
干旱胁迫对蚬木幼苗表型可塑性的影响
2017-12-20 03:40庞世龙欧芷阳申文辉郝海坤彭玉华
中南林业科技大学学报 2017年5期
庞世龙,欧芷阳,申文辉,郝海坤,彭玉华
(广西壮族自治区林业科学研究院,广西 南宁 530002)
干旱胁迫对蚬木幼苗表型可塑性的影响
庞世龙,欧芷阳,申文辉,郝海坤,彭玉华
(广西壮族自治区林业科学研究院,广西 南宁 530002)
以3年生蚬木Excentrodendron hsienmu幼苗为试验材料,采用盆栽控水法,设置4个水分处理梯度,模拟研究不同强度干旱胁迫下蚬木幼苗生长和形态的表型可塑性。结果表明:干旱胁迫对蚬木幼苗地上部分的抑制作用明显大于地下部分;随着干旱胁迫程度的增加,蚬木幼苗地上部分形态指标均呈下降的趋势,而地下部分根系先升高后降低,在中度胁迫处理条件下达到峰值;各组分生物量积累显著降低,其中,叶生物量对干旱胁迫的反应更为敏感,变化的幅度最大,茎次之,根最小;干旱胁迫下,植株将更多的同化物分配到根系,根生物量比增加,根冠比增大,增强了植株抵御干旱胁迫的能力;综合分析表明,土壤含水量11.23%~14.44%是蚬木幼苗生长、生物量积累受到明显抑制的阈值,在育苗和造林实践中可将该值作为土壤水分含量的下限。
蚬木;生物量;干旱胁迫;表型可塑性
中国西南喀斯特地区出露碳酸盐岩面积达54万km2,是全球喀斯特碳酸盐岩出露面积最广、最集中,喀斯特地貌发育最强烈,景观类型最丰富,生态环境最脆弱,人地矛盾最尖锐的地区[1-3]。其独特的地质背景和二元水文结构导致“地高水低、雨多地漏、石多土少、土薄易旱”,生境多样,普遍存在着不同程度的水分亏缺,植物难以获得充足的水分,生长发育受到限制,植被一旦遭受破坏极难恢复,水分是喀斯特地区植被恢复与重建的关键性限制因子。筛选适生植物,利用植物的抗旱能力重建退化喀斯特生态系统已成为植物学和生态学研究的热点之一[4]。
蚬木Excentrodendron hsienmu属椴树科Tiliaceae蚬木属常绿大乔木,是中国最重的木材,有“木中之王”的美称,广泛分布于广西右江以南至左江流域的喀斯特山地,是广西北热带喀斯特季节性雨林的优势种和关键种之一,常与肥牛树Cephalomappa sinensis、石山樟Cinnamomum saxatile、菲律宾朴树Celtis philippensis、青冈栎Cyclobalanopsis glauca或大苞藤黄Garcinia bracteata等组成该区域特有的、最具代表性的原生性植被[5],承载着水源涵养、土壤保持、生物多样性保护等维持生态平衡、保障区域生态安全的重要功能。长期以来,由于乱砍滥伐和缺乏有效保护,蚬木种群数量急剧下降,分布面积日渐萎缩,趋于濒危,现已被列为国家Ⅱ级保护植物[6]。