
不同竹龄毛竹冠层叶片PSⅡ叶绿素荧光特性的比较
2017-12-29 02:45温国胜
中南林业科技大学学报 2017年1期
朱 弘 ,温国胜 ,b
(浙江农林大学 a.林业与生物技术学院;b. 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
不同竹龄毛竹冠层叶片PSⅡ叶绿素荧光特性的比较
朱 弘a,温国胜a,b
(浙江农林大学 a.林业与生物技术学院;b. 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300)
为了解不同竹龄毛竹对于光能的动态响应,以亚热带常绿阔叶林优势树种——毛竹Phyllostachys edulis3种竹龄(2年生、4年生、6年生)的冠层叶片(2龄)为材料,利用便携式叶绿素荧光仪(PAM-2100)和相对叶绿素含量计(SPAD-502),研究不同竹龄毛竹冠层叶片光系统Ⅱ(PSⅡ)荧光参数特征。结果表明:3种竹龄毛竹在荧光参数日变化均值(NPQ、qN、ETR)、PSII可逆恢复趋势(Ft、Fo、Fm)和RLC曲线(α、ETRmax、Ik)皆为4年生>2年生>6年生,表明在相同环境条件下,4年生毛竹的综合光合性能最优;2年生毛竹冠层叶具有较高的SPAD、Fv/Fo、Fv/Fm,反映其具有较高的潜在光能利用率和耐环境胁迫能力;在6月(大年新竹展叶后期或成竹换叶末期),2龄竹叶SPAD值有随年龄增加而降低的趋势;相关分析表明,SPAD与Yield、Fv/Fo呈极显著正相关(P<0.01),与PAR、ETR、Ft呈极显著负相关(P<0.01),与NPQ呈显著负相关(P<0.05),表明SPAD值在研究毛竹叶绿素荧光过程中具有重要的参考价值;主成分分析进一步表明,PSⅡ叶绿素荧光参数涵盖了毛竹叶片光适应下的荧光产量和能量耗散、暗适应下的光能利用效率和相对电子传递速率3个主要方面,可以作为研究毛竹光合生理的有效探针。因此,可通过及时择伐6年生毛竹(或III度竹)以上老竹,适当提高4年生毛竹(或Ⅱ度竹)在竹林中的比重;同时,针对笔者所在样地,建议于6月换叶集中末期对竹林追施氮肥以促进后期生长,通过优化毛竹立竹结构和林肥管理来提高毛竹林经营综合效益。
毛竹;林冠层;叶绿素荧光;日变化;相关性分析;快速光响应曲线;竹龄结构;择伐
林冠是森林与外界环境相互作用最直接和最活跃的界面层,是当今生物多样性和全球气候变化研究的焦点。由于林冠结构和几何特征对太阳辐射和降水等因素的影响,冠层的能量传输和分配呈现较大差异[1-2]。作为中国森林资源重要组成部分,毛竹Phyllostachys edulis是我国分布面积最大,范围最广,开发利用程度最高,集经济、生态、社会效益于一体的笋材两用竹种[3]。随着林龄的增加,毛竹的生物量和养分含量不断变化,由于枝干高大,其光能利用效率及分层效应显著[4]。
叶绿素荧光(Chlorophyll fluorescence)技术是一种以光合作用理论为基础,研究和探测其光合生理状况及外界因子对其影响的植物活体测定和诊断技术,具有快速、灵敏和非破坏性等优点,可以反映环境因子对植物的影响以及植物生理功能在不同水平上的变化[5-7],与“表观性”的气体交换指标相比,具有“内在性”特征,被视为测定植物光合作用快速、无损伤的良好探针[8]。应用该项技术已经在毛竹的逆境生理、栽培管理[5]等方面得到了相应研究:毛竹幼苗叶片叶绿素荧光参数的异质性随水分胁迫程度的加深而加强[9];不同程度条件下都会对毛竹叶片光系统Ⅱ(PSⅡ)活动中心造成损伤,-10 ℃造成损伤不可逆,且与不同转录因子具有显著相关性[10];对毛竹种子不同剂量137Cs-γ辐射并测定实生幼苗叶片表明,低剂量能提高PSII反应中心的能量捕获效率和光合能力[11];不同氮素施肥水平条件下,毛竹叶片叶绿素荧光诱导曲线、反射光谱和荧光光谱与毛竹的生长具有良好相关性,叶绿素荧光诱导曲线可以作为快速、准确、无损的施肥检测方法[12]。
