球果为研究对象,通过与叶片的比较,分析其光合作用相关参数的表现,为深入开展球果光合作用模式分析及其对种子发育的影响提供参考。
1 材料和方法
1.1 供试材料
本试验所用材料为国家林业局杉木工程技术研究中心筛选的优良无性系,2010年春出圃并定植于福建农林大学。于2016年3月12至5月15日每7 d取样一次,共取10次。在取样日早上10:00—10:30选取生长势一致且无明显病虫害的3株单株,从每株单株上选择生长健康、大小一致、着生位置和朝向相同的球果,连枝条剪下,插于带水的广口瓶中迅速带回实验室进行球果的叶绿素含量、叶绿素荧光参数和光合速率的测定,分别测定不同单株材料以作为生物学重复,并以同枝的成熟叶片作为对照。
1.2 研究方法
1.2.1 光合参数测定
光合速率测定采用LICOR公司生产的Li-6400XT portable Photosynthesis System光合测定仪,测定采用针叶叶室,CO2浓度为(380±10)μL/L,测定光强为2000 Lux。测定球果的光合速率时将球果下方的6片叶片剪下去除,然后将球果置入光合测定仪叶室内,使测定时无叶片干扰。测定叶片的光合速率时采用与球果同一枝条上的叶片,自球果底部而下第8片叶起,向下共10片叶片作为测定叶。供试材料在叶室中经过20 min光照适应(2000Lux),待仪器数值稳定后开始读取测定值,测定时间为3 min,每5 s读取1次,测定指标为净光合速率(Pn)、胞间二氧化碳浓度(Ci)和气孔导度(Gs)。球果表面积Wang W.等[12]的方法,叶片表面积采用Imagepro plus v6.0软件计算。
1.2.2 叶绿素荧光参数测定
以光合参数相同的球果和测定叶进行叶绿素荧光参数测定。供试材料经20 min暗适应后,采用Photom Systems Instruments公司生产的Handy Flour Cam荧光成像系统,以Quenching模式测定初始荧光值F0、最大荧光值Fm、可变荧光值Fv、最大光量子产量Fv/Fm、非光化学淬灭系数NPQ和光化学淬灭系数qp。
1.2.3 光合色素含量测定
分别称量球果上的鳞片以及测定叶各0.5 g用于叶绿素含量测定。测定方法参考赖善聪等[11]的方法,即乙醇丙酮混合液法,测定叶绿素a(Ca),叶绿素b(Cb),叶绿素总量(CT)和类胡萝卜素(Cx.c)含量。
1.2.4 数据处理
各测定数值输入Excel 2013后计算平均值与标准差,以Origin v.9.1软件作图。
2 结果与分析
2.1 杉木球果与叶片光合色素含量的比较
由图 1(a)~图 1(d)可知,春季杉木球果与叶片中Ca、Cb、CT和Cx.c在春季均随时间而呈上升趋势。5月15日末次测定值与3月12日初次测定值相比,球果的 Ca、Cb、CT和 Cx.c分别增长了 169.7%、25.9%、96.4%和556.9%;而在叶片中则分别增长了26.1%、12.4%、22.4%和18.8%,可见球果中的光合色素增长趋势显著高于叶片的。球果与叶片中Ca/Cb值在春季也呈上升趋势(图1(e)),末次测定与首次测定相比,增长率分别为114.2%和12.3%,这说明球果中Ca的积累速度较Cb更快,且高于叶片。相较于成熟叶片中较为稳定的CT/Cx.c值(末次测定与首次测定相比,增长率为3.0%),球果中的CT/Cx.c急剧下降(图 1(f)),下降率达到 78.3%,4 月中下旬下降至略高于叶片的水平,之后下降趋势变缓。总体而言,春季杉木球果中的光合色素含量虽显著低于叶片,但仍有一定的浓度水平,且不断增长,具备发生光合作用的物质基础。
