
岭南槭不同种源家系种子形态和幼苗生长变异
2018-01-03 00:52:05吴培培罗水兴闫玮明
林业科学研究 2017年6期
吴培培,孙 冰,罗水兴,陈 雷,闫玮明
(中国林业科学研究院热带林业研究所,广东 广州 510520)
岭南槭不同种源家系种子形态和幼苗生长变异
吴培培,孙 冰*,罗水兴,陈 雷,闫玮明
(中国林业科学研究院热带林业研究所,广东 广州 510520)
目的探讨不同种源家系岭南槭种子形态、种子萌发与幼苗生长的特性,分析它们与种源地地理气候因子之间的关系。方法对广东省岭南槭天然分布区的3个种源18个家系的种子表型性状、发芽特征、幼苗生长性状和叶色表现进行方差分析和相关性分析。结果表明:肇庆黑石顶(HSD)种源的种子最厚、最宽,深圳七娘山(QNS)种源的种子最薄、最窄,惠州南昆山(NKS)种源的种子最长,种子千粒质量最高;翅长与翅宽呈极显著正相关(P<0.01),种子千粒质量与发芽率显著正相关(P<0.05);岭南槭1年生幼苗地径与苗高成正比,幼苗生长受种源地的地理气候因子影响;在色彩表现方面,QNS3、QNS13家系的明度L*值较大,QNS13色相b*值显著高于其他家系,NKS14家系幼苗的色相a*值最大。结论种子千粒质量越高,发芽率越高;幼苗的地径苗高生长受种源地的地理气候因子影响。NKS14家系幼苗红叶特征最明显,色彩表现较好。
岭南槭;种子形态;种子萌发;幼苗生长;叶色

目前,槭属植物作为优良的彩叶观赏植物引起了学者的广泛关注[3-7]。陈勇等[5]调查发现,生态风景林带中广泛应用的彩叶植物较少,岭南槭作为彩叶观赏植物尚未被开发。近些年,关于岭南槭的研究主要集中在嫁接[8]、耐热性[9]以及种子发芽率[1, 10-12]等方面,并未对岭南槭种子表型、幼苗生长和叶色特征的变异进行进一步研究。本文通过对不同种源家系岭南槭种子形态、发芽特征、幼苗生长情况及色彩表现进行分析,为进一步开展岭南槭选育工作奠定基础。
1 试验材料