前人对毛竹的光合生理生态特性与外界环境因子之间的关系进行了广泛研究,但对毛竹冠层空间上不同竹龄的叶绿素荧光异质性研究报道较少。基于此,笔者以3 种不同竹龄毛竹为实验材料,通过测定其冠层叶片PSⅡ叶绿素荧光及环境相关参数,旨在了解毛竹不同竹龄对于环境尤其是光能的动态适应,为揭示毛竹的快速生长机制和毛竹林的科学经营、高效利用提供理论依据。
1 材料与方法
1.1 试验地概况
样地设在浙江省临安市青山镇研里村——浙江农林大学竹类研究所毛竹生理生态定位监测站(地理坐标30°14′N,119°42′E)。地形地貌为低山丘陵,研究样地的海拔为100~250 m。该区域属亚热带季风气候,年平均气温为15.9 ℃,年降水量为1 442.0 mm,毛竹林土壤发育于凝灰岩和粉砂岩的红壤土类,土层厚度约为 60 cm。pH 值为5.20~5.90,有机质为 3.97 g·kg-1,全氮、速效磷、速效钾分别为 0.14,4.25 和 78.16 mg·kg-1[13]。实验区所属为毛竹林地一般经营类型[14],密度约为3 200株·hm-2,林下尚生长少量灌木有白马骨Serissa serissoides、大青Clerodendrum cyrtophyllum、赛山梅Styrax confuses等,草本植物有鸡矢藤Paederia scandens、小果菝葜Smilax davidiana、寒苺Rubus buergeri、异叶蛇葡萄Ampelopsis humulifoliavar.heterophylla等。
1.2 试验材料
研究区毛竹林样地采取隔年留养新竹及采伐老竹的方式,依据成竹标记及毛竹换叶规律方法[15]表明现存竹龄结构为2年生(12竹)、4年生(10竹)、6年生(08竹)(见表1),并均已钩梢,选取有代表性的西南坡中部地段,搭建定位观测塔。毛竹的冠层中上部位于观测塔的8.5~10.0 m处。

表1 毛竹不同竹龄主要形态识别Table 1 The prime morphological identification of Phyllostachys edulis of different years old
1.3 PS II叶绿素荧光参数的测定
测定时间为2014年6月,从 7:00开始 到16:00结束,每间隔1 h 测定 1 次,共计10次,选取3种竹龄林冠的中上部向阳面枝条2龄生健康竹叶,每株随机选择4片进行测定。使用便携式调制式叶绿素荧光仪(PAM-2100,Walz,Germany),在光适应阶段即冠层叶自然光照强度下,测定毛竹3种竹龄叶片的PSⅡ实际光化学效率(Yield)、给定光强下的稳定荧光(Ft)、非光化学猝灭系数(qN)。在暗适应阶段中,供试毛竹叶片使用配套暗适应叶夹DLC-8遮光20 min以上,先以弱调制测量光(0.05 μmol·m-2s-1)诱导测得初始荧光(Fo),再照内源饱和脉冲(6 000 μmol·m-2s-1)测得最大荧光产量(Fm)、PSⅡ最大光化学量子产量(Fv/Fm)主要荧光参数。同时利用仪器自带光量子探头和热电偶记录光合有效辐射(PAR)及叶表面温度。以上所有参数重复4次,读取稳定数值作为该时刻的生理指标,每次测定随机更换叶片,避免叶片因叶夹长时间挤压。
根据已经测定的荧光参数继续计算:相对电子传递速率(electron transport rates,ETR,RET)RET=ΦPSII×PAR×0.5×0.84, 其 中,ΦPSII为 PSII实际光化学效率 , 由 (Fm′-Ft) /Fm′计算得出;Ft为稳态荧光,0.84为叶片的经验性吸光系数,0.5为光能在PSII和PSI两个光系统中分配的比例系数[16]。PSⅡ潜在活性(Fv/Fo)=(Fm-Fo)/Fo;非光化学淬灭系数(NPQ,NPQ)=Fm/Fm′-1,qN=1-(Fm′-Fo)/(Fm-F)[17]。
快速光响应曲线(RLC)根据Platt[8]的方法进行,光化学梯度设定从int1至int10,直到达到饱和光强。每个梯度持续时间为10 s,并打开饱和脉冲测得各梯度下毛竹叶片的表观电子传递速率(ETR)和Yield。
1.4 相对叶绿素含量(SPAD)的测定