野外调查发现,因人为大量砍伐导致蚬木生境退化,种群天然更新不良,加强蚬木野生种群的保护与研究迫在眉睫。目前对蚬木的研究主要集中在生物学特性、种群动态和群落结构等方面[7-10],而关于蚬木对环境因子水分胁迫的生理生态适应性研究鲜有报道。为此,本研究以3年生蚬木实生苗为研究对象,设置4个水分处理梯度,模拟研究不同强度干旱胁迫对蚬木幼苗生长和表型可塑性的影响,探寻蚬木幼苗对干旱环境的适应机制,以期为珍稀濒危树种蚬木的保育、推广应用及植被演替更新提供理论依据。
1 材料与方法
1.1 试验材料
选取长势基本一致、生长健壮的3年生蚬木实生苗作为试验材料。供试土壤为喀斯特山地棕色石灰土,土壤容重为1.36 g·cm-3,田间持水量为32.08%,pH值为7.30,有机质含量为53.90 g·kg-1,全氮含量为 3.77 g·kg-1,全磷含量为 3.84 g·kg-1,全钾含量为 13.16 g·kg-1,碱解氮含量为253.96 mg·kg-1,速效磷含量为 30.26 mg·kg-1,速效钾含量为 98.97 mg·kg-1。
1.2 试验设计与处理
2015年7月15日盆栽定植,塑料花盆规格为口径31 cm×底径20 cm×高28 cm,每盆栽植1株。将盆栽移入可控试验大棚,苗木适应生长2个月后,每盆浇水至饱和,待土壤自然落干后进行水分胁迫试验。试验设置4个持续性干旱胁迫处理水平:(1)正常水分处理(对照,CK),为田间持水量的65%~75%;(2)轻度胁迫(LS),为田间持水量的50%~60%;(3)中度胁迫(MS),为田间持水量的35%~45%;(4)重度胁迫(SS),为田间持水量的20%~30%。与之相对应的盆内土壤实际质量含水量分别为20.85%~24.06%、16.04%~19.25%、11.23%~14.44%、6.42%~9.62%。每处理30盆,共120盆。试验期间,为使各处理稳定在设定的标准水分范围内,采用美国产Thetaprobe- MI2X型土壤水分探头监测土壤含水量的变化,及时补充盆内蒸发散失的水分。
1.3 测定指标及方法
2016年1月5日试验结束,每处理随机选取5株,用自来水冲洗根部和地上部分,除去泥土,再用去离子水冲洗干净,吸干表面水分。计算每株须根数、分枝数和叶片数;用钢卷尺(精度0.1 cm)测量株高、分枝长和根长;用游标卡尺(精度0.01 mm)测量基径粗;用电子天平(精度0.001 g)称量根、枝、叶鲜质量,然后将样品放置烘箱中105 ℃杀青30 min,再于80℃烘至恒质量。称量干质量并按植株计算下列参数:叶生长量比(LMR,叶生物量/整株生物量)、枝生物量比(BMR,枝生物量/整株生物量)、根生物量比(RMR,根生物量/整株生物量)、叶/地上比(L/S,叶生物量/地上总生物量)、枝/地上比(L/S,枝生物量/地上总生物量)、根冠比(R/S,根生物量/地上生物量)[11]。