试验材料为岭南槭3个种源18个家系的种子,均采自岭南槭天然分布区。根据前期实地考察,在广东省寻找岭南槭典型集中分布区域,并采集种子,单株作为1个家系。经过3年野外调查发现,岭南槭野外分布较少,种质资源匮乏,结实率低,已尽可能收集较多不同种源家系的种子。其中,肇庆市黑石顶(HSD)自然保护区4个家系,深圳市七娘山(QNS)国家地质公园11个家系,惠州市南昆山(NKS)自然保护区3个家系。参试岭南槭种源信息及其气候资料见表1,采种母株的信息见表2。

表1 参试岭南槭种源地理信息及其气候资料

表2 岭南槭采种母树的生物学性状及环境因子
2 研究方法
2.1 种子形态的测量
每个家系随机抽取3份完整的种子,每份100粒,用游标卡尺(0.01 mm)分别测量种厚、种长、种宽、翅长和翅宽。每个岭南槭家系随机抽取8份自然干燥状态下纯净、饱满的种子,每份100粒,用电子天平(0.000 1 g)分别称其质量,计算千粒质量。
2.2 种子发芽率的测量
发芽率(GR)=种子发芽总数/供试种子数×100%
2.3 幼苗生长性状与叶色表现测量
分别于2016年9月(幼苗0.5年生阶段)、2017年3月(幼苗1年生阶段)对岭南槭幼苗进行调查,每个小区随机调查50株,测量苗高、地径和叶色值。用卷尺测量苗高、电子游标卡尺(0.01 mm)测量地径。用CM-2600d分光测色仪在10°/ D65光源下测量叶色(包括明度L*值、色相a*值和b*值),明度L*表示白色至黑色范围,L*值越大说明叶色越亮;色相a*值表示红色至绿色范围,a*值越大,说明叶色越红;色相b*值表示黄色至蓝色范围,b*值越大,说明叶色越黄。
试验数据采用Excel 2007软件绘制图表,运用SPSS 20.0软件进行方差分析、相关性分析和Duncan多重比较。
3 结果与分析
3.1 种子形态的变异分析
岭南槭不同种源家系种子形态变异分析结果(表3)表明:不同种源间的种厚、种长、种宽、翅长和翅宽均差异显著(P<0.05),但种子千粒质量差异不显著;HSD种源的种厚、种宽、翅长和翅宽最大,QNS种源的种厚、种长、种宽、翅长和翅宽均最小,NKS种源的种长最大且千粒质量最高。