2.2 杉木球果与叶片叶绿素荧光参数比较
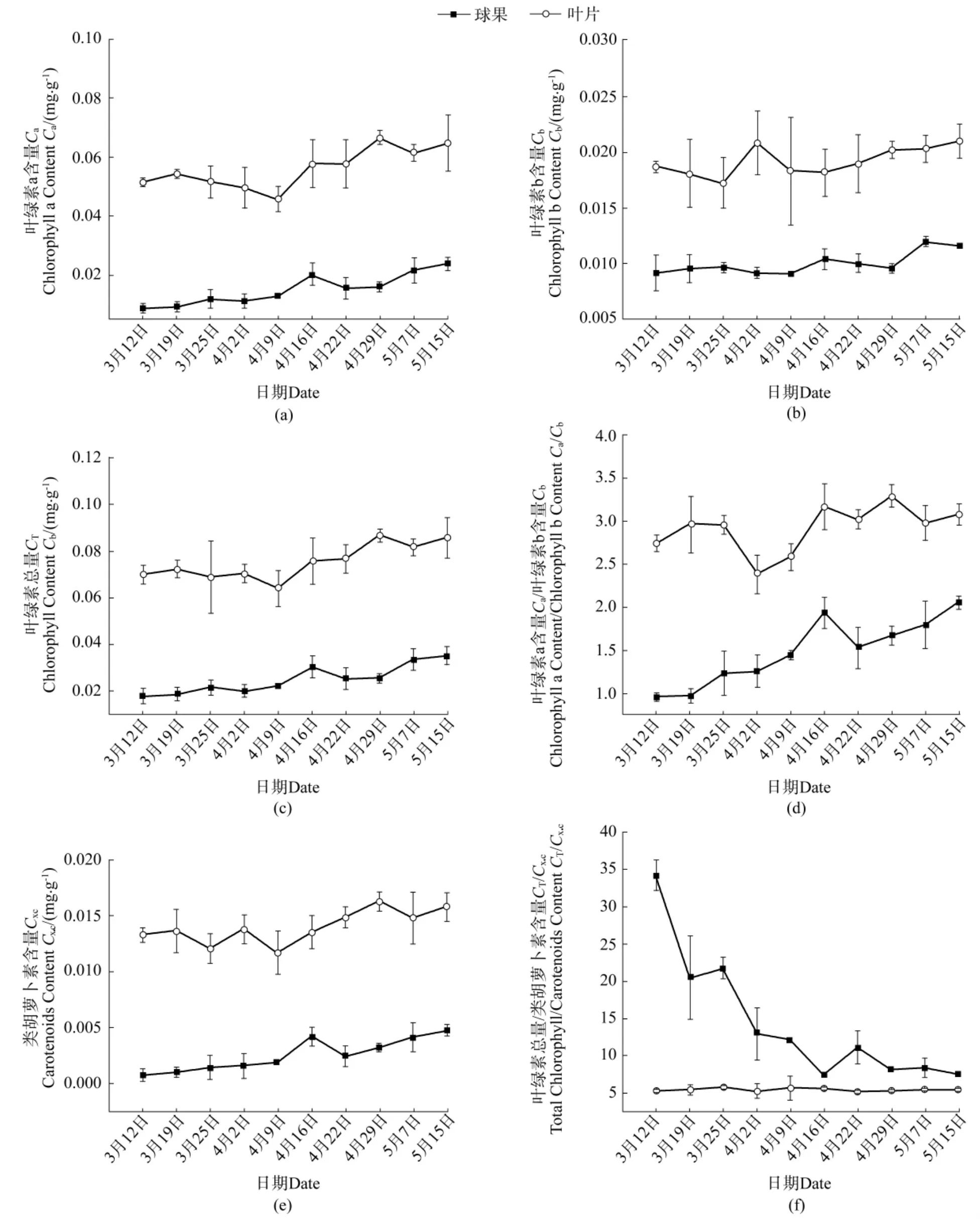
图1 杉木球果与叶片光合色素含量Figure1 Photosynthetic pigment content between cones and leaves of Chinese fir
由图2可知,春季杉木球果与叶片的F0、Fm和Fv在3月中旬表现相当,随后的表现则有所不同:3月下旬至4月上旬,球果的F0和Fm呈下降趋势,伴随时间的推移,又呈上升趋势,且上升幅度相对较大,但Fv从3月下旬至4月中旬均呈上升趋势,其后表现为上升趋势;叶片的F0、Fm和Fv在3月上中旬的变化幅度不大,但3月下旬至4月上旬有较大幅度的升高,自4月中旬起至4月底随时间变化而呈小幅下降趋势,随后又呈上升趋势(图2(a)~图2(c))。总体而言,球果的F0、Fm和Fv上升速度高于叶片。一般认为,F0来自天线叶绿素a,代表不参与光化学反应的光能辐射部分。F0在不同组织中的差异表现体现了叶绿素a的差异;而Fm也与叶绿素浓度呈负相关。这与图1(a)的结果一致,表明了球果中的PSⅡ体系转化潜力与原初反应强度均弱于叶片。

图2 杉木球果与叶片叶绿素荧光参数比较Figure2 Comparison of chlorophyll fluorescence parameters between cones and leaves of Chinese fir
球果与叶片的Fv/Fm的变动范围均在0.75~0.85之间(图2(d)),表明两种组织在春季未受环境条件的抑制。但叶片在春季的Fv/Fm值呈较为明显的下降趋势,而球果则较为稳定,表明球果组织中的光量子转换效率受环境因素影响可能更小。
在过剩光能耗散途径方面,球果的NPQ总体呈缓慢下降的趋势,而叶片则呈先上升后下降的趋势(图2(e));球果的qp表现为振荡式的快速下降,而叶片则呈小幅上升趋势(图2(f))。这表明相较于叶片,球果通过热耗散形式和光化学形式消耗的过剩光能较少。
2.3 杉木球果与叶片光合作用参数比较
由图3可知,球果的Pn与叶片相比有较大的差距。自3月中旬至4月中旬,杉木球果的Pn为负值,表明这一阶段其组织的呼吸作用强度大于光合作用强度。随着球果不断发育,其光合作用强度也不断增强,其增强幅度大于叶片的幅度。至4月下旬起,其Pn值开始转变为正值,5月中旬增长至2.08 μmol/(m2·s),相当于叶片的70.4%,表明其组织的光合作用强度已大于呼吸作用(图3(a))。这与球果的CT/Cx.c值下降至较为稳定值的时间点(图2(f))相吻合,反映出球果组织内光合作用结构的成熟。
球果的Gs值在春季也呈逐渐上升趋势,但与叶片相比,上升幅度并不大(图3(b)),这表明球果气孔张开程度在春季增长程度有限,可能与球果的组织特异性有关。球果的Ci值大于叶片(图3(c)),表明球果组织中有充足的CO2以供给光能转换,这可能是由于球果较强的呼吸作用与较低的气孔导度共同造成。而球果中较低的净光合速率可能并非由CO2限制造成,而与其组织中光合色素含量水平有关。