相对叶绿素含量(SPAD)使用便携式叶绿素含量测定仪(SPAD-502,Japan)测定。随机选取6片完好的功能竹叶,在每张叶片中部避开中脉的区域均匀选取读取,平均值±SD作为该叶片的相对叶绿素含量。
1.5 数据处理
采用SPSS16.0对实验数据进行SPAD值的单因素方差分析(One -Way ANOVA)和最小显著差法(LSD)多重比较(α=0.05)、叶绿素荧光参数的相关分析与主成分分析;快速光响应曲线以非线 性回 归 经验 模 型P=Pm(1-e-αPAR/Pm)×e-βPAR/Pm[18],其中Pm代表无光抑制时的最大光合速率,即最大表观光合电子传递速率rETRmax;α为快速光曲线的初始斜率;β为光抑制参数,由Pm和α可以得到半饱和光强 Ik =Fm/α。采用Microsoft Excel 2003进行数据整理和图表绘制。
2 结果与分析
2.1 光合有效辐射及毛竹叶表面温度日变化
6月毛竹叶表面温度日变化规律呈平缓单峰曲线,在27.53~36.08℃之间变化;PAR日变化趋势则较为显著,自11:00到12:00迅速提高,并在14:00达到最高峰,随后又迅速下降(见图1)。
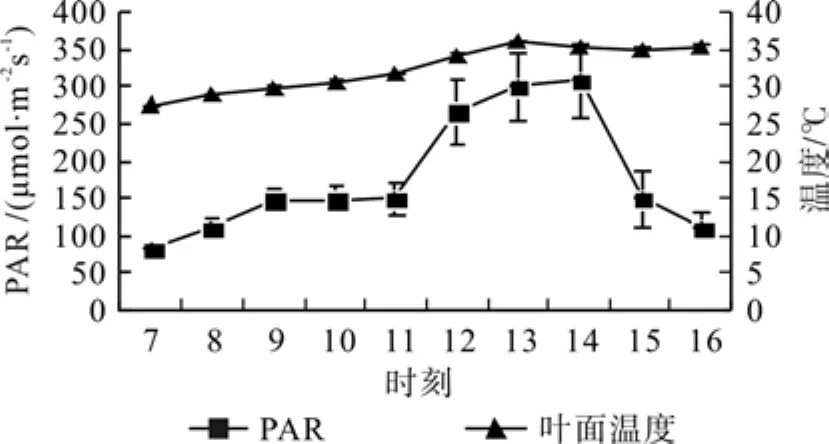
图 1 毛竹光合有效辐射及叶表面温度日变化Fig.1 Daily variation of PAR and leaf temperature in leaves of Phyllostachys edulis of different years old
2.2 毛竹不同竹龄PSⅡ荧光参数日变化特征
Ft(或Fs)为给定光强下的稳定荧光;Fo为初始荧光,是PSⅡ反应中心处于完全开放时的荧光产量;Fm为暗适应的样品照射饱和脉冲得到的最大荧光产量,是PSⅡ反应中心处于完全关闭时的荧光产量[17]。毛竹3种竹龄冠层的Ft、Fo、Fm日变化趋势基本一致且呈现明显的倒“V”型:10:00至11:00,3种竹龄迅速下降至最低点,12:00后又迅速上升;在上午时段,6年生>2年生>4年生,而下午时段则呈现相反趋势,4年生>2年生>6年生(见图2)。
NPQ与qN 表示 PSII反应中心天线色素吸收的光能不能用于光合电子传递而以热的形式耗散的部分,是一种自我保护机制[17]。在NPQ中,2年生毛竹从7:00至13:00逐步上升并达到峰值,随后下降;4年生毛竹上午迅速提升,在10:00达到峰值,14:00后维持平稳;不同的是6年生毛竹于8:00、12:00和15:00依次出现3个峰值。在qN中,2年生毛竹从7:00至12:00逐步上升,随后又平缓下降;而4年生毛竹9:00就达到其峰值,13:00之前均趋于平缓;6年生毛竹8:00出现峰值,随后下降并趋于平缓,至13:00达到第2峰值,之后又下降。日变化均值比较:4年生(2.790/0.904)>2年生(2.756/0.904)>6年生(2.343/0.886)(见图3)。
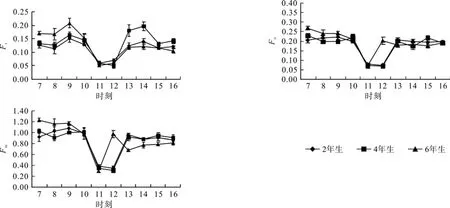
图2 毛竹不同竹龄Ft、Fo、Fm日变化Fig.2 Daily variation of Ft、Fo、Fm in leaves of Phyllostachys edulis of different years old
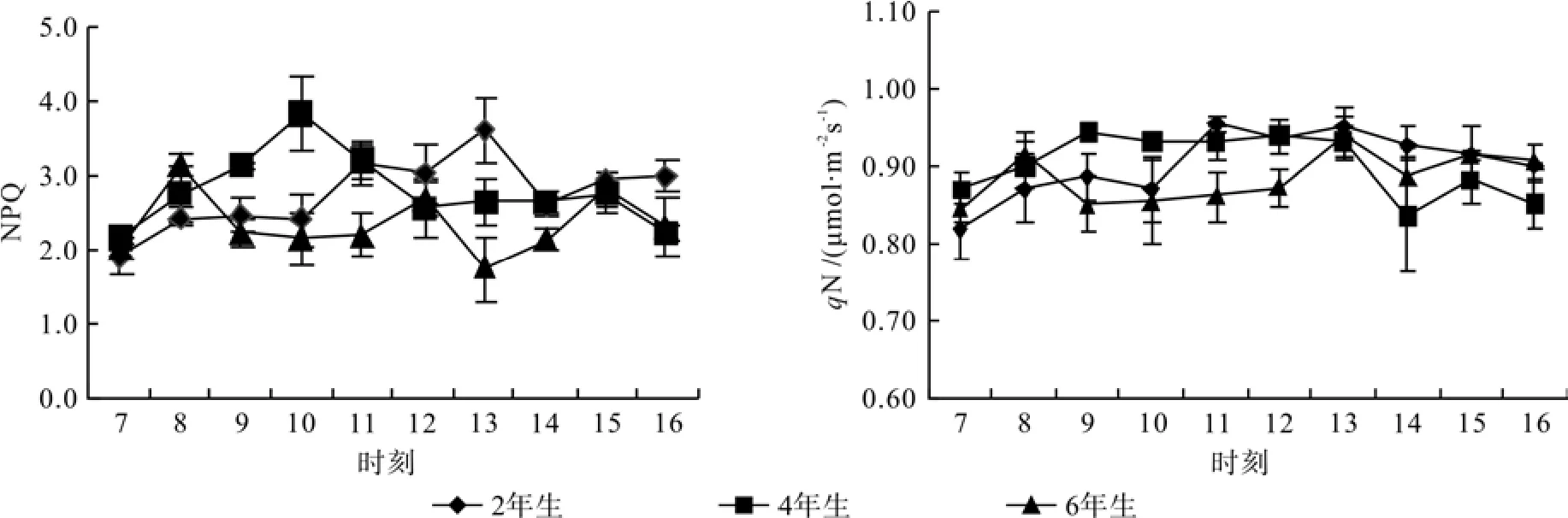
图 3 毛竹不同竹龄NPQ、qN日变化Fig.3 Daily variation of NPQ、qN in leaves of Phyllostachys edulis of different years old
Fv/Fm是PSⅡ最大光化学量子产量,常用于度量植物叶片PSⅡ原初光能转换效率,反映植物对光能的利用效率, 在非胁迫条件下,该指标不受物种和环境条件的影响[5]。3种竹龄毛竹冠层的Fv/Fm在一天中保持平稳趋势,但稍有区别:其中2年生毛竹最为稳定,并与4年生毛竹在12:00均达到最低值,而此时6年生毛竹陡增至一天中最大,随后下降至稳定水平。日变化均值比较:2年生(0.784)>4年生(0.780)>6年生(0.775)。
