
海南岛油楠天然群体种子表型变异及其对种子萌发、幼苗生长的影响
2017-12-19 08:22吴忠锋杨锦昌尹光天李荣生邹文涛
中南林业科技大学学报 2017年4期
吴忠锋,杨锦昌,尹光天 ,李荣生 ,邹文涛,袁 洁
(1.广州市林业和园林科学研究院,广东 广州 510405;2.中国林业科学研究院热带林业研究所,广东 广州510520)
海南岛油楠天然群体种子表型变异及其对种子萌发、幼苗生长的影响
吴忠锋1,杨锦昌2,尹光天2,李荣生2,邹文涛2,袁 洁2
(1.广州市林业和园林科学研究院,广东 广州 510405;2.中国林业科学研究院热带林业研究所,广东 广州510520)
以海南岛油楠主要分布区6个天然群体为研究对象,对种子性状、种子萌发及幼苗生长指标进行系统的比较分析,采用巢式方差分析、变异系数及聚类分析等多种分析方法,对油楠种子表型变异进行了讨论,运用相关分析对种子性状间及其与地理因子、种子萌发、幼苗发育的相关性进行探讨。结果表明:油楠种子性状在种群间及种群内的差异均达到极显著水平;种子各性状平均表型分化系数为24.45%,即群体内变异远大于群体间变异;种子性状与地理因子间相关性较差;系统聚类可将油楠划分为3类,且与各地理分布区有高度吻合性;种子萌发及幼苗发育特征的种群间差异极显著,种子萌发率与种子性状不存在显著相关性,但种子千粒重与幼苗地径、苗高分别存在极显著、显著的相关性,说明种子千粒重越大,幼苗初期地径、苗高越大。本研究对油楠种质资源评价利用及苗木更新具有重要意义。
油楠;种子;表型变异;种子萌发;地理因子;幼苗
遗传多样性是物种多样性的基础,同时也是生物多样性的重要组成部分[1]。研究物种的遗传多样性,能够加深人们对生物多样性的起源和进化的了解,为物种分类、濒危动植物保护策略的制定提供理论依据[2-3]。遗传多样性的研究方法主要从表型、生化、蛋白质、分子水平等层面进行[4],其中,表型是物种各形态特征的组合,是生物遗传变异的表征,表型变异作为遗传变异的重要组成部分,已广泛应用于多样性研究。而种子表型变异较为稳定、不易受环境影响,在遗传多样性研究方面具有重要意义[5]。采用种子表型变异为研究对象,不仅可以揭示群体的遗传变异程度,同时根据种子变异与种子萌发率、幼苗出土、幼苗大小等方面的相关关系,进而了解种苗的定居和竞争能力及种群更新状况[6-10]。因此,研究种子表型变异在遗传多样性和种群更新方面的表现具有重要的生态学意义。
油楠Sindora glabra隶属于苏木科油楠属常绿阔叶乔木,为国家二级重点保护野生植物[11],天然分布于东南亚热带地区及我国海南,高20~30 m、胸径1 m以上,油楠属约10种,我国仅有一亚种,天然分布区海南白沙、昌江、东方、乐东、陵水、五指山等地[12-14]。油楠因其树干钻孔后流出油状树脂油,在能源、精油开发、医药等领域具有较高的发展潜力,引起了国内学者的关注[15-19]。然而,由于对油楠的遗传背景了解甚少,针对油楠的研究主要停留在资源调查、苗期实验以及树脂油成分分析,给油楠种质资源的评价利用和育种工作带来很大阻碍。本研究通过对我国油楠天然分布区的油楠种子性状的调查及苗期发芽试验,以期揭示表型变异的地理格局以及种子变异对种子萌发和幼苗特征影响,为今后油楠种质资源的评价利用、种子萌发及苗期研究提供理论依据。
1 材料与方法