HSD、QNS和NKS各种源内不同家系间的种子形态绝大多数差异显著(P<0.05),仅NKS不同家系的种厚差异不显著。HSD9家系的翅长(25.92 mm)最长,HSD20家系的翅宽(9.36 mm)最大;QNS2家系的种宽(4.54 mm)和种子千粒质量(66.410 g)最大;QNS6家系的种厚(3.49 mm)最大,QNS10家系的种长(6.87 mm)最大。

表3 岭南槭3个种源18个家系种子形态变异
注:表中字母为Duncan多重比较结果,同列不同大写字母表示种源之间差异显著(P<0.05),同列不同小写字母表示种源内家系间差异显著(P<0.05);正负号(±)后数值表示标准误;下同。
Note: Letters in the table are the results of Duncan multiple comparisons. Significant differences among provenances are denoted by different capital letter(P<0.05),significant differences among families in the same provenance are denoted by different normal letters in the same column. Standard errors are denoted by data behind ‘±’; the same below.
3.2 种子萌发特征变异分析
3.3 幼苗地径、苗高生长差异分析
在试验期间,样本中有5个岭南槭家系的种子未发芽,故仅对剩余13个家系的幼苗生长情况进行分析。方差分析(表4)表明:HSD种源与NKS种源,除0.5年生幼苗的地径差异显著外,其它均差异不显著;HSD种源与QNS种源的0.5年生和1年生幼苗的地径、苗高和高径比均差异显著;QNS种源与NKS种源除0.5年生幼苗地径差异不显著外,其它均差异显著。