Fv/Fo是PSⅡ潜在活性。2年生毛竹上午平缓升高,到10:00达到峰值,其后便平缓下降,直至15:00稍有上升;4年生毛竹在9:00就达到峰值,随后便呈波动下降;相较而言,6年生毛竹生日变化波动较大,分别在9:00、12:00和15:00出现3个峰值,在11:00及13:00出现低值。日变化均值比较,2年生(3.612)>4年生(3.581)>6年生(3.473)。
Yield也称为PSⅡ的实际光化学效率,反映植物叶片用于光合电子传递的能量占所吸收光能的比例[19]。3种竹龄毛竹冠层Yield日变化图均呈现“V”型,与PAR及叶表面温度呈相反趋势;毛竹4年生、6年生有着较好的一致性,8:00以后,6年生Yield值均大于4年生,最低值均出现在12:00;而2年生毛竹最低值则推迟至14:00,而后迅速升高至正常水平。日变化均值比较:6年生(0.450)>2年生(0.472)>4年生(0.362)。
ETR是反映实际光强条件下的表观电子传递效率。3种竹龄毛竹基本呈“M”型,其中4年生毛竹和6年生毛竹的日变化走势一致,两者均在10:00达到第一峰值,并在11:00迅速降至最低点,随后两者迅速提升,6年生毛竹在13:00达到其第二峰值,4年生毛竹继续缓慢增长,15:00达到其第二峰值。2年生毛竹在9点就已经达到其第一峰值,12:00过后迅速提高,13:00达到最大峰值。日变化均值比较:4年生(29.69)>6年生(22.80)>2 年生 (22.24)。
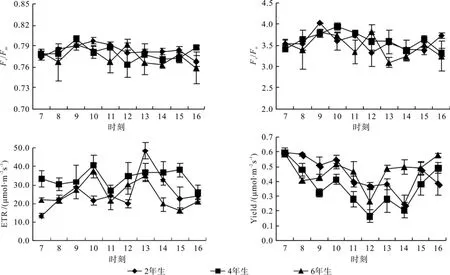
图 4 毛竹不同竹龄Fv/Fm、Fv/Fo、ETR、Yield日变化Fig.4 Daily variation of Fv/Fm、Fv/Fo、ETR、yield in leaves of Phyllostachys edulis of different years old
2.3 叶片相对叶绿素含量
便携式叶绿素含量测定仪(SPAD-502,Japan)被证明具有快速、便捷、无损的优点,已经被广泛使用于农、林等领域,其SPAD值与叶绿素含量具有显著的相关性, 适用于植物不同季节、年龄、营养状况和基因型[20-22],能较好地反映叶绿素含量的变化趋势,是测定活体状态下色素含量的有效工具。
6月SPAD值最高是2年生毛竹(42.23±0.50),其SPAD与4年生毛竹(39.28±1.03)和6年生毛竹(38.52±0.89)均达到显著差异(P<0.05),分别高出7.51%和9.63%,但其余2个竹龄差异不显著(P>0.05)(见图5) 。
2.4 3种竹龄毛竹叶绿素荧光参数及其环境因子的相关性分析
毛竹叶片PAR与ETR、Fo、Ft,Yield与SPAD,ETR 与Ft,Fo与Ft、Fm,Fv/Fo与SPAD均呈极显著正相关(P<0.01),其中,PAR与ETR正相关系数最大,为0.959,这说明ETR受PAR影响最大;PAR与Yield、SPAD,SPAD与Ft、ETR均呈极显著负相关(P<0.01),其中PAR与Yield负相关系数最大,为-0.960,说明Yield受PAR影响最大(见表2)。
PAR与NPQ,Yield与Fv/Fo,ETR与NPQ、Fo,NPQ与Fo均呈显著相关(P<0.05);PAR与Fv/Fo, Yield与ETR、NPQ, NPQ与SPAD均呈负相关(见表2)。
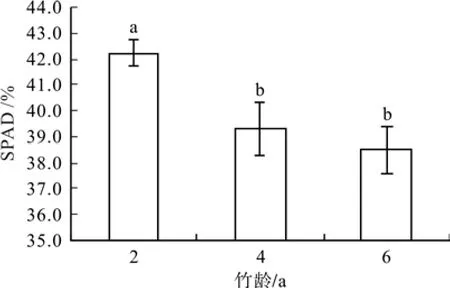
图 5 不同竹龄毛竹相对叶绿素含量(SPAD值)比较Fig.5 Comparison of SPAD in leaves of Phyllostachysedulis of different years old
2.5 不同竹龄毛竹间PSⅡ快速光响应曲线比较
2.5.1 PAR与ETR的关系
