
水稻叶鞘原生质体转化体系的构建及Pik-H4和AvrPik-H4蛋白的瞬时表达
2017-12-18 03:47刘维刘浩董双玉古丰玮陈志强王加峰王慧
中国农业科学 2017年23期
刘维,刘浩,董双玉,古丰玮,陈志强,王加峰,王慧
水稻叶鞘原生质体转化体系的构建及Pik-H4和AvrPik-H4蛋白的瞬时表达
刘维,刘浩,董双玉,古丰玮,陈志强,王加峰,王慧
(华南农业大学国家植物航天育种工程技术研究中心,广州 510642)
探索水稻叶鞘原生质体最佳游离时间以及转化时间,提高瞬时表达效率,在蛋白水平对目的基因进行检测且大批量表达。探索该系统表达抗稻瘟病蛋白Pik1-H4、Pik2-H4及无毒蛋白AvrPik-H4的可行性,对目的基因的功能进行分析。以高抗稻瘟病品种H4及对照品种中二软占作为试验材料,利用1/2 MS培养基种植水稻幼苗25℃恒温培养7—10 d。利用纤维素酶及离析酶对水稻叶鞘进行酶解,血球计数板分别统计游离1、2、3、4、5、6、7、8、9、10、11和12 h的细胞数目获得最佳的酶解时间。将目标基因Pik-H4、Pik-H4及分别与GFP融合,构建瞬时表达载体,利用PEG介导转入水稻叶鞘原生质体。设置转化10、12、14、16、18、20、22和24 h,分别提取细胞总RNA,管家基因为对照,设计特异性扩增引物,利用实时荧光定量PCR(qRT-PCR)检测的相对表达量,探索最佳的转化时间。通过激光共聚焦扫描显微镜对目标基因进行亚细胞定位观察,推测基因功能。提取细胞总蛋白,以Anti-GFP为一抗用Western blot对目标蛋白表达进行验证。相对室温土壤栽培,营养丰富且均衡的1/2 MS培养基恒温种植的水稻幼苗质量更优质,活力更高。游离时间的长短对游离效率影响较大,游离的最佳时间为4—6 h,在3—4 h细胞游离数目增长速度最快,4—6 h细胞数量趋于平稳,6 h以后细胞总量呈现下降趋势,特别是7 h以后,显微下细胞碎片增多,细胞死亡速度加快。检测的相对表达量,获得最佳转化时间为14—16 h,16 h达到最高值,之后逐渐下降。随着时间的推移,荧光显微镜下观察到GFP蛋白发出的荧光逐渐淬灭。亚细胞定位观察发现AvrPik-H4蛋白主要被定位于水稻细胞膜上,初步推测是一种膜蛋白,通过某种形式运输到宿主细胞作为激发子触发一系列反应。是高效广谱抗稻瘟病基因,分为Pik-H4和Pik-H4两个部分,Pik1-H4主要定位在内质网,Pik2-H4主要定位在质体,从定位结果初步推定Pik1-H4可能主要参与AvrPik-H4蛋白的识别反应,Pik2-H4主要起到调控下游抗病的作用。Western blot结果显示目标蛋白表达成功,分子大小正确。Pik1-H4和AvrPik-H4的表达量高于Pik2-H4,说明分子量的大小不是影响转化效率的关键因素。水稻叶鞘原生质体瞬时表达系统具有高效快速的特点,通过对游离及转化时间的探索为水稻瞬时表达体系的广泛实践应用提供参考。目标基因的成功表达为Pik-H4与无毒蛋白互作机制的研究提供了有价值的理论依据。
水稻;原生质体;亚细胞定位;western 杂交
0 引言