幼苗0.5年生时,QNS种源的地径、苗高和高径比最大,分别比HSD种源增加26.38%、47.90%和25.57%,比NKS种源增加4.57%、38.68%和33.08%;1年生时,QNS种源的地径、苗高和高径比比HSD种源增加13.41%、49.77%和31.99%,比NKS种源增加20.0%、52.88%和25.04%。岭南槭幼苗从0.5年生到1年生时,HSD种源幼苗的地径增加了1.01倍,苗高增加了46.68%;QNS种源幼苗的地径增加了80.58%,苗高增加了48.53%;NKS种源幼苗的地径和苗高涨幅最小,分别为57.36%和34.73%。
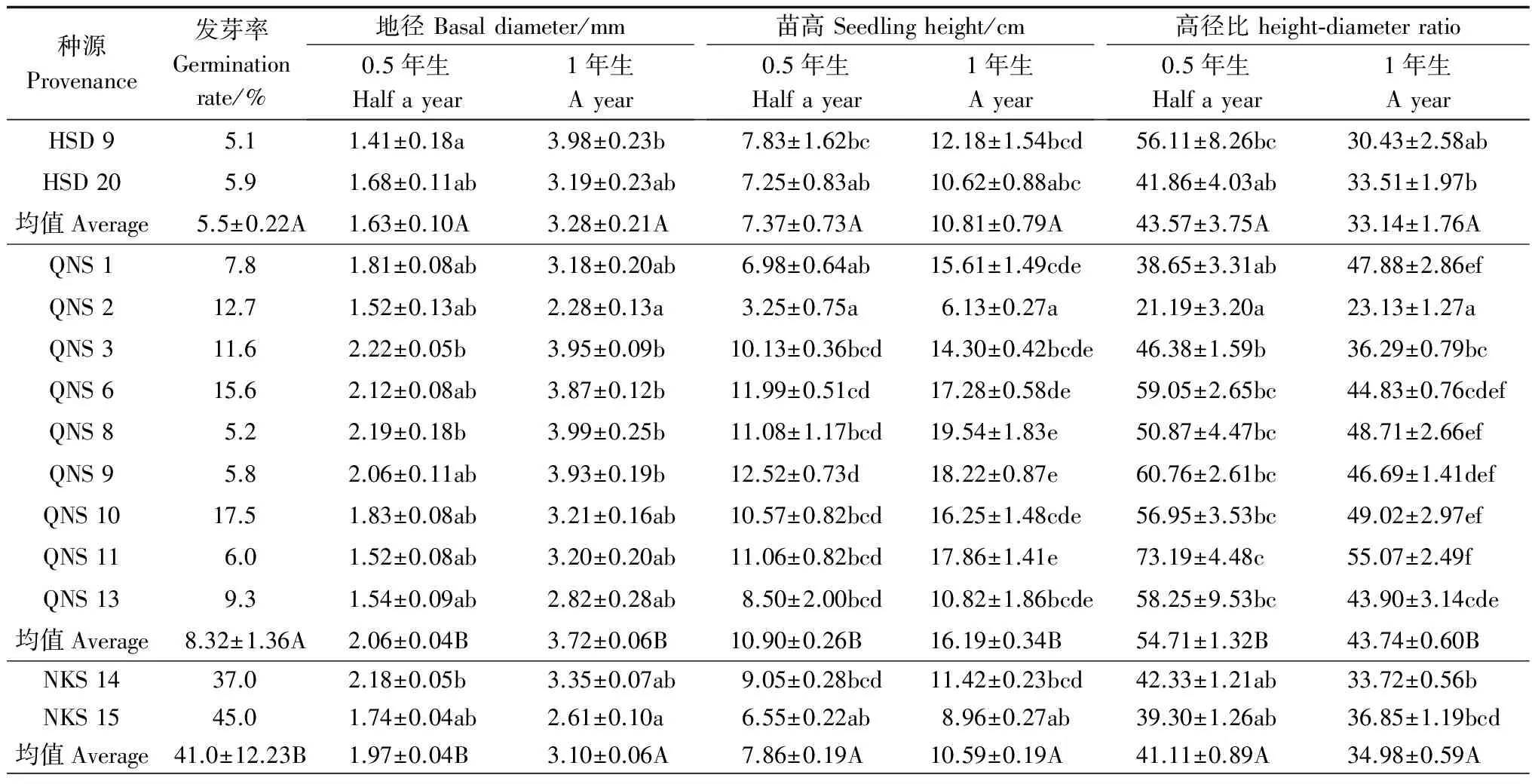
表4 岭南槭不同种源家系种子萌发和幼苗地径生长差异
岭南槭13个家系间,0.5年生、1年生幼苗的地径、苗高和高径比存在差异,说明各个采种家系间的生长表现差别较大。岭南槭幼苗0.5年生时,QNS3家系的地径最大,为2.22 mm,比地径最小的HSD9家系增加了57.45%;QNS9家系的苗高最大,为12.52 cm,比苗高最小的QNS2家系增加2.85倍。1年生时,QNS8家系的地径(3.99 mm)和苗高(19.54 cm)最大,分别比QNS2家系增加0.75倍和2.19倍。除QNS1和QNS2家系外,其他家系幼苗的高径比随着年龄的增加而下降。
3.4 幼苗叶色表现分析
从图1可看出:在明度L*水平,QNS13与其他12个家系均差异显著,QNS3与HSD9、QNS2、NKS14、NKS15家系差异显著;其中,QNS3、QNS13家系的明度L*值较大。在色相a*水平,HSD20、QNS2、NKS14和NKS15家系的色相a*值显著高于其他家系,其中,NKS14家系幼苗的色相a*值最大,QNS10家系的最小。在色相b*水平,QNS13家系幼苗的色相b*值显著高于其他家系,QNS3家系与HSD20、QNS1、QNS2、QNS9、QNS13、NKS14和NKS15家系的色相b*值差异显著,QNS2家系的色相b*值最小。
3.5 相关性分析