1.1 材料收集
依据油楠在海南岛的大致分布范围,于2007年8月在前期实地勘查的基础上,选取了白沙、乐东、昌江、陵水、东方、五指山6个居群进行采种。鉴于油楠属国家二级保护植物,多零星分布于自然保护区内,居群密度偏小,加之受结实习性的影响,因此部分居群采种单株数量偏少。各采样居群的地理及气候因子见表1。

表1 海南岛油楠采样居群的地理因子及气候因子†Table 1 Geographical and ecological factors of natural populations of S.glabra for seed collection
1.2 种子大小和萌发及幼苗观测
种子采集和处理后测定其长度、宽度、厚度以及千粒重;采用千分尺测量种子长度、宽度和厚度,每个重复30粒,4次重复;千粒重采用百分粒法测定,8次重复。
测定不同单株种子大小后,从中随机抽取300粒种子,经80 ℃热水浸泡处理后均匀播于经消毒处理的沙床中[14];每个单株播种设3个重复,每重复100粒种子。播种期间进行常规浇水;播种后40 d观测不同单株种子的发芽数。幼苗移栽及管理措施如文献所述[18];移栽后12个月后量测苗木苗高和地径,并统计叶片数。
1.3 数据处理与分析
在所测种子表型性状指标的基础上,定义种子长宽比为种子形状指数。方差分析参照李斌等[20]、李文英和顾万春[21]的巢式方差分析方法,线性模型为Yijk=u+Pi+Ti(j)+e(ij)k。其中,Yijk为第i个种群第j个个体的第k个观测值;u为总均值;Pi为第i个群体的效应值(固定);Tj(i)为第i个群体内的第j个个体的效应值(随机);eijk=试验误差,分析油楠群体的表型变异特征;利用相关分析计算种子表型性状间及其与地理因子、气候因子间的相关性。
群体间表型分化系数Vst=δ2t/s/(δ2t/s+δ2s),其中δ2t/s是群体间方差值,δ2s是群体内方差值[22]。采用常规统计方法,统计各形态性状的平均值和标准差,用变异系数(CV)表示表型性状的离散程度,CV=S/x。
其它统计运算按照规定方法利用Excel2003,SPSS18.0,SAS8.1相关程序进行。
2 结果与分析
2.1 油楠天然群体种子表型变异特征
油楠5个种子表型性状群体的差异分析及多重比较见表2,方差分析结果表明,油楠种子性状在群体内和群体间都存在极显著的差异,这表明油楠种子性状稳定性较差,在群体内和群体间均存在广泛的变异,具有极其丰富的多样性。

表2 油楠6个天然群体种子性状平均值及方差分析†Table 2 Mean value and variance analysis of seed traits for 6 populations of S.glabra
各性状平均值和多重比较表明,不同种群间种子性状存在显著差异,种子长、宽、厚、形状指数、千粒重最大的群体依次为CJ、CJ、WZS、DF、WZS,最小的群体依次为LS、DF、LD、LS、LD。
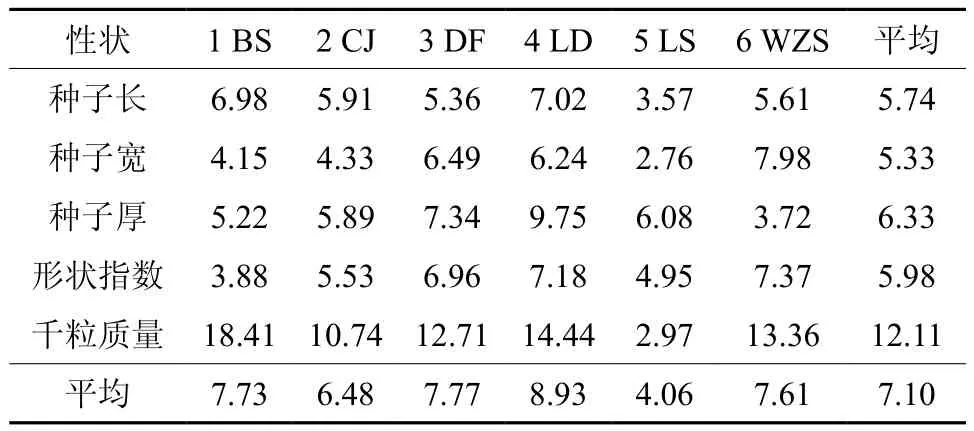
表3 油楠天然群体种子性状的变异系数(%)Table 3 Variation coefficients of phenotypic traits in S.glabra