在低光下,3种竹龄毛竹的ETR无明显差异,但随着光强的增强,ETR迅速产生分化并逐渐趋向饱和(见图6)。由快速光响应曲线得出3种竹龄毛竹ETR变化初始斜率,可以用来表示光化学反应的启动速率。不同林龄毛竹光化学反应的启动初始斜率(α)为4年生>2年生>6年生;当光化学反应启动后,快速光响应曲线升高并趋于饱和,ETRmax为4年生>2年生>6年生;半饱和光强(Ik)反映了植物对强光的耐受能力,其大小为4年生>2年生>6年生(见表3)
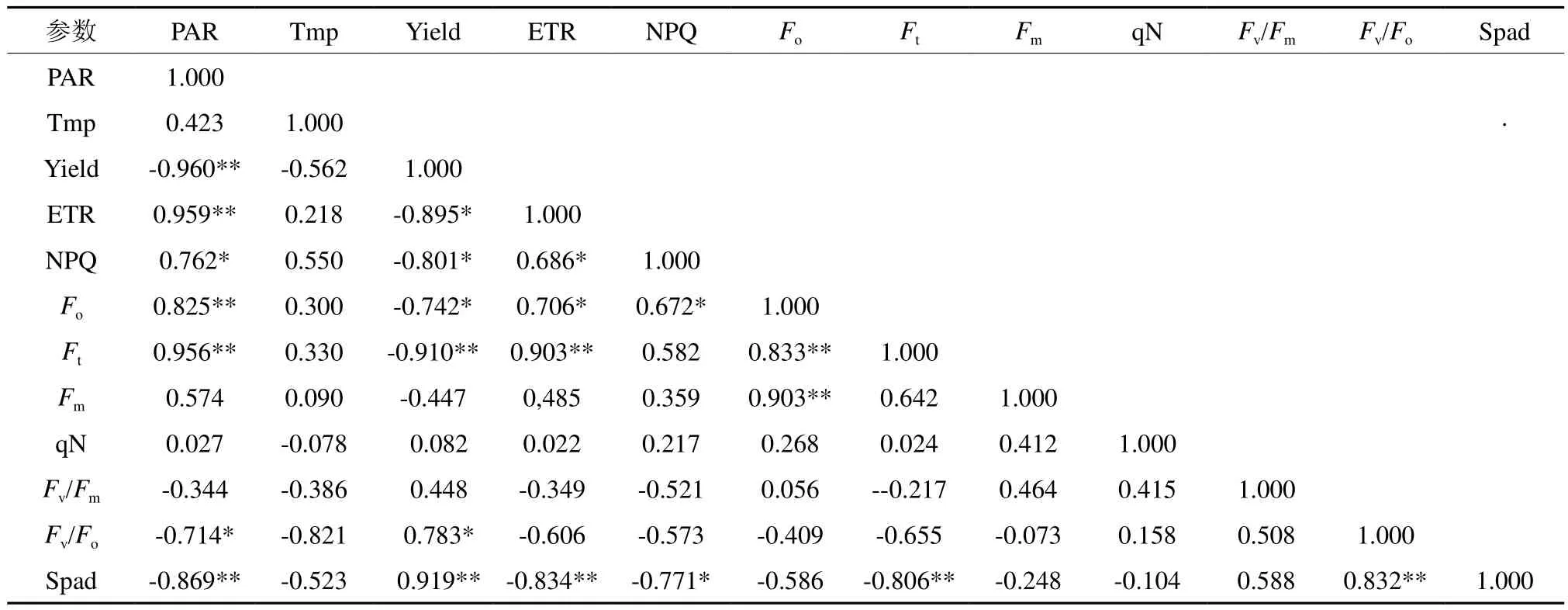
表2 毛竹不同竹龄叶绿素荧光参数及其环境因子的相关性分析†Table 2 Analysis of the correlation of chlorophyll fluorescence parameters and environmental factors of Phyllostachys edulis of different years old
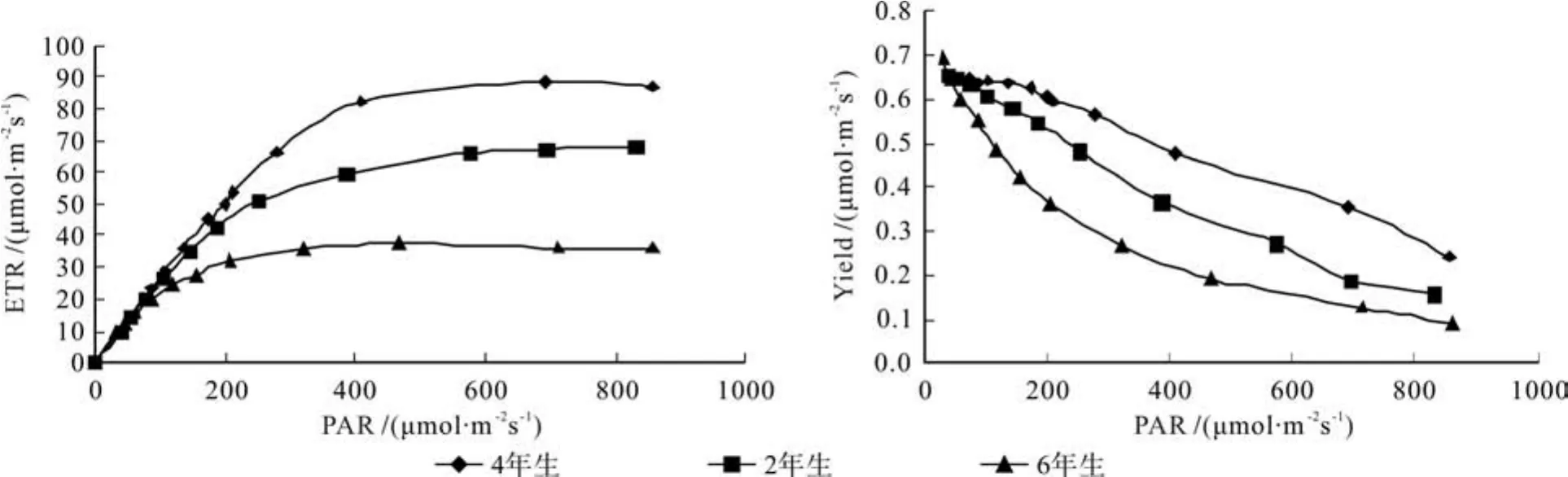
图6 不同竹龄毛竹快速光响应曲线的比较Fig.6 Comparison of RLC in leaves of Phyllostachys edulis of different years old
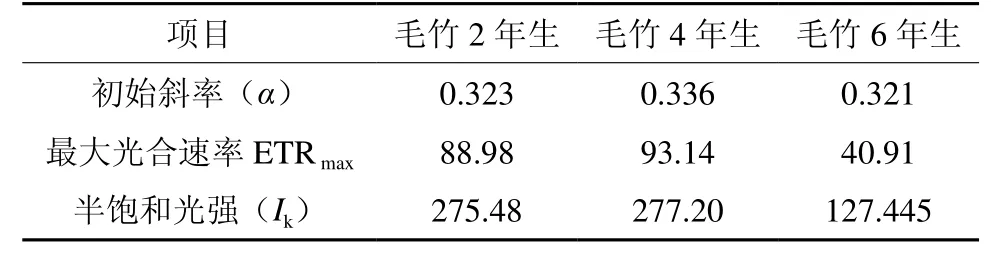
表3 不同竹龄毛竹ETR初始斜率与 ETR max 的比较Table 3 Comparison of initial slops of response of ETR to light and ETRmax in leaves of Phyllostachys edulis of different years old
2.5.2 PAR与Yield的关系
与前者相反,随着光强的升高,3种竹龄毛竹Yield逐渐下降,且差异显著,毛竹有效量子产量下降率,进而表明毛竹耐光抑制斜率(绝对值),其大小为4年生(0.698)>2年生(0.672)>6年生(0.626)(见图6)。
2.6 3种竹龄冠层叶毛竹PSⅡ叶绿素荧光参数的主成分分析
进一步对毛竹PSⅡ叶绿素荧光参数进行主成分分析,按照特征根大于1的原则提取了3个主要成分,其累计贡献率达到86.905%,具有分析价值。第1主成份的特征根为4.030,方差贡献率为44.773%,并在Fm、Fo、Ft、Yield上具有较大正载荷,在qN上有较大负载荷,主要反映光适应下荧光产量和能量耗散;第2主成分的特征根为2.257,方差贡献率为25.081%,其中在Fv/Fm、Fv/Fo上有较大的正载荷,主要反映暗适应下植物PSⅡ对光能的利用效率;第3主成分的特征根为1.535,方差贡献率为17.051%,对ETR有较大的正载荷,主要反映相对电子传递速率(见图7)。
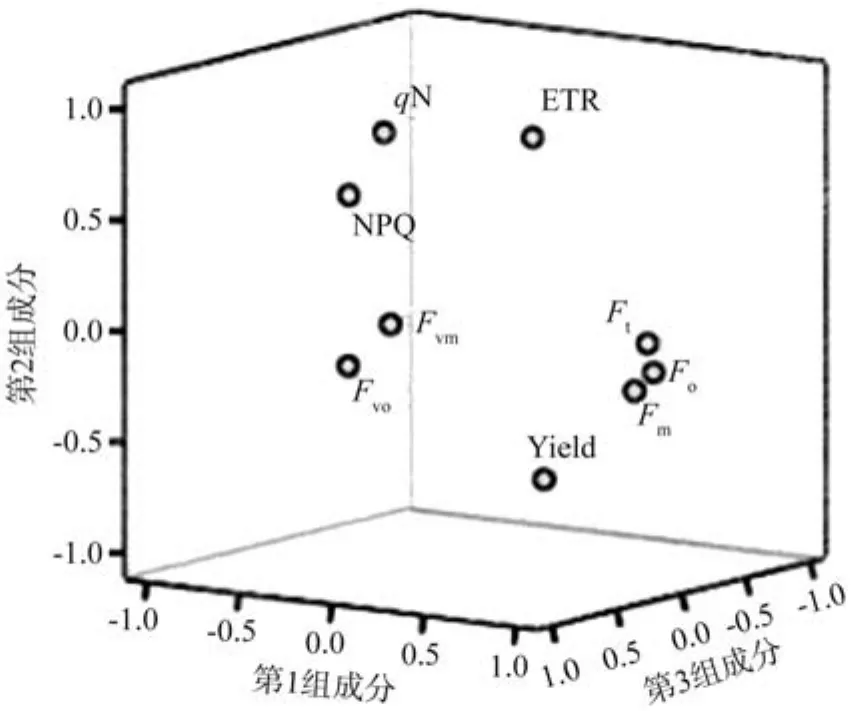
图7 不同竹龄毛竹荧光参数的主成分分析Fig.7 Principal component analysis of PSⅡchlorophyll fluorescence in leaves of Phyllostachys edulis of different years old
3 结论与讨论
PSⅡ位于叶绿体类囊体膜上,是光抑制的原初位点和主要作用部位[23],其代表参数常用于评价光合机构的功能和环境胁迫的影响[24]。3种竹龄毛竹Yield日变化动态显示:6年生较2年生和4年生具有较高的光能转化效率,这为暗反应的光合碳同化积累更多所需的能量。非光化学淬灭系数一般有qN或NPQ两种表示方法,综合两者日变化规律,即4年生>2年生>6年生,表明4年生毛竹能够最大程度通过增加光能耗散来减少光抑制的发生,具有较强的光适应能力;应用叶绿素荧光对毛竹的研究也证实了用NPQ比qN能更准确地反映无性系的非光化学淬灭的情况,这与张守仁[17]的研究结论一致。