对岭南槭不同种源家系种子发芽、1年生幼苗生长与采种地的地理气候因子进行相关性分析,结果(表5)表明:种宽与种厚呈极显著正相关,与种长、翅长呈显著正相关;翅长与翅宽呈极显著正相关,翅长与年均高温呈显著正相关,与经度和年均温呈显著负相关;岭南槭种子发芽率与种子千粒质量呈显著正相关;地径与苗高呈极显著正相关,地径与海拔、纬度呈显著负相关;苗高与海拔、纬度、年均低温呈极显著或显著负相关,与经度呈显著正相关,与年均降水量呈极显著正相关。
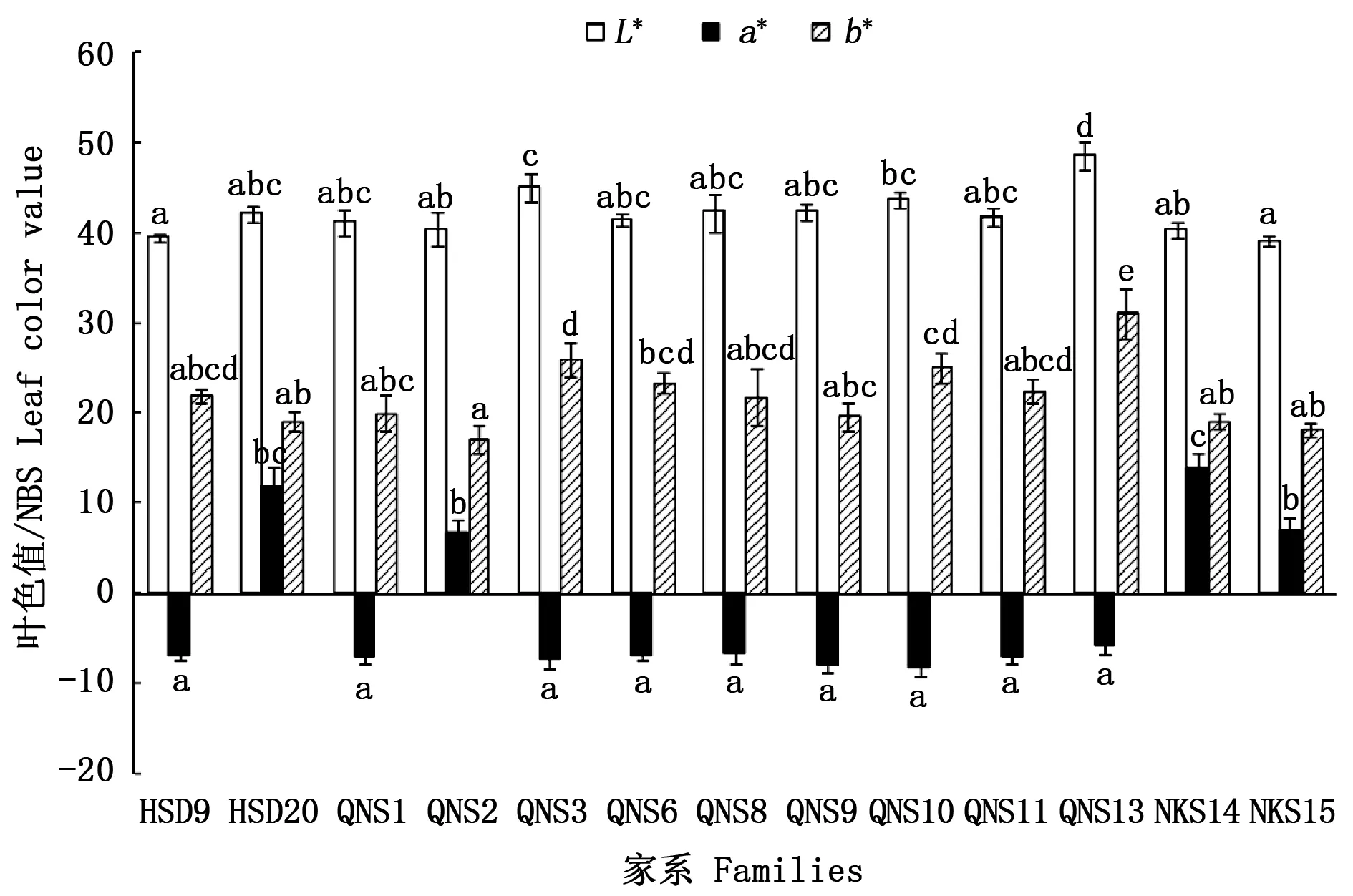
图中不同小写字母表示不同家系差异显著(P<0.05)Significant differences among families are denoted by different normal letter(P<0.05)图1 岭南槭不同家系幼苗叶色表现Fig.1 Differences of seedling leaf color value among families of Acer tutcheri
相关分析(表5)表明:翅长随着经度和年均温的升高而降低,随着年均高温的升高而增大;种子发芽率随着千粒质量的升高而升高。对于岭南槭1年生幼苗,地径随着海拔和纬度的升高而降低;苗高随着海拔、纬度和年均低温的升高而减小,随着经度和年均降水量的增大而增大。即自西向东,地径呈递增趋势,自南向北,地径、苗高有递减趋势。从高海拔到低海拔,幼苗的地径和苗高均逐渐增大;年平均降水量越高,幼苗地径和苗高也越大。
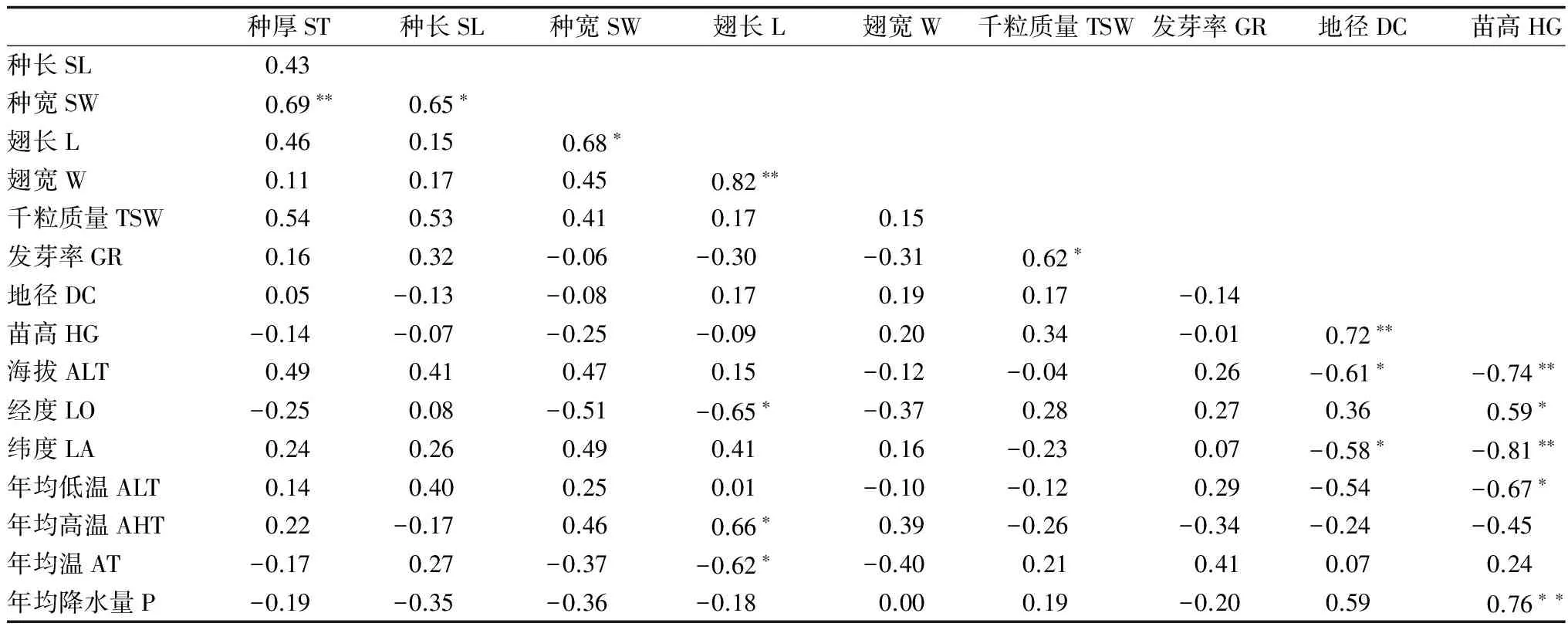
表5 岭南槭种子发芽和幼苗生长性状与种源地地理气候因子间的相关系数
注:*表示显著相关(P<0.05);**表示极显著相关(P<0.01)。
Note:*indicates reaching significant difference at 0.05 level;**indicates reaching highly significant at 0.01 level.