1.4 数据处理
采用SPSS 21.0软件对试验数据进行统计分析,采用单因素方差分析(One-Way AVOVA)和Duncan多重比较法,比较不同干旱胁迫强度下蚬木幼苗的生长和形态响应差异。采用Origin 9.0软件绘制图形。
2 结果与分析
2.1 水分处理对蚬木叶片水分状况的影响
叶片含水量能够很好地反映植株水分状况与萎蔫之间的平衡关系,抗旱性强的品种,叶片失水率低,含水量下降速率较小,能保持较好的水分平衡。蚬木幼苗在对照、轻度胁迫、中度胁迫和重度胁迫处理下的叶片含水量分别为(57.22±0.45)%、(55.42±1.18)%、(51.56±1.12)%、(50.82±1.06)%,与对照相比较,叶片含水量分别下降了3.15%、9.38%和10.42%。对照及轻度胁迫分别与中度胁迫、重度胁迫之间存在显著差异(p<0.05),其他处理间无显著差异。这可能是因为蚬木叶片革质,具有发达的角质层,从而避免了水分过多散失,同时也表明蚬木幼苗具有很强的耐旱性,可塑性强,适应范围广。
2.2 形态特征对干旱胁迫的响应
干旱胁迫显著降低了蚬木幼苗的株高(F=6.181,p=0.008)、基径(F=7.685,p=0.004)、叶片数(F=5.372,p=0.010)和累加分枝长(F=3.775,p=0.038)的生长,干旱胁迫程度越高,对其生长的抑制作用越显著(见图1)。与对照相比较,在轻度胁迫、中度胁迫和重度胁迫处理条件下株高分别降低了6.64%、20.70%和23.4%;基径降低了1.17%、8.72%、20.72%;叶片数降低了4.55%29.55%、50.00%;累加分枝长降低了2.28%、34.53%、49.84%。四者对照与轻度胁迫之间均无显著差异,这进一步表明蚬木幼苗具有一定的抗旱能力。蚬木幼苗的株高、叶片数和累加分枝长在中度胁迫和重度胁迫处理下均差异不显著,这可能是因为蚬木幼苗在中度胁迫处理时,土壤中可利用的水分已经小于株高、叶片数和累加分枝长的生长所必需的水分阈值所致。虽然干旱胁迫对蚬木幼苗基径生长的抑制作用显著,但显著性仅在重度胁迫处理下显现,这表明干旱胁迫对蚬木幼苗基径生长的抑制作用有限。主根长(F=1.360,p=0.302)和一级须根数(F=4.186,p=0.030)呈现先升后降的趋势,在中度胁迫处理下达到最大值,表明中度胁迫可促进蚬木幼苗根系的生长,主根长和一级须根数量增加,有利于扩大根系表面积,增强吸水能力,抵御干旱胁迫。
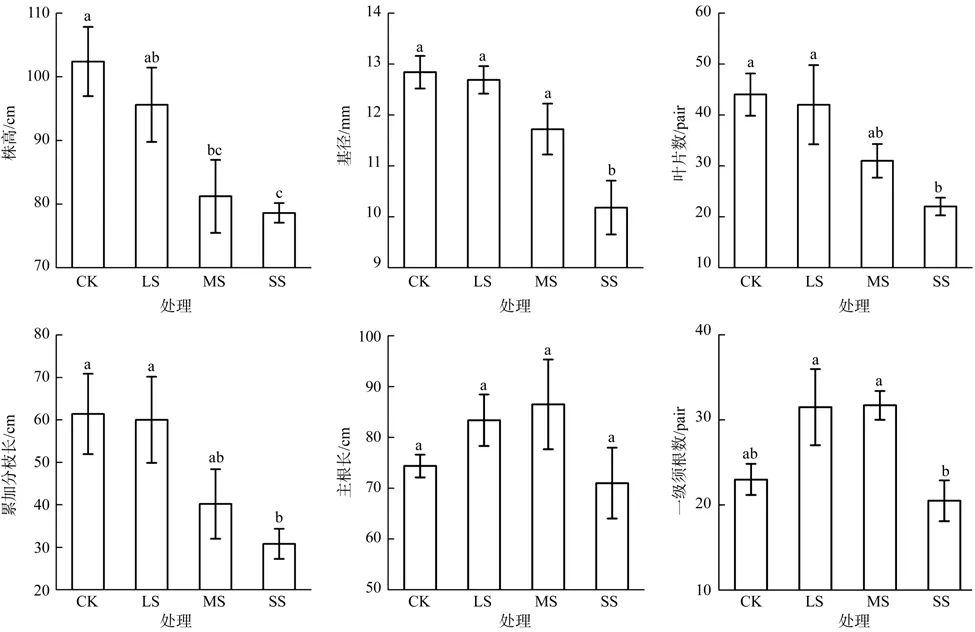
图1 干旱胁迫对蚬木幼苗形态特征的影响Fig. 1 Effect of drought stress on the morphological characteristics of Excentrodendron hsienmu seedlings