由表3可知,油楠各性状的平均变异系数为7.1%,变异幅度为5.33%~12.11%。同一群体不同性状,如除LS群体外,千粒重的离散程度较其他性状有明显差异。同一性状在不同群体内的变异幅度也有较大差异,说明不同种群的环境异质性导致群体种子性状的差异。各群体种子性状的平均变异系数的排序为:LD(8.93)>DF(7.77)>BS(7.73)>WZS(7.61)>CJ(6.48)>LS(4.06),说明LD群体的表型多样性最为丰富,LS群体的表型多样性程度较低。
2.2 油楠种子变异来源及群体间的表型分化
按照非平衡巢式设计将各种子性状的变异分解为群体间变异、群体内变异和随机误差。群体间方差分量占总变异(群体间、群体内方差分量之和)的百分比即表型分化系数。由表4可以看出,油楠种子各性状遗传组成存在很大差异,群体间与群体内方差分量百分比分别为6.8%~37.65%和54.46%~91.61%,说明油楠天然群体种子性状的种群内变异远高于种群间变异。油楠种子性状群体间的表型分化系数在6.96%~39.71%之间,其中千粒重的表型分化系数最大(39.71%),而种子厚这一性状的表型分化系数最小(6.96%)。5个种子性状群体间的表型分化系数平均为24.45%,说明油楠的平均表型分化系数水平偏低。

表4 油楠群体间的方差分量及表型分化系数Table 4 Variance component and phenotypic differentiation coefficient among S.glabra populations
2.3 油楠种子性状间及其与地理因子相关性分析
表5显示了油楠种子性状间及其与地理、气候因子间的相关性。结果表明,种子千粒重与种子长、种子宽、形状指数存在显著相关,形状指数与种子长存在极显著负相关。种子性状与地理气候因子的相关分析表明,种子大小、种子千粒重与经纬度存在密切的正相关关系,与年均温存在密切的负相关关系,即越往东北方向,种子越大种子千粒重越大;温度越低,种子越小,千粒重越小。种子性状与海拔、降水的相关性不显著。
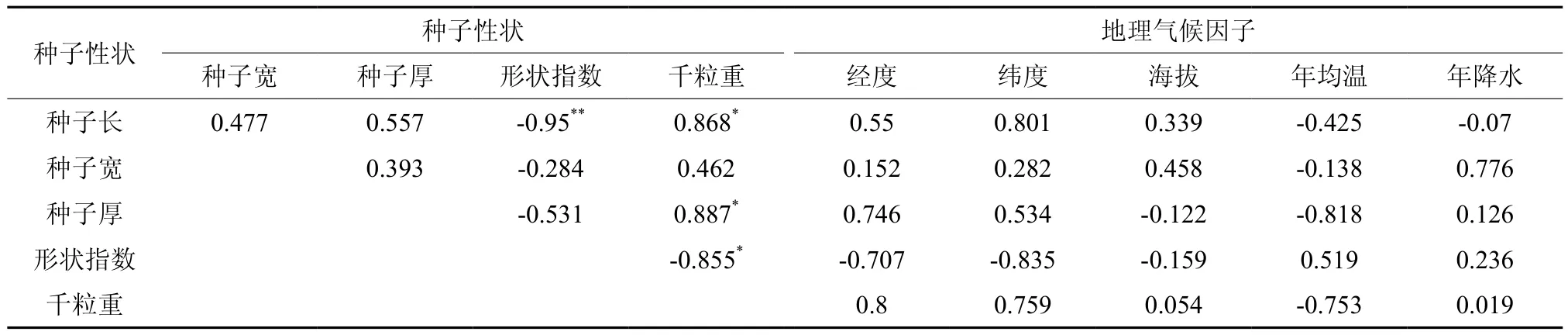
表5 油楠种子性状间及与地理因子间的相关性†Table 5 Correlations among phenotypic traits and correlation between phenotypic traits and geographic variables
2.4 不同地理种群油楠种子表型性状的聚类分析
利用欧式平均距离系统聚类方法,针对油楠6个天然群体的种子性状的表型数据进行聚类分析(图1),可将油楠6个天然种群分为3类:昌江(CJ)、WZS(五指山)、(白沙)BS划为一类;东方(DF)、乐东(LD)划为一类;陵水(LS)单独划为一类。考虑到油楠分布区的地理位置,昌江、五指山、白沙位于分布区东北部,东方、乐东位于分布区西南部,陵水位于分布区东部,说明分布区种群划分与地理位置具有高度的吻合型。
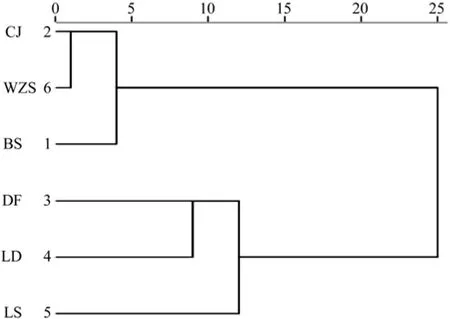
图1 不同地理种群油楠种子性状聚类Fig.1 UPGMA cluster based on the phenotypic traits of six populations in S.glabra
2.5 种子变异对萌发率的影响