4 讨论
植物的果实和种子性状是比较稳定的遗传特征,在植物分类与遗传上有重要价值[15],种子表型变异研究对开展良种选育、种质资源保存具有十分重要的意义[16]。本研究中,翅长与年均高温显著正相关,与经度、年均温显著负相关,这表明种源地的经度和温度是影响种子翅长的重要生态因子,与Derakhshan等[17]的研究结果一致。在相同的环境条件下,岭南槭种子的发芽率与种子的千粒质量呈显著正相关,与林玲等[18]对云杉属植物种子的研究及赵曦阳等[19]对梓树属植物种子的研究结论一致;但也有研究表明,班克木属种子的发芽率与种子千粒质量相关不显著[20]。岭南槭千粒质量低的种子发芽率比较低,这可能与部分槭属植物种子出现种胚发育不良现象有关[21]。鉴于岭南槭种子的发芽率比较低,可通过筛选岭南槭种子千粒质量性状,选择发芽率高的种子。
对岭南槭幼苗生长研究结果表明,岭南槭1年生幼苗地径与苗高成正比,与姚小华等[22]的研究结果相同。幼苗0.5年生至1年生,绝大多数种源家系幼苗的地径涨幅大于苗高的涨幅,这与幼苗0.5年生至1年生阶段高径比减小的数据吻合。从岭南槭幼苗的生长性状与种子形态、种源地地理气候因子间的相关性看,幼苗的苗高和地径与种子形态的各个性状相关性不显著,说明种子形态并不影响幼苗的生长。岭南槭幼苗的地径生长与产地海拔、纬度呈显著负相关;苗高与产地的经度、年均降水量呈显著或极显著正相关,与海拔、纬度和年均低温呈极显著或显著负相关,说明岭南槭幼苗的生长受种源地地理气候因子的影响明显[23-24];所以,研究种源地的地理和气候因子与幼苗生长的关系,对岭南槭优良种源家系的进一步选择具有重要意义。
5 结论
本研究对不同种源家系岭南槭种子形态、发芽特征和幼苗生长性状进行研究,分析了不同种源家系岭南槭种子的表型性状、幼苗生长与种源地地理气候因子之间的相关关系。研究发现,经度和温度是影响种子翅长的重要因子;种子千粒质量与发芽率呈显著正相关。岭南槭种子发芽率从高到低依次为:种源间NKS > QNS > HSD,家系间(前5)NKS15>NKS14>QNS10>QNS6>QNS2。1年生幼苗的地径、苗高与海拔、纬度成反比;苗高与经度、年均降水量成正比,与年平均低温成反比。不同种源间,1年生地径、苗高生长由高到低为:QNS > HSD > NKS;不同家系间,岭南槭1年生幼苗地径生长由高到低(前5)为:QNS8>HSD9>QNS3>QNS9>QNS6,苗高生长由高到低(前5)为:QNS8>QNS9>QNS11>QNS6>QNS10。在色彩表现方面,QNS3、QNS13家系的明度L*值较大,QNS13色相b*值显著高于其他家系,NKS14家系幼苗的色相a*值最大,NKS14家系幼苗红叶特征最明显。
[1] 黄锦荣, 钟奕灵. 优良景观树种岭南槭及其栽培技术[J]. 南方农机, 2016,47(8):31-32.