2.3 生物量积累与分配对干旱胁迫的响应
干旱胁迫对蚬木幼苗叶生物量(F=24.050,p<0.001)、茎生物量(F=22.736,p<0.001)和根生物量(F=3.518,p=0.041)的积累产生了显著影响,其中叶生物量对轻度胁迫反应较敏感(见图2)。与对照相比较,在轻度胁迫、中度胁迫和重度胁迫处理条件下叶生物量分别降低了18.34%、40.61%和58.64%,茎生物量降低了5.35%、33.61%、50.10%,根生物量降低了8.20%、17.25%、47.18%。叶生物量各处理之间均存在显著差异(p<0.05),干旱胁迫程度越高,叶生物量积累越少,这表明蚬木幼苗在干旱胁迫下通过新生叶形变小、成熟叶凋落的方式,主动减少蒸腾耗水,应对干旱胁迫。茎生物量的对照与轻度胁迫之间无显著差异,但轻度胁迫、中度胁迫和重度胁迫之间均存在显著差异(p<0.05),表明蚬木幼苗通过降低茎的生长量和生物量积累来适应不同程度的干旱胁迫。根生物量的对照及轻度胁迫分别与重度胁迫之间存在显著差异(p<0.05),其他处理间无显著差异,干旱胁迫使蚬木幼苗的主根长和一级须根数量增加,但其生物量并没有增大,表明随着干旱胁迫程度的加剧,主根和一级须根变得细而长,从而使其更好地吸收深层土壤的水分,积极抵御干旱胁迫。
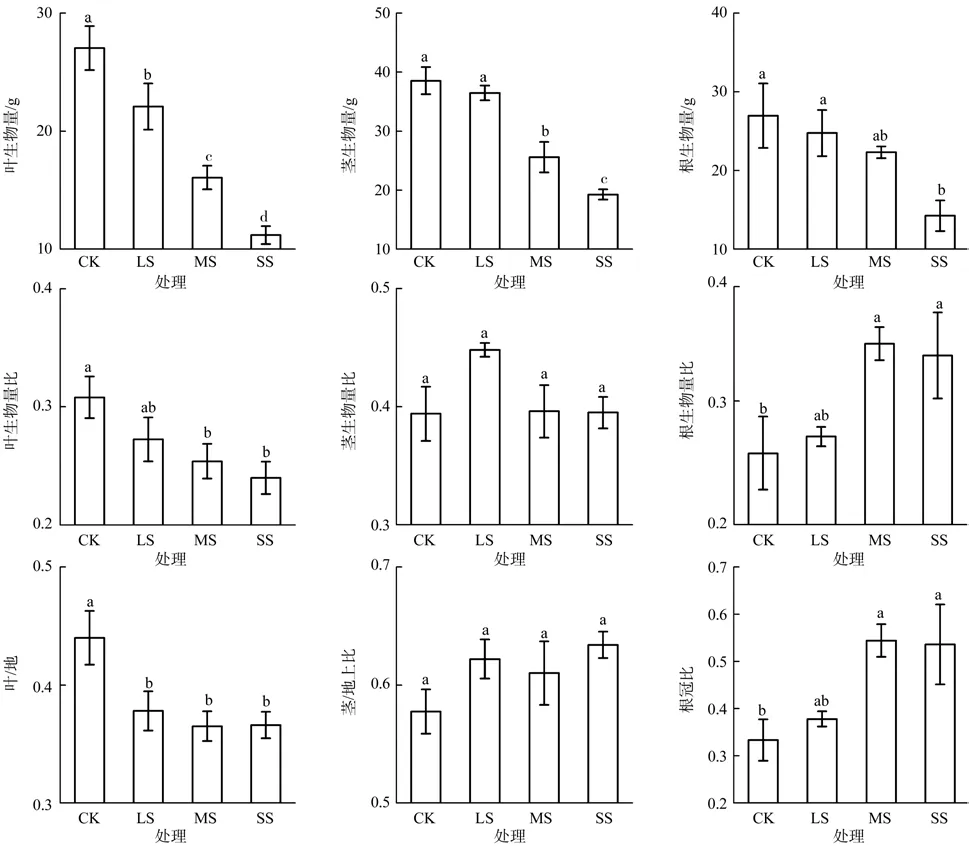
图2 干旱胁迫对蚬木幼苗生物量积累和生物量分配的影响Fig. 2 Effect of drought stress on the biomass accumulation and allocation of Excentrodendron hsienmu seedlings
生物量分配方式随之发生变化。随着干旱胁迫程度的加剧,叶生物量比(F=3.761,p=0.032)、叶/地上比(F=4.570,p=0.018)显著降低,根生物量比(F=3.503,p=0.044)、根冠比(F=3.693,p=0.038)显著增加,而茎生物量比(F=2.250,p=0.125)、茎/地上比(F=1.442,p=0.270)差异不显著。由此可见,干旱胁迫对蚬木幼苗地上部分的抑制作用明显大于地下部分,蚬木幼苗通过减小地上部分的生长量,主动降低消耗,提高根冠比来适应干旱环境。
3 结论与讨论
表型可塑性是指同一基因的生物个体对不同环境应答而产生不同表型的特征[13-14],是植物适应异质生境的主要对策之一[14-17]。本研究中,蚬木幼苗在不同干旱胁迫强度下植株形态和生长指标存在显著差异(p<0.05),表明蚬木具有较高的表型可塑性,对生境具有较强的耐受性和适应性,适宜在干旱的环境中生长。