由表6可知,各种群间发芽率存在极显著差异,其中发芽率最高种群为东方(69.26%),最低为陵水(31.11%)。但种子性状和地理因子与发芽率的相关分析结果显示,种子发芽率与种子性状不存在显著相关性,而与年降水量存在显著负相关,主要原因可能是种子萌发行为主要影响因素为物种特性、种子质量、播前处理状况和环境调控等,而与种子本身性状相关性不大。
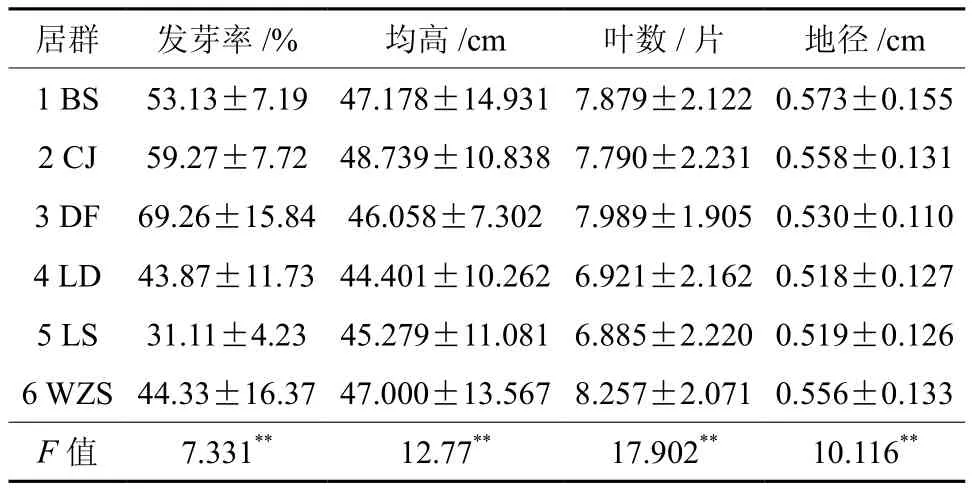
表6 油楠各群体发芽率及幼苗特征Table 6 The seed germination percentages and seedling characters of geographic populations
2.6 种子变异对幼苗发育的影响
对油楠幼苗初期生长特性的统计分析结果见表6,各种群间幼苗特征差异达极显著水平。幼苗生长状况(苗高、地径)较好的两个种群为白沙和昌江。由表7、表8可以看出,种子性状对幼苗生长初期具有很大影响。其中,种子长与苗高,形状指数与幼苗地径、苗高都存在显著相关性,地径与经度、苗高与纬度均存在极显著正相关,可见种子本身性状特征及地理因子都与幼苗生长存在密切的关系。种子千粒重与幼苗地径、苗高分别存在极显著、显著的相关性,说明种子千粒重越大,幼苗初期地径、苗高越大。
以种子千粒重、幼苗地径、苗高为研究对象,进行回归分析。种子千粒重与幼苗地径呈极显著正相关(R=0.925,P<0.01),回归方程为M(千粒重)=-3.892+11.0D(幼苗地径)(可信度为95%);种子千粒重与幼苗苗高呈显著正相关(R=0.907,P<0.05),回归方程为M(千粒重)=-2.006+0.087H(幼苗苗高)(可信度为95%)。

表7 幼苗发育特性与种子性状的相关性分析Table 7 Correlations coefficient between phenotypic traits and seedling characters

表8 幼苗发育特性与地理因子的相关性分析Table 8 Correlations coefficient between phenotypic traits and geographic variables
3 结论与讨论
种子大小和幼苗发育是植物生活史的重要组成部分,它们影响子代的更新,进而影响植物种群扩大及延续[23-24]。通过对油楠5个种子性状进行统计分析,各指标在种群间、种群内都存在极显著的差异,各指标种群内变异均大于种群间变异,符合一般自然种群的变异规律[25]。种群间、种群内多层次的变异为种质资源评价利用和生物多样性的保护提供了丰富的基础,然而这些变异主要来源于自身遗传因素或是环境因素还不得而知,通过分子标记技术了解油楠遗传多样性显得势在必行,以期能在遗传和环境两个方面了解油楠的变异情况,从而能更好的为种质资源的评价利用及多样性保护提供更全面的理论依据。