[2] 中国科学院中国植物志编辑委员会. 中国植物志(第46卷)[M]. 北京: 科学出版社, 1981:66.
[3] Kurimoto S, Sasaki Y F, Suyama Y,etal. Acylated triterpene saponins from the stem bark ofAcernikoense(Aceraceae)[J]. Chenical & Pharmaceutical Bulletin, 2016,64(7):924-929.
[4] Jiang W, Zhuang M, Han H,etal. Progress on color emerging mechanism and photosynthetic characteristics of colored-leaf plants[J]. Acta Horticulture Sinica, 2005,32(2):352-358.
[5] 陈 勇, 李芳东, 廖绍波, 等. 深圳市生态风景林彩叶植物资源调查[J]. 中南林业科技大学学报, 2012,32(8):12-17.
[6] Sokolowska K, Zagorska-Marek B. Symplasmic, long-distance transport in xylem and cambial regions in branches ofAcerpseudoplatanus(Aceraceae) andPopulustremula×P.tremuloides(Salicaceae)[J]. American Journal of Botany, 2012,99(11):1745-1755.
[7] 邱迎君, 祝志勇, 易官美. 槭树科植物的种质资源及其开发利用价值[J]. 安徽农业科学, 2014,42(12):3598-3599.
[8] 陈华玲, 彭火辉, 朱启东, 等. 3个槭树品种嫁接繁殖技术研究[J]. 江西科学, 2014,32(1):35-38.
[9] 林富平, 程雪梅, 刘济祥, 等. 9种槭属植物宁波地区夏适应性评价[J]. 现代园艺, 2014(4):10-11.
[10] 刘济祥, 李红怀. 岭南槭育苗试验研究[J]. 现代农业科技, 2007(20):15-17.
[11] 管帮富, 彭火辉, 陈华玲, 等. 江西四种野生观赏槭资源的收集和开发应用[J]. 现代园艺, 2010(6):18-20.
[12] 陈 勇, 孙 冰, 廖绍波, 等. 彩叶树种岭南槭早期生长及其美景度评价研究[J]. 广东林业科技, 2015,31(1):19-22.
[13] 杜鹏珍, 孙 冰, 陈 勇, 等. 班克木种子形态及发芽特征在种内与种间的变异[J]. 林业科学研究, 2013,26(4):433-437.
[14] 石柏林, 吴家森, 钟泰林. 6种槭树属植物种子特性及其发芽试验[J]. 浙江林业科技, 2006,26(3):38-40.
[15] Minuto L, Fior S, Roccotiello E,etal. Seed morphology in Moehringia, L. and its taxonomic significance in comparative studies within the Caryophyllaceae[J]. Plant Systematics and Evolution, 2006,262(3):189-208.
[16] Nwangburuka C C, Denton L A, Oyelana O. Genetic variability and heritability of vegetative, fruit and seed yield traits in fluted pumpkin (TelfairiaoccidentalisHook F)[J]. 2014,13(32):3262-3270.
[17] Derakhshan A, Gherekhloo J. Factors affecting Cyperus difformis seed germination and seedling emergence[J]. Planta Daninha, 2013,31(4):823-832.
[18] 林 玲, 段二龙, 罗 建. 云杉属植物种子形态及萌发特征的种间变异[J]. 南京林业大学学报:自然科学版, 2015,39(1):62-66.
[19] 赵曦阳, 王军辉, 张金凤, 等. 梓树属4个种种子表型性状和发芽特性的研究[J]. 西北农林科技大学学报:自然科学版, 2008,36(12):149-154.
[20] 杜鹏珍. 观赏植物班克木种子变异及幼苗生长与生理特征研究[D]. 北京:中国林业科学研究院, 2014:23-24.