图3 杉木球果与叶片光合作用参数比较Figure3 Comparison of photosynthesis parameters between cones and leaves of Chinese fir
3 讨论与结论
花是种子植物为了适应生殖而进化出的具有生殖功能的变态叶。尽管一些学者认为裸子植物的生殖器官并不是真正的花,而称之为孢子叶球,但从形态发生学角度讲,其与被子植物花器官基本上是同源的[14]。球果类裸子植物经由单性复轴演化线演化出苞鳞-种鳞复合体构造的大孢子球型生殖器官[15],与大多数被子植物功能高度分化的花器官不同,球果除能够保护种子在其发育过程中不受外界环境条件的影响之外,还具备碳同化作用的功能。大多数球果在发育过程中其外观表现为绿色,其组织内含有可利用光能的光合色素;球果上的种鳞从形态上看也更接近叶器官。但对于球果光合作用能力强弱,以及其同化作用对种子质量的影响,目前知之甚少。
从本研究的结果来看,杉木球果的光合色素含量虽在春季随时间而显著增加,且增加幅度大于叶片,但其总量还远小于叶片。赖善聪等[11]研究春季新叶、成熟叶与老叶在叶绿素总量与叶绿素荧光表现的差异,认为4月份杉木新抽嫩叶的叶绿素总量约为成熟叶的一半;而在本研究中球果的叶绿素总量在5月中旬末次测量时也仅为成熟叶的1/3左右。这可能是因为,一方面,与球果同生于一个枝条上的叶片在发育时间上早于球果,光合作用相应组织较球果发育更完全;另一方面,叶片是植物同化作用的主要器官,长期进化作用的结果使其功能集中于将外界光能转化为化学能以供植株生长发育,故而在叶片中可检测到更高的光合色素含量。球果的Cx.c在春季发育过程中迅速积累,叶绿素与类胡萝卜素比值(CT/Cx.c)迅速下降,这可能是球果组织趋于成熟的一种表现。
叶绿素荧光参数可反映植物光合作用的原初反应过程,是植物对环境光能吸收、光化学反应及光量子激发传递的体现。最大光化学量子效率Fv/Fm是衡量植物体是否受外界环境胁迫的一个敏感高效的指标,绿色植物在未受外界环境胁迫的条件下Fv/Fm一般在0.75~0.85之间,且不受物种与组织差异的影响[16]。春季杉木球果与叶片的Fv/Fm都在正常范围内,这说明球果的低光合作用水平并非由外界环境胁迫因素引起。其次,初始荧光F0表示植物PSⅡ系统反应中心完全开放时的荧光产量,一般在植物受到胁迫时会显著上升,较为幼嫩的组织中因叶绿素含量相对较少,F0的表现也相对较高[8,17-18]。本研究中球果的F0显著高于叶片,表明春季球果的光合作用主要限制因素为叶绿素含量。球果与叶片对过剩光能的消耗途径不同,则体现了二者的组织差异性。
杉木球果的净光合速率在3月中旬至4月中旬先为负值,后逐渐增强,于4月下旬表现为正值,表明球果光合作用强度由弱变强,于4月下旬开始强于呼吸作用;气孔导度小幅上升,细胞间隙CO2浓度明显大于叶片,表明球果组织对CO2的羧化效率较低,影响光合作用强度主要因素为非气孔限制。A.Andres等[9]在对挪威云杉球果的呼吸作用与光合效率的研究中发现,挪威云杉球果发育初期其光合作用呈上升趋势,对呼吸作用产生的CO2再利用效率达到16%~17%。另外,在落叶松的研究中也发现了其球果呼吸作用产生的CO2使得胞间CO2浓度大于叶片,且有高达30%~40%的可被再利用于光合作用[19]。本研究中杉木球果春季光合作用趋势与挪威云杉相同,而从气孔导度和胞间CO2浓度表现看,可能也存在对呼吸作用产生的CO2的再利用过程。但落叶松球果发育初期净光合速率达到5.3 μmol/(m2·s),是叶片净光合速率(约为 4.0 μmol/(m2·s),与杉木叶片的 Pn相当)的 1.26 倍[19]。有关杉木球果与落叶松球果光合作用强度的差异,还有待进一步研究。