罗建勋和顾万春[26]认为,用变异系数表示性状值离散型特征,变异系数越大,则性状值离散程度越大。研究发现,油楠种子表型分化系数和变异系数水平偏低(24.45%,7.1%),说明油楠的种子形态的遗传特征较为稳定,种子作为生殖器官,极大程度上保证了油楠的物种稳定性[21];造成此结果的原因,也可能与油楠在我国的分布现状有关,到目前为止,油楠天然分布的报道只有我国海南,因此,所选种群的地理距离较小,造成种群间基因交流水平较高,降低了油楠种群间分化的可能性;环境因素作为影响表型变异的主要因素之一,分布区的环境因素越类似,则种群的遗传变异越小,从而导致了油楠表型分化系数偏低。与其他研究结果相比,高于皂荚Gleditsia sinensis的20.42%[27]、青钱柳Cyclocarya paliurus的20.54%[4],低于林芝云杉Picea linzhinesis的29.55%[28]、花椒树Sorbus pohuashanensis的45.76%[29]、山苍子Litsea cubeba的 60.19%[30]。后期研究中需考虑国外油楠的表型性状,扩大样本量,从而能更真实的反应油楠表型变异情况。
研究结果显示,油楠种子大小变异与种子萌发率没有显著相关性,但对幼苗初期生长特征有显著相关性,种子千粒重越大,幼苗初期地径与苗高越大。种子大小显著影响种子萌发只有少量报道[31],大量研究表明种子大小对发芽率没有显著影响[32-34],却对幼苗初期生长有显著相关性,Elwell等[35]、Hosseini等[36]研究结果表明,种子大小与幼苗初期生长特征(苗高、根系生长)存在显著正相关,这是由于大种子较小种子拥有更多的营养物质积累,为幼苗初期生长提供充足的营养,为幼苗后期的生长奠定条件,随着时间推移,优势逐渐减弱[37]。由于本苗期试验是在苗圃内完成的,有较好的水肥条件、光照条件,Gross等[6]研究表明较大的种子具有更好的萌发和生长能力,但在天然林中,林分郁闭度较大,幼苗初期生长较好,由于受光照不足等的影响,这种优势对幼苗后期的生长影响并不显著[38]。
对于种子大小与幼苗初期生长的相关性尚无定论,这需要开展从种子生长到苗木更新全过程的系统研究,由于种子大小影响植被幼苗更新的同时,也受到生长环境和时间的影响,因此环境异质性的结合研究更利于深入了解种子大小对植被更新的贡献[39],这对于种群生态学研究具有重要意义。目前,油楠天然分布在我国海南岛,群落水平及区系水平上群落更新及进化发育等方面的研究还没有报道,由于分布范围相对狭小,地理气候因子差异不大,遗传变异分析的样本量偏少,今后研究中可考虑国外油楠或者油楠属其他种作为研究对象,增大样本的丰富度,这将为了解油楠生态学特性及其他相关方面的研究提供理论基础。
[1]沈 浩, 刘登义.遗传多样性概述[J].生物学杂志, 2001,18(3): 5-7.
[2]张建英, 毛向红, 张莹莹, 等.利用RAMP分子标记对酸枣资源的遗传多样性分析[J].经济林研究, 2014, 32(2):14-18.
[3]Pigliucci M, Murren C J, Schlichting C D.Phenotypic plasticity and evolution by genetic assimilation[J].Journal of Experimental Biology, 2006, 209(12): 2362-2367.
[4]佘诚棋, 杨万霞, 方升佐, 等.青钱柳天然群体种子性状表型多样性[J].应用生态学报, 2009, 20(10): 2351-2356.
[5]孙 鹏, 李加茹, 段 伟, 等.河南省柿种质资源果实表型特征分析[J].经济林研究, 2015, 33(4): 9-17.