Ft代表给定光强下的稳态荧光,Fo与PSⅡ受激发后的电子密度相关,其变化可以推测反应中心的状况和可能的光保护机制[5]。Fm可反映通过PSⅡ的电子传递情况[17]。Yield是 PSⅡ反应中心部分关闭时的光化学效率,可作为植物叶片光合电子传递速率快慢的相对指标[19]。Ft、Fo、Fm及Yield 4个荧光参数日变化趋势一致均为倒“V”型,表明各竹龄都不同程度地出现了光抑制现象,其中Ft、Fo、Fm上午时段与下午时段呈明显的相反趋势,原因可能是受到正午的高光、高温的胁迫,毛竹冠层叶PSII天线出现短暂可逆失活,进而开启非辐射耗散机制消耗过剩光能以保护光合机构。从PSII可逆恢复趋势(即下午时段)看,Ft、Fo、Fm三者均表现为毛竹4年生>2年生>6年生,这与非光化学淬灭系数(NPQ、qN)的规律一致。而Yield日变化均值与Ft、Fo、Fm上午时段趋势一致,表现为6年生>2年生>4年生,这可能与PSII部分可逆恢复周期长短有关。3种竹龄Fv/Fm日变化动态显示:除12:00毛竹2年生、4年生低于6年生竹龄外,其余数值均为2年生>4年生>6年生,且变化平稳,表明在强光、高温、大气干旱等综合因子胁迫下,2年生和4年生毛竹比6年生毛竹具有更好的应对环境耐胁迫再调整能力,Fv/Fo日均变化趋势也再次证明毛竹PSⅡ潜在活性2年最好,4年生其次,6年生最差。而ETR日变化均值为4年生>2年生>6年生,表明在相同光强下,4年生、2年生毛竹较6年生所形成的活跃化学能(ATP和 NADPH)更大,为光合碳同化提供更多的能量和还原能力。
叶绿素含量是反映植物衰老状况和光合能力的一个重要指标,SPAD值能较好地反映植物叶片叶绿素含量及含氮的变化趋势。相关分析表明:SPAD与Yield、Fv/Fo呈极显著正相关(P<0.01),与PAR、ETR、Ft呈极显著负相关(P<0.01),与NPQ呈显著负相关(P<0.05),表明SPAD值在研究毛竹叶绿素荧光过程中具有重要的参考价值。2年生毛竹(2龄叶)SPAD值比4年生和6年生分别高出7.51%和9.63%,表明快速生长期幼龄毛竹具有更高的捕获光能进行光合作用的能力,且SPAD值随竹龄增长有不断降低的趋势,这与封焕英等[25]的结论一致,却与陈嘉琦等[4]的结论相反,这可能与研究时间、叶龄不一致及SPAD值处于动态变化有关。
PSⅡ快速光响应曲线(RLC)利用ETR随PAR变化规律,反映了植物叶片对光强的响应,蕴含丰富的信息,是研究植物光合能力的一种重要手段[26]。在同样的生存环境下, 毛竹光能的利用效率和耐光抑制能力均为4年生>2年生>6年生。
利用主成分分析法进一步研究表明,光适应下的荧光产量和能量耗散、暗适应下的光能利用效率、相对电子传递速率3个主要方面能够较好地反映所测毛竹3种竹龄冠层PSⅡ叶绿素荧光参数的大部分信息,可以作为研究毛竹光合生理的有效探针。本实验发现,3种竹龄毛竹荧光参数日变化均值NPQ、qN、ETR,PSII可逆恢复程度(Ft、Fo、Fm),RLC曲线(α、ETRmax、Ik)最高,皆为4年生毛竹>2年生>6年生,表明在相同环境条件下,4年生毛竹的综合光合性能最优,因而是毛竹林生物量形成的主体,2年生其次,6年生最差;而2年生毛竹在SPAD、Fv/Fo、Fv/Fm均值最高,说明测量期间幼龄毛竹生长旺盛,富含较高的叶绿素含量,并具有潜在的光化学效率和耐环境胁迫的能力,因而是竹林更新的后备力量。
人工经营措施会显著影响毛竹林生产力水平[27-28],其中合理的林分结构对毛竹的光合效能及其产量有着极为重要的作用[29],不同年龄的立竹结构在生活力、碳密度[30]能力不同,造成对毛竹群落更新及生物量的影响极为显著。有研究表明,丰产合理的竹林立竹结构为Ⅰ度竹、Ⅱ度竹、Ⅲ度竹,各占总株数的30%左右, IV度竹占10%以下[3],并且Ⅱ度竹的比重显著影响当年产笋量[31]。施肥可显著提高Ⅰ度竹和Ⅱ度竹的氮含量,且对Ⅲ度竹氮含量影响不显著[25]。就笔者研究的青山样地而言,测量期6月是毛竹新竹展叶形成期也是成竹集中换叶的末期,表明该时期毛竹2龄叶片的SPAD值呈随竹龄增长而明显降低趋势。综上所述,从竹林集约经营的角度,应适当提高4年生毛竹(或Ⅱ度竹)在竹林中的比重,及时择伐6年生毛竹(或III度竹)以上老竹,同时建议于毛竹春季换叶集中末期对青山毛竹林样地追施氮肥以促进后期生长,通过优化毛竹立竹结构和林肥管理来提高毛竹林经营综合效益。