本研究仅就春季杉木球果的光合作用特征展开简单的探讨,结果表明,杉木球果在春季虽已具备一定的光合作用能力,但还明显低于同一时期的成熟叶片。有关杉木球果的光合作用变化规律,还有很多问题有待进一步研究,例如同化作用产物是否对种子发育提供养分支持,其贡献度有多大,是否对杉木种子败育产生影响,还需对球果长达13个月的发育过程进行跟踪观测。
[1]SCARTAZZA A,DI B D,BERTOLOTTO P,et al.Investigating the European beech(Fagus sylvatica L).leaf characteristics along the vertical canopy profile:leaf structure,photosynthetic capacity,light energy dissipation and photoprotection mechanisms[J].Tree Physiology,2016,36(9):1060.
[2]隆春艳,古洪辉,汪正香,等.外源脱落酸对高温胁迫下菠菜光合与叶绿素荧光参数的影响[J].四川农业大学学报,2017,35(1):24-30.
[3]王舒甜,张金池,张亮.油菜素内酯对盐胁迫下香樟叶片光合色素以及叶绿素荧光的影响[J].浙江大学学报(农业与生命科学版),2017,43(1):45-53.
[4]SHI D W,WEI X D,CHEN G X.Effects of low temperature on photosynthetic characteristics in the super-high-yield hybrid rice'Liangyoupeijiu'at the seedling stage[J].Genetics&Molecular Research,2016,15(4).
[5]LIU T,WANG Z,CAI T.Canopy Apparent Photosynthetic Characteristics and Yield of Two Spike-Type Wheat Cultivars in Response to Row Spacing under High Plant Density[J].Plos One,2016,11(2):e0148582.
[6]姜武,沈志军,姜卫兵,等.不同季节水蜜桃品种光合生理指标的比较[J].江苏农业学报,2008,24(3):321-330
[7]唐敏,翟秀明,姚永红,等.不同茶树品种(系)叶绿素荧光参数季节性差异研究[J].西南农业学报,2015,28(1):79-83.
[8]Nguyen Ba Trieu,陈宇,林小琴,等.不同种源越南杉木幼龄期叶绿素荧光特征比较[J].四川农业大学学报,2016,34(1):34-38,47.
[9]KOPPEL A,TROENG E,LINDER S.Respiration and photosynthesis in cones of Norway spruce(Picea abies(L.)Karst.)[J].Trees,1987,1(2):123-128.