[6]Gross K L.Effects of seed size and growth form on seedling establishment of six monocarpic perennial plants[J].The Journal of Ecology, 1984, 72(2): 369-387.
[7]Seiwa K.Effects of seed size and emergence time on tree seedling establishment: importance of developmental constraints[J].Oecologia, 2000, 123(2): 208-215.
[8]Walters M B, Reich P B.Seed size, nitrogen supply, and growth rate affect tree seedling survival in deep shade[J].Ecology, 2000,81(7): 1887-1901.
[9]Fenner M, Thompson K.The ecology of seeds[M].Cambridge University Press, 2005.
[10]肖祥希, 高 楠, 何文广, 等.油杉表型差异分析及优树选择[J].中南林业科技大学学报, 2015, 35(7): 1-6.
[11]国家林业局, 农业部.国家重点保护野生植物名录(第一批).1999.
[12]中国科学院中国植物志编辑委员会.中国植物志(第39卷),豆科(一) [M].北京:科学出版社, 1988: 214-216.
[13]吴忠锋, 杨锦昌, 成铁龙, 等.海南油楠的重要生物学特性及产油特征[J].林业科学, 2014(4): 144-151.
[14]杨锦昌, 尹光天, 李荣生, 等.油楠实生苗培育试验研究[J].林业实用技术, 2011a(1): 29-30.
[15]杨锦昌, 尹光天, 吴忠锋, 等.香脂树泌油特性与机制及其应用研究进展[J].世界林业研究, 2015, 28(4): 32-38.
[16]李荣生, 尹光天, 杨锦昌, 等.油楠作为能源植物开发利用的思考[J].生物质化学工程, 2006, 40(B12): 161-164.
[17]李意德, 黄 全, 周铁锋, 等.海南岛能源植物资源及其利用潜力[J].生物质化学工程, 2006, 40(B12):240-246.
[18]杨锦昌, 尹光天, 吴仲民, 等.海南尖峰岭油楠树脂油的主要理化特性[J].林业科学, 2011b, 47(9): 21-27.
[19]张安菊, 梁振益, 陈祎平,等.油楠树脂油化学成分的研究[J].化学分析计量, 2013(3): 33-35.
[20]李 斌, 顾万春, 卢宝明.白皮松天然群体种实性状表型多样性研究[J].生物多样性, 2002, 10(2): 181-188.
[21]李文英, 顾万春.蒙古栎天然群体表型多样性研究[J].林业科学, 2005, 41(1): 49-56.
[22]葛 颂, 王明庥, 陈岳武.用同工酶研究马尾松群体的遗传结构[J].林业科学, 1988, 24(4): 399-409.
[23]Coomes D A, Rees M, Grubb P J, et al.Are differences in seed mass among species important in structuring plant communities?Evidence from analyses of spatial and temporal variation in dune-annual populations[J].Oikos, 2002, 96(3): 421-432.
[24]张 荣, 陈亚明, 孙国钧, 等.繁殖体与微生境在退化草地恢复中的作用[J].生态学报, 2004, 24(5):1000-0933.
[25]Thompson J N.Variation among individual seed masses in Lomatium grayi(Umbelliferae)under controlled conditions:magnitude and partitioning of the variance[J].Ecology, 1984,65(2): 626-631.
[26]罗建勋, 顾万春.云杉天然群体表型多样性研究[J].林业科学, 2005, 41(2): 66-73.
[27]李 伟, 林富荣, 郑勇奇, 等.皂荚南方天然群体种实表型多样性[J].植物生态学报, 2013, 37(1): 61-69.
[28]贾子瑞, 张守攻, 王军辉.林芝云杉天然群体针叶与种实的变异及其地理趋势[J].林业科学研究, 2011, 24(4): 428-436.
[29]郑 健, 郑勇奇, 宗亦尘, 等.花楸树天然群体种实多样性研究[J].植物遗传资源学报, 2009, 10(3): 385-391.
[30]田胜平, 汪阳东, 陈益存, 等.山苍子天然种群叶片和种实性状的表型多样性[J].生态学杂志, 2012, 31(7): 1665-1672.
[31]王桔红, 崔现亮, 陈学林, 等.中旱生植物萌发特性及其与种子大小关系的比较[J].植物生态学报, 2007,31(6):1037-1045.