[1] 李 勇,韩海荣,康峰峰,等. 油松人工林冠层光合生理特性的空间异质性[J]. 东北林业大学学报, 2013, 41(4): 32-35.
[2] 孟令曾,张教林,曹坤芳,等. 迁地保护的4种龙脑香冠层叶光合速率和叶绿素荧光参数的日变化[J]. 植物生态学报,2005, 29(6): 110-118.
[3] 马乃训,赖广辉,张培新,等. 中国刚竹属[M]. 浙江: 浙江科学技术出版社, 2013: 360.
[4] 陈嘉琦,温国胜,王艳红,等. 春季毛竹光化学效率空间异质性比较[J]. 福建林学院学报, 2014, 34(1): 52-56.
[5] 温国胜,田海涛,张明如,等. 叶绿素荧光分析技术在林木培育中的应用[J]. 应用生态学报, 2006, 17(10): 1973-1977.
[6] Kumar R, Pal M, Teotia U V S. Diurnal changes in chlorophyll fluorescence in four species of bamboo[J]. Journal of Bamboo and Rattan, 2002, 1(4): 341-349.
[7] 杨柳青,张 柳,廖飞勇,等. 低温胁迫对南美天胡荽光合作用和叶绿素荧光的影响[J]. 经济林研究, 2014, 32(4): 99-102.
[8] White A J, Critchley C. Rapid light curves: A new fluorescence method to assess the state of the photosynthetic apparatus[J].Photosynthesis Research, 1999, 59(1): 63-72.
[9] 应叶青,郭 璟,魏建芬,等. 水分胁迫下毛竹幼苗光合及叶绿素荧光特性的响应[J]. 北京林业大学学报, 2009,31(6):128-133.
[10] 李 想,史世京,曹 颖,等. 冷冻胁迫下毛竹叶绿素荧光参数变化及抗寒相关转录因子表达[J]. 福建林学院学报, 2014,34(1): 57-63.
[11] 桂仁意,刘亚迪,郭小勤,等. 不同剂量137Cs-Y辐射对毛竹幼苗叶片叶绿素荧光参数的影响[J]. 植物学报, 2010,45(1):66-72.
[12] 高培军. 氮素施肥对毛竹光合能力与光谱特性的影响[D]. 北京: 北京林业大学, 2013.
[13] 董大川. 毛竹冠层叶片光谱及叶绿素荧光特性研究[D]. 临安:浙江农林大学, 2011.
[14] 陈存及. 毛竹林分密度效应的初步研究[J]. 福建林学院学报,1992, 12(1): 98-104.
[15] 熊文愈,周芳纯,胡长龙. 毛竹竹株年龄的确定方法[J]. 林业科学, 1965, 10(2): 87-88.
[16] Force L, Critchley C, Rensen J J V. New fluorescence parameters for monitoring photosynthesis in plants[J]. Photosynthesis Research, 2003, 78(1): 17-33.
[17] 张守仁. 叶绿素荧光动力学参数的意义及讨论[J]. 植物学通报, 1999, 16(4): 444-448.
[18] Ralph P J, Gademann R. Rapid light curves: a powerful tool to assess photosynthetic activity[J]. Aquatic Botany, 2005, 82(3):222-237.
[19] 钟泰林,李根有,石柏林. 3种浙江特产濒危植物气体交换特征和叶绿素荧光特性研究[J]. 上海交通大学学报:农业科学版, 2009, 27(2): 149-152, 176.