[10]吴鹏飞,马祥庆,邹显花,等.钙对低磷胁迫杉木叶绿素及其荧光特性的影响[J].中国农学通报,2011,27(13):20-24.
[11]赖善聪,佘志媛,石零珊,等.杉木幼苗不同叶位叶绿素含量及荧光特性差异[J].莆田学院学报,2015,22(5):28-31.
[12]黄华宏,陈奋学,童再康,等.矮生杉木光合特性及叶绿素荧光参数研究[J].北京林业大学学报,2009,31(2):69-73.
[13]WANG W,ZU Y,CUI S,et al.Carbon dioxide exchange of larch(Larix gmelinii)cones during developmen[tJ].Tree Physiology,2006,26(10):1363-1368.
[14]谷安根,汪矛,王立军.蕨类植物和裸子植物的起源与进化[J].植物学通报,1990,7(3):58-62.
[15]葛传吉.从蕨类植物到被子植物看生殖器官的进化[J].生物学通报,1983(03):9-11.
[16]姜英,黄志玲,郝海坤.不同光照强度下红锥幼苗叶绿素荧光参数的季节变化[J].西南林业大学学报.2014,34(2):8-12.
[17]BAUNTHIYAL M,SHARMA V.Response of three semi-arid plant species to fluoride;consequences for chlorophyll florescence[J].International Journal of Phytoremediation,2014,16(4):397.
[18]TRIPATHI D K,SINGH V P,PRASAD S M,et al.Siliconmediated alleviation of Cr(VI)toxicity in wheat seedlings as evidenced by chlorophyll florescence,laser induced breakdown spectroscopy and anatomical changes[J].Ecotoxicology&Environmental Safety,2015,113:133-144.
[19]陈曦,陈彬丽,周晨,等.生长季油松针叶叶绿素荧光对温度变化的响应[J].北京林业大学学报,2011,33(6):70-74.
Comparison of Photosynthetic Characteristics between Chinese Fir Cones and Leaves in Spring
ZUO Dan-dan1,2,CAO Shi-jiang2,3,LI Jia2,3,XIONG Sheng2,3,LIN Si-zu1,3,CHEN Yu2,3*
((1.College of Forestry,Fujian Agriculture and Forestry University;2.Forest Adversity Physiological Ecology and Molecular Biology of Fujian;3.State Forestry Administration Engineering Research Center of Chinese Fir;Fuzhou 350002,China)
【Objective】The aim of the study was to reveal the photosynthetic capacity of cones of Chinese fir(Cunninghamia lanceolate)in spring.【Method】The photosynthetic pigment contents,photosynthetic properties and chlorophyll florescence were measured in the cones and mature leaves of the same branches.【Results】During development process of cone in spring,the contents of Ca,Cb,CTand Cx.cin cones gradually increased,but often were lower than those in leaves.The values of F0,Fm and Fvin cones were higher than those in leaves.The values of NPQ and qpwere lower as compared to the values of leaves.The Pn was minus before late April,reflecting there was a weak photosynthetic activity in contrast to respiration in cones.However,it grew fast and then exceeded the respiration rate.The Gsvalue increased slightly with time and the Ciwas significantly greater than that in leaves.【Conclusion】In general,the cone of Chinese fir has the photosynthetic capacity.Its Pngradually increase during the spring growth but is generally weaker than that of the leaves.
Chinese fir;photosynthesis;cones;leaf;spring
Q948
A
1000-2650(2017)04-0523-06
10.16036/j.issn.1000-2650.2017.04.010
2017-10-28
国家自然基金项目(KAe16010A);国家林业局杉木工程技术研究中心孵化基金(6213C011103)。
左丹丹,硕士研究生。*责任作者:陈宇,研究实习员,主要从事林木遗传育种研究,E-mail:28811852@qq.com。
(本文审稿:陈良华;责任编辑:巩艳红;英文编辑:徐振锋)
猜你喜欢
科学(2022年4期)2022-10-25
农民致富之友(2020年8期)2020-05-11
广西林业科学(2019年2期)2019-07-25
中国林副特产(2019年3期)2019-01-10
福建林业(2018年3期)2018-11-07
现代园艺(2017年23期)2018-01-18
现代园艺(2017年23期)2018-01-18
Coco薇(2016年5期)2016-06-03
广西林业科学(2016年2期)2016-03-20
考试周刊(2016年6期)2016-03-11