随着干旱胁迫强度的加剧,蚬木幼苗的株高、基径、叶片数和累加分枝长等地上部分形态指标显著降低,而主根长和一级须根数呈先升后降的趋势,在中度胁迫处理条件下达到峰值,然后下降。这表明干旱胁迫首先抑制了地上部分的生长,蚬木幼苗通过减小地上部分的生长量,主动降低消耗来适应干旱环境。相比而言,叶片对干旱胁迫的响应更为敏感,表现为新生叶缓慢、单叶面积减小、成熟叶凋落等,使得冠层叶面积减小,光合作用下降,最终导致幼苗生长率与生产力下降[17-18]。
大量研究表明,植物生长受到限制时,会通过调节其生长和生物量分配来适应环境的变化[4,14-20,25],而其生物量就成了表征植物对胁迫适应性的重要指标[12]。本研究中,蚬木幼苗各组分生物量均随土壤含水量的减少呈下降的趋势,这是因为干旱胁迫破坏了植物水分吸收与蒸腾之间的动态平衡,气孔关闭,蒸腾速率和光合速率下降,单位叶面积的同化物减少,进而导致干物质积累受阻,生物量增量下降。随着干旱胁迫的加剧,叶生物量积累变化的幅度最大,茎次之,根最小。这主要是当土壤水分较长时间不能满足植物蒸腾需要时,叶片萎蔫甚至凋落,以减少水分散失;同时植株将更多的同化物分配到根系生长,主根长和一级须根数量增加,有利于扩大根系表面积和增强吸水能力,抵御干旱胁迫。这是植物对逆境的一种适应性反应[4,14-25],即在有限的土壤水分条件下,通过降低蒸腾表面积、增加根冠比来提高水分及养分的利用效率。
综合分析表明,不同强度的干旱胁迫对蚬木幼苗水分状况、形态特征和生物量积累产生了显著影响。在土壤含水量为20.85%~24.06%时,幼苗生长、水分利用效率和生物量积累均最大;土壤含水量为16.04%~19.25%时,仅对幼苗的株高、叶面积等产生轻度抑制;而土壤含水量11.23%~14.44%是幼苗生长受到明显抑制的干旱胁迫强度阈值,成熟叶凋落,生物量分配格局也发生了显著变化;土壤含水量为6.42%~9.62%时,幼苗生长受到了严重抑制,成熟叶大量凋落但植株并未死亡,表明蚬木幼苗具有较强的干旱胁迫适应能力。面对不同强度的干旱胁迫,蚬木幼苗表现出了形态和生理多种适应机制,如通过关闭气孔或叶片凋落,减小水分蒸腾散失,生物量分配向地下部分的比例增加,以增强对水分的获取能力,保持水分蒸腾与吸收之间的动态平衡,保证异质环境下的正常生长。显然,土壤含水量11.23%~14.44%是蚬木幼苗干旱胁迫的最小适应阈值,在育苗和造林实践中可将该值作为土壤水分含量的下限,采取施放保水剂、覆盖地膜等技术措施,控制土壤含水量不低于11.23%~14.44%的水平,以提高造林成活率。本研究的结论是基于蚬木幼苗外部形态、生物量积累与分配等表型特征对干旱胁迫的响应结果,还不能完全说明蚬木对干旱的适应机制及形态策略,尚需结合植物形态学、生理学和解剖学特征开展系统的研究[4,12,25],进行综合评价。
[1]广西壮族自治区地方志编篡委员会编.广西通志——岩溶志[M]. 南宁:广西人民出版社,2000.