【研究意义】稻瘟病是由稻瘟病菌()引起的全球性真菌性病害,目前世界上有85个国家已出现该病害,每年减产10%—35%[1]。实践证明,选育和使用水稻抗病品种是控制稻瘟病最为经济有效的措施。植物瞬时表达系统以宿主单个细胞为基础,转入外源的DNA在短时间内进行蛋白质高水平表达。由于水稻原生质体的生物功能在一定程度上与完整的细胞相似,为研究植物细胞内信号转导提供了非常有利的细胞环境[2]。利用水稻叶鞘原生质体瞬时表达系统表达Pik1-H4、Pik2-H4及AvrPik-H4蛋白并进行亚细胞定位,有助于目的基因分子功能及抗病基因和无毒基因互作机制的研究。【前人研究进展】水稻抗稻瘟病蛋白基因由和组成,属于典型的NBS-LRR类基因,对广东的稻瘟病菌生理小种大多表现出高抗[3]。为编码113 aa的小分子分泌蛋白,不含有保守区域,容易发生突变。相较稳定的转基因表达系统,瞬时表达体系具有宿主范围广、周期短、检测快速及高通量的特点,已被广泛应用于分子生物学研究领域,如启动子活性分析[4]、Cas9编辑效率检测[5-6]、mRNA衰变[7]、microRNAs对靶基因的调控[8]、蛋白功能研究[9]、信号转导[10-11]等。目前主要通过基因枪法、农杆菌渗透法、聚乙二醇(PEG)介导法、电击法及植物病毒载体介导对目标基因进行转化[12],在小麦[5]、葡萄[9]、玉米[10]、水稻[13]、拟南芥[14]、樱桃[15]、莴苣[16]、马铃薯[6,17]、烟草[18]等都有应用。水稻叶表面具有蜡质层,不利于原生质体的游离,如鹿连明等[19]利用烟草的瞬时表达体系研究水稻条纹病毒(,RSV)的相关蛋白互作,但基因产物的正确折叠、亚细胞精细定位都依赖于宿主特异性表达系统[20]。近几年,有关水稻原生质体瞬时表达系统已经有相关报道,转化效率也相对提高[21],如利用Co-IP技术验证互作蛋白[13],探索ABA信号通路[22],邻近生物素(BioID)技术筛选近端蛋白[23]等。【本研究切入点】尽管对水稻原生质体瞬时表达系统的应用已经有少量报道,但原生质体游离转化效率限制了该技术的应用,利用该技术进行抗稻瘟病基因蛋白的表达还未见报道。【拟解决的关键问题】探索原生质体游离及转化的最佳时间,通过将目的片段融合绿色荧光蛋白(GFP)构建载体,利用水稻叶鞘原生质体瞬时表达体系成功表达目标蛋白,在激光共聚焦扫描显微镜下观察蛋白在细胞内的表达部位,并且利用Western blot技术验证结论的真实性,为水稻原生质体瞬时表达系统的推广应用提供依据,为水稻抗病相关基因的功能性研究及互作蛋白的筛选打下基础。
1 材料与方法
试验于2016年10月至2017年3月在华南农业大学国家植物航天育种技术工程研究中心完成。
1.1 试验材料
水稻材料为H4以及中二软占,H4是经过空间搭载诱变的中二软占突变体经地面选育的广谱、高抗的水稻优质种质资源。
1.2 载体构建
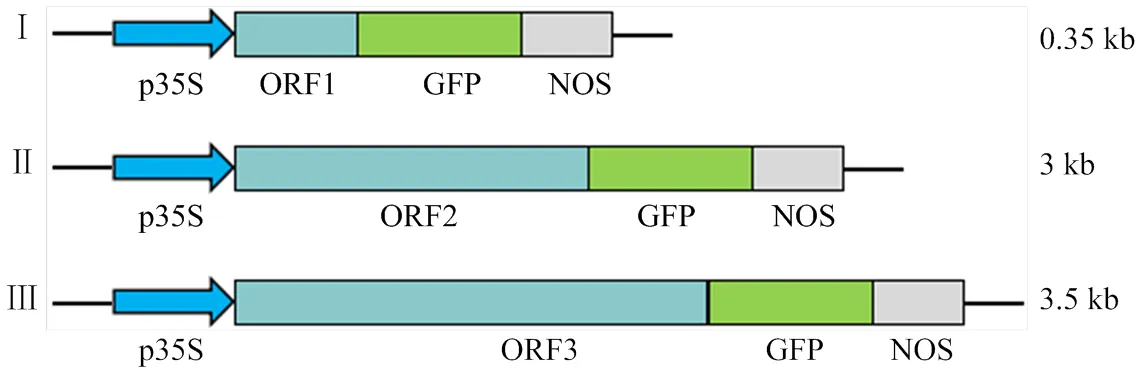
试验中所有的瞬时表达载体均以pYL322d1- eGFPn为骨架,如图1所示,由花椰菜花叶病毒(CaMV)35S启动子、绿色荧光蛋白(GFP)片段和NOS终止子组成。目标基因Pik-H4、Pik-H4克隆于高抗稻瘟病水稻品种H4的cDNA,克隆于稻瘟病菌GD0193,利用CE Design V1.03软件设计引物,如表1所示,去除开放阅读框架片段终止密码子,利用同源重组方法构建p35S-Pik1-H4/Pik2-H4/ AvrPik- H4-GFP载体。载体骨架由亚热带农业生物资源保护与利用国家重点实验室刘耀光研究员惠赠。