[21] 陈香波, 刘 杨, 赵明水, 等. 极度濒危树种羊角槭的种胚发育与休眠解除[J]. 林业科学, 2017,53(4):65-73.
[22] 姚小华, 任华东, 孙银祥, 等. 樟树种源/家系苗期性状变异分析[J]. 林业科学研究, 1999,12(3):283-290.
[23] Rafii M Y, Arolu I W, Omar M H A,etal. Genetic variation and heritability estimation inJatrophacurcasL. population for seed yield and vegetative traits[J]. Journal of Medicinal Plants Research, 2012,6(11):2178-2183.
[24] 杨晓霞, 冷平生, 郑 健, 等. 暴马丁香不同种源种子和幼苗的表型性状变异及其与地理-气候因子的相关性[J]. 植物资源与环境学报, 2016,25(3):80-89.
[25] 葛雨萱, 王亮生, 周肖红, 等. 香山黄栌叶色和色素组成的相互关系及时空变化[J]. 林业科学, 2011,47(4):38-42.
[26] 张 敏, 黄利斌, 周 鹏, 等. 榉树秋季转色期叶色变化的生理生化[J]. 林业科学, 2015,51(8):44-51.
VariationofSeedMorphologyandSeedlingGrowthamongDifferentProvenancesandFamiliesofAcertutcheri
WUPei-pei,SUNBing,LUOShui-xing,CHENLei,YANWei-ming
(Research Institute of Tropical Forestry,Chinese Academy of Forestry,Guangzhou 510520,Guangdong,China)
ObjectiveTo detect the characteristics of seed morphology, seed germination and seedling growth, and analyze the correlation with geographic and climatic factors of provenance locations in different provenances ofAcertutcheri.MethodANOVA and correlation analysis were used to analyze the seed morphology, seed germination, seedling growth and leaf color value of 18 families of three provenances from natural distribution areas in Guangdong.ResultThe provenance from Zhaoqing (HSD) had the thickest and the widest seeds, the provenance from Shenzhen (QNS) had the thinnest and the narrowest seeds, the provenance from Huizhou (NKS) had the biggest length and 1 000-seed weight. The samara length and width showed extremely significant positive correlation (P<0.01). 1 000-seed weight and germination rate showed significant positive correlation (P<0.05). Seedling basal diameter of 1-year-oldA.tutcheriwas proportional to seedling height; the growth of seedling was influenced by geographic and climatic factors of provenance locations. On the aspect of leaf color, theL*value of the families QNS3 and QNS13 was bigger than that of the other families, the b*value of the family QNS13 was bigger than that of the others significantly, and the family NKS14 had the biggest a*value.ConclusionThe higher the 1 000-seed weight, the higher the germination rate. The basal diameter and height of the seedlings are influenced by geographic and climatic factors of provenance locations. Family NKS14 with better leaf color has the most obvious red color than the others.
Acertutcheri; seed morphology; seed germination; seedling growth; leaf color
10.13275/j.cnki.lykxyj.2017.06.019
2017-03-21
广东省林业科技创新项目“岭南槭良种选育研究与示范”(2015KJCK017);国家自然科学基金“岭南槭引种适应性及栽培区域选择研究”(31600449)
吴培培(1992— ),女(回族),在读硕士研究生,主要从事风景园林植物应用研究. E-mail:cleverwp@126.com
* 通讯作者.
S722.7
A
1001-1498(2017)06-1015-07
徐玉秀)
猜你喜欢
种子(2023年7期)2023-09-24 07:40:54
特产研究(2022年6期)2023-01-17 05:05:02
辽宁林业科技(2021年1期)2021-03-16 09:17:08
农技服务(2020年1期)2020-12-17 08:15:01
山东农业科学(2019年11期)2019-12-24 01:11:27
现代农业科技(2019年1期)2019-07-11 01:36:25
现代农业科技(2017年24期)2018-01-22 21:35:06
广西林业科学(2016年3期)2016-03-16 05:43:29
广东海洋大学学报(2015年4期)2016-01-13 08:39:30
听力学及言语疾病杂志(2015年5期)2015-12-24 01:47:04