[2]卢 峰.溶土地现状与石漠化治理模式探析[J].广西林业科学 ,2012,41(2):183–185.
[3]农胜奇.广西岩溶地区石漠化危害及治理对策[J].广西林业科学,2007,36(3):170-172.
[4]吴丽君,李志辉.不同种源赤皮青冈幼苗生长和生理特性对干旱胁迫的响应[J].生态学杂志,2014,33(4):996-1003.
[5]李先琨,苏宗明,吕仕洪,等.广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义[J].山地学报,2003,21(2):129-139.
[6]梁建平.广西珍稀濒危树种[M].南宁:广西科学技术出版社,2001.
[7]袁铁象,欧芷阳,苏志尧,等.桂西南喀斯特山地蚬木野生种群结构与动态[J].中南林业科技大学学报,2013,33(3):1-8.
[8]向悟生,王 斌,丁 涛,等.喀斯特季节性雨林蚬木种群结构和数量动态[J].生态学杂志,2013,32(4):825-831.
[9]欧芷阳,苏志尧,彭玉华,等.桂西南喀斯特山地蚬木幼龄植株的天然更新[J].应用生态学报学报,2013,24(9):2440-2446.
[10]欧芷阳,朱积余,曹艳云,等.蚬木生存群落木本植物与土壤和地形因子的耦合关系[J].生态学杂志,2013,32(12):3182-3189.
[11]Hunt R. Plant growth analysis[M]. London: Edward Arnold,1978.
[12]孙洪刚,刘 军,董汝湘,等.水分胁迫对毛红椿幼苗生长和生物量分配的影响[J].林业科学研究,2014,27(3):381-387.
[13]Pigliucci M. Phenotypic plasticity: beyond nature and nurture[M].Baltimore and London: The Johns Hopkings University Press, 2001.
[14]徐 军,朱雅娟.干旱胁迫对不同种源油蒿幼苗的生长和形态可塑性的影响[J].东北林业大学学报,2015,43(10):55-57.
[15]卓 露,管开云,李文军,等.不同生境下细叶鸢尾表型可塑性及生物量分配差异性[J].生态学杂志,2014,33(3):618-623.
[16]徐 飞,郭卫华,徐伟红,等.刺槐幼苗形态生物量分配和光合特性对水分胁迫的响应[J].北京林业大学学报,2010,32(1):24-30.
[17]李芳兰,包维楷,吴 宁,等.白刺花幼苗对不同强度干旱胁迫的形态与生理响应[J].生态学报,2009,29(10):5407-5416.
[18]罗永忠,李 广.土壤水分胁迫对新疆大叶苜蓿的生长及生物量的影响[J].草业学报,2014,23(4):213-219.