[32]宗文杰, 刘 坤, 卜海燕, 等.高寒草甸51种菊科植物种子大小变异及其对种子萌发的影响研究[J].兰州大学学报(自然科学版), 2006, 42(5): 52-55.
[33]朱雅娟, 董 鸣, 黄振英.沙埋和种子大小对固沙禾草沙鞭的种子萌发与幼苗出土的影响[J].植物生态学报, 2005, 29(5):730-739.
[34]柯文山, 钟章成, 席红安, 等.四川大头茶地理种群种子大小变异及对萌发, 幼苗特征的影响[J].生态学报, 2000, 20(4):697-701.
[35]Elwell A L, Gronwall D S, Miller N D, et al.Separating parental environment from seed size effects on next generation growth and development in Arabidopsis[J].Plant cell & environment,2011, 34(2): 291-301.
[36]Hosseini N M, Palta J A, Berger J D, et al.Sowing soil water content effects on chickpea (Cicer arietinum L.): Seedling emergence and early growth interaction with genotype and seed size[J].Agricultural Water Management, 2009,96(12):1732-1736.
[37]李荣平, 蒋德明, 刘志民, 等.沙埋对六种沙生植物种子萌发和幼苗出土的影响[J].应用生态学报,2004,15(10):1865-1868.
[38]Westoby M, Jurado E, Leishman M.Comparative evolutionary ecology of seed size[J].Trends in Ecology & Evolution, 1992,7(11): 368-372.
[39]Moles A T, Westoby M.Seed addition experiments are more likely to increase recruitment in larger-seeded species[J].Oikos,2002, 99(2): 241-248.
The phenotypic variation of seed of Sindora glabra geographic populations and its effect on seed germination and seedling
WU Zhongfeng1, YANG Jinchang2, YIN Guangtian2, LI Rongsheng2, ZOU Wentao2, YUAN Jie2
(1.Guangzhou Institute of Forestry and Landscape Architecture, Guangzhou 510405, Guangdong, China;2.Research Institute of Tropical Forestry, Chinese Academy of Forestry, Guangzhou 510520, Guangdong, China)
Totally 6 natural population of Sindora glabra as test materials, fi ve characteristics of seeds, seed germination and seedling characteristics were investigated, using correlation analysis to study their relationship among them, the phenotypic diversity among and within the populations were studied by the method of nested variance analysis, clustering analysis, etc.Signi fi cant differences were observed in the 1 000-seed weight, seed length, seed width, seed thickness, seed length/width ratio among and within populations.The mean phenotypic differentiation coef fi cient among populations was 24.45%, and the variation within the populations was higher than that among the populations.The phenotypic traits had poor correlation within geographic factors.The 6 S.glabra populations could be clustered into 3 groups, and there were highly consistent within the geographic distribution.Signi fi cant differences were observed in the seed germination and seedling among the populations.Seed germination had no signi fi cant correlation with phenotypic traits, but 1 000-seed weight had significant correlation with stem base and height of seedling, all these phenomena demonstrated that the larger 1 000-seed weight was, the faster of seedling growth.These fi ndings offer basic data for the conservation biology and renewing of natural vegetation of these species.
Sindora glabra; seed;phenotypic variation; seed germination; geographic factors; seedling
S718.54;S722.1+3
A
1673-923X(2017)04-0064-06
10.14067/j.cnki.1673-923x.2017.04.011
2015-11-24
国家自然科学基金项目(31570656);林业公益性行业科研专项(201304604)
吴忠锋,硕士研究生 通讯作者:杨锦昌,副研究员,博士;E-mail:fjyjc@126.com
吴忠锋,杨锦昌,尹光天,等.海南岛油楠天然群体种子表型变异及其对种子萌发、幼苗生长的影响[J].中南林业科技大学学报,2017, 37(4): 64-69.
[本文编校:文凤鸣]
猜你喜欢
今日农业(2022年15期)2022-09-20
河北果树(2021年4期)2021-12-02
湖南电力(2021年1期)2021-04-13
天津医科大学学报(2021年1期)2021-01-26
医药前沿(2020年20期)2020-11-10
趣味(数学)(2020年4期)2020-07-27
支部建设(2020年15期)2020-07-08
河北农业科学(2019年6期)2019-03-21
中学生物学(2018年8期)2018-03-01
百科知识(2015年18期)2015-09-10