[20] Hawkins T S, Gardiner E S, Comerl G S. Modeling the relationship between extractable chlorophyll and SPAD-502 readings for endangered plant species research[J]. Journal for Nature Conservation, 2009, 17(2): 123-127.
[21] 鱼 欢,祖 超,杨建峰,等. 应用SPAD叶绿素仪测定不同位置胡椒叶片的SPAD值[J]. 热带作物学报, 2012, 33(10):1890-1895.
[22] 张日清,何小燕,叶 航,等. 油茶叶片SPAD值差异的初步研究[J]. 中南林业科技大学学报, 2011, 31(3): 30-33.
[23] Murata N, Takahashi S, Nishiyama Y,et al. Photoinhibition of photosystem II under environmental stress [J]. Biochimica Et Biophysica Acta, 2006, 1767(6): 414-421.
[24] 刘春英,陈大印,盖树鹏,等. 高、低温胁迫对牡丹叶片PSⅡ功能和生理特性的影响[J]. 应用生态学报,2012,23(1):133-139.
[25] 封焕英,范少辉,苏文会,等. 竹林专用控释肥对毛竹地上生物量和氮利用率的影响[J]. 生态学杂志,2014,33(9):2357-2362.
[26] 柏 祥,陈开宁,古小治,等. 水深梯度对荆三棱(Scirpus yagara)生长的影响[J]. 应用与环境生物学报,2011,17(1):34-38.
[27] 朱强根,金爱武,唐世刚,等. 毛竹枝叶生物量的冠层分布对钩梢和施肥的响应[J].中南林科技大学学报,2015,35(1):24-29.
[28] 陈卫东,谷战英,陈建华,等. 不同抚育措施对毛竹成竹和胸径地影响[J]. 经济林研究, 2014, 32(1): 168-170.
[29] 杨迪蝶,黄启民,高爱新. 毛竹冠层各层次叶片光合速率的变化[J]. 林业科学研究, 1988, 1(2): 217-223.
[30] 漆良华,刘广路,范少辉,等. 不同抚育措施对闽西毛竹林碳密度、碳贮量与碳格局的影响[J]. 生态学杂志,2009,28(8):1482-1488.
[31] 何 林,何小勇. 毛竹笋用林合理竹龄结构及其笋期生长规律[J]. 竹子研究汇刊, 2011, 12(3): 52-56.
Comparison of PSⅡchlorophyll lluorescence characteristic at canopy layers in different agedPhyllostachys edulis
ZHU Honga, WEN Guoshenga,b
(a. School of Forestry and Biotechnology; b. Cultivation Base for State Key Laboratory of Subtropical Silviculture, Zhejiang A & F University, Lin’an 311300, Zhejiang, China)
A study on daily variation and Characteristic of PSⅡchlorophyll fluorescence at canopy layers in leaves of Phyllostachys edulis of three different years old, a dominant bamboo species in subtropical evergreen broadleaved forest of China were conducted. The parameters were measured by portable chlorophyll fluorometer (PAM-2100) and chlorophyll meter (SPAD-502) on a fixed observation tower in Qingshan Mountain Linan, Zhejiang Province. The results showed that Mean daily variation of fluorescence parameters(NPQ,qN, ETR), PSII reversible recovery ability (Ft,Fo,Fm) and rapid light curve (α, ETRmax, Ik) were in the same order: 4 years old bamboo>2 years old bamboo>6 years old bamboo, indicated that under the same conditions, 4 years old bamboo had optimum photosynthetic performance followed with 2 years old bamboo and 6 years old bamboo; 2 years old bamboo recorded high value in SPAD,Fv/Fo,Fv/Fm, Re flected its higher potential energy utilization and the ability of resistance to environmental stresses; There was a tendency that leaf SPAD of 2 years old bamboo were decreased with increasing age in rapid growth period; Correlation Analysis showed that the SAPD had high signi ficant positive correlation with Yield andFv/Fo(P<0.01), had high signi ficant negative correlation with PAR, ETR andFt(P<0.01), and also had signi ficant negative correlation with NPQ (P<0.05),demonstrated that the SPAD index had important reference value in the process of the research on bamboo chlorophyll fluorescence; Principal Components Analysis further suggested PSⅡchlorophyll fluorescence parameters involving fluorescence Yield and energy dissipation under light adaptation,light use ef ficiency under dark adaptation and relative electron transfer rates in three main aspects may be used as an effective probe in research of Photosynthetic Physiology.Therefore,from the perspective of intensive management of Phyllostachy edulis,proportion of 4 years old bamboo (or II degree bamboo) should be enhanced properly,selective cutting above 6 years old bamboo in time (or Ⅲdegree bamboo), meanwhile, additional nitrogenous fertilization applied in June, in order to promote the growth of late stage. All in all,Through optimizing the structure of bamboo standing and application of fertilization to enhance the bene fits of moso bamboo forest management comprehensively.
Phyllostachys edulis; canopy layer; PSⅡchlorophyll fluorescence; correlative analysis; daily variation; rapid light curve;age structure; selective cutting
S718.43;S795.7
A
1673-923X(2017)01-0012-08
10.14067/j.cnki.1673-923x.2017.01.003
2015-06-20
国家自然科学基金(31270497);省院合作项目(2014SY16)
朱 弘,硕士研究生 通讯作者:温国胜,教授; E-mail: wgs@zafu.edu.cn
朱 弘,温国胜. 不同竹龄毛竹冠层叶片PSⅡ叶绿素荧光特性的比较[J].中南林业科技大学学报,2017, 37(1): 12-19.
[本文编校:谢荣秀]
猜你喜欢
作物杂志(2022年3期)2022-07-06
中国农业信息(2022年1期)2022-05-25
农业机械学报(2021年11期)2021-12-07
阅读(科学探秘)(2020年8期)2020-11-06
意林·少年版(2020年13期)2020-08-02
东坡赤壁诗词(2020年2期)2020-06-04
绿色科技(2019年2期)2019-05-21
阅读与作文(小学高年级版)(2019年3期)2019-04-20
干旱地区农业研究(2019年6期)2019-02-25
中国果业信息(2019年1期)2019-01-05