[19]林武星,谷 凌,朱 炜,等.干旱胁迫对台湾海桐生长和生理生化特性的影响[J].草业科学,2014,31(10):1915-1922.
[20]龚吉蕊,黄永梅,葛之葳,等.4种杂交杨对土壤水分变化的生态学响应[J].植物生态学报,2009,33(2):387-396.
[21]钟飞霞,王瑞辉,李 婷,等.土壤水分对油茶果实主要经济指标的影响[J].经济林研究,2015,33(4):32-37.
[22]梁文斌,聂东伶,吴思政,等.水分胁迫对短梗大参生理生化特征的影响[J].经济林研究,2016,34(3):99-104,186.
[23]刘全勇,卢 锟,李 泽,等.水分胁迫对刨花润楠幼苗生长及光合特性的影响[J].中南林业科技大学学报,2016,36(9):29-35,43.
[24]Sala A, Piper F, Hoch G. Physiological mechanisms of drought induced tree mortality are far from being resolved[J]. New Phytologist, 2010,186(2):274-281.
[25]张中峰,尤业明,黄玉清,等.模拟喀斯特生境条件下干旱胁迫对青冈栎苗木的影响[J].生态学报,2012,32(20):6318-6325.
Effect of drought stress on phenotypic plasticity ofExcentrodendron hsienmuseedlings
PANG Shilong, OU Zhiyang, SHEN Wenhui, HAO Haikun, PENG Yuhua
(Guangxi Zhuang Autonomous Region Forestry Research Institute, Nanning 530002, Guangxi, China)
An experiment was conducted to explore effect of drought stress on biomass allocation and phenotypic plasticity of 3-yearold pottedE. hsienmuseedlings. The seedlings grown under four soil water stressed conditions, of field moisture capacity, which were imposed by controlling the water supply. Results showed that inhibitory effect of drought stress on above-ground ofE. hsienmuseedlings was stronger than that on under-ground. All above-ground morphological indexes ofE. hsienmuseedlings decreased as the soil drought stress increasing, while the under-ground ascended firstly then descended, with the greatest under MS condition. Biomass of each group significantly decreased. Among them, response of leaf biomass to drought stress was more sensitive. Variation range of leaf biomass was the highest, that of stem biomass the second, and root was the least. Under drought stress conditions, plants allocated more assimilates to the root, and root mass ratio and root-shoot ratio increased as the soil drought stress increasing, which enhanced the capacity ofE.hsienmuseedlings to resist drought stress. Comprehensive analysis showed that soil water content of 11.23%-14.44% was the threshold to the growth and biomass accumulation ofE. hsienmuseedlings be significantly inhibited by drought stress, which can be as the limit of soil water content forE. hsienmuseedling cultivation and afforestation.
Excentrodendron hsienmu; biomass; drought stress; phenotypic plasticity
10.14067/j.cnki.1673-923x.2017.05.004 http: //qks.csuft.edu.cn
2016-09-09
广西自然科学基金项目(2014GXNSFAA118132);广西林业科技项目(桂林科字[2014]02号)
庞世龙,本科,工程师
欧芷阳,博士,高级工程师;E-mail:ozhiyang@126.com
庞世龙,欧芷阳,申文辉,等.干旱胁迫对蚬木幼苗表型可塑性的影响[J].中南林业科技大学学报,2017, 37(5): 21-25.
S718.43;Q945. 3
A
1673-923X(2017)05-0021-05
[本文编校:谢荣秀]
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
河北果树(2021年4期)2021-12-02
天津医科大学学报(2021年1期)2021-01-26
今日农业(2020年19期)2020-12-14
医药前沿(2020年20期)2020-11-10
电脑报(2020年12期)2020-06-30
安徽农学通报(2020年7期)2020-05-26
中国野生植物资源(2019年2期)2019-06-11
河北农业科学(2019年6期)2019-03-21
农民致富之友(2017年15期)2017-08-20