表1 引物寡核苷酸序列
1.3 水稻幼苗种植
挑选饱满的H4水稻种子,去壳,75%乙醇消毒2 min,2%的次氯酸钠150 r/min消毒30 min。在超净工作台中用灭菌水洗净残留的次氯酸钠后,将种子接种到1/2 MS培养基中,放到恒温培养箱,25℃,光周期为12 h光照/12 h黑暗培养10 d左右。
1.4 水稻原生质体制备
参照YANG等[13]的原生质体游离方法,沿根部将水稻幼苗剪下,用锋利的刀片将水稻叶鞘切割成0.5—1 mm的片段,放到0.6 mol·L-1甘露醇中,室温预质壁分离30 min,吸干残留的甘露醇,用W5(154 mmol·L-1NaCl,125 mmol·L-1CaCl2,5 mmol·L-1KCl,2 mmol·L-1MES,pH 5.7)溶液稍清洗后加入现配酶液(0.5 mol·L-1甘露醇,10 mmol·L-1MES,1.5%纤维素酶,0.75%离析酶,10 mmol·L-1CaCl2,0.1% BSA),30℃,60 r/min,酶解4—6 h。在普通光学显微镜40×物镜下观察细胞,视野中有20—40个完整的细胞即可。用200目筛子过滤酶液,加入10 ml预冷的W5溶液于酶解的残渣快速手摇1 min,再次过滤,合并滤液,300×,离心5 min,吸出上清液。向BD管中加入2 ml预冷的W5溶液,让原生质体重新悬浮,200×,离心3 min,重复此步骤一次。用500 μl W5重悬沉淀,置于冰上30 min后,150×,离心2 min,去除上清,用1 ml MMg悬浮细胞(使终浓度约为107个/ml)。

图1 322-d1-eGFPn载体结构图
1.5 PEG介导水稻原生质体转化
在100 μl原生质体中,加入高纯度10 μg(约10 μl)质粒,再加入110 μl 40% PEG(40% PEG4000,0.3 mol·L-1甘露醇,0.1 mol·L-1CaCl2),混匀后,28℃黑暗中横放15 min。加入500 μl的W5溶液终止反应,充分混匀,300×离心5 min,去除上清,加入600 μl WI溶液(4 mmol·L-1MES,pH 5.7,0.5 mol·L-1甘露醇,20 mmol·L-1KCl),28℃黑暗中培养14—16 h。
1.6 亚细胞定位
使用激光扫描共聚焦显微镜(LSM 7 DUO)观察转化后含有GFP蛋白以及GFP融合蛋白的原生质体,GFP、mCherry的激发波长分别为488、561 nm,发射波长分别为530—560、580—620 nm。叶绿体自发荧光的激发波长为488 nm,发射波长为650—750 nm。
1.7 Western验证
收集转化后的原生质体,300×,离心6 min,尽量去除上清。加入20 μl的SDS-PAGE样品缓冲液(50 mmol·L-1Tris-HCl,2% SDS,0.1%溴酚蓝,10%甘油,1%巯基乙醇)100℃煮沸5 min,提取原生质体中的总蛋白。室温离心1 min,吸取上清,用10%的SDS-聚丙烯凝胶(Bio-Rad)200 V 35 min分离样品的蛋白,300 mA 3 h将蛋白转到硝酸纤维素膜上,使用一抗Anti-GFP温室孵育1 h,二抗Anti-Mouse温室孵育1 h,使用化学发光试剂(Thermo Scientific)温室孵育5 min后,进行观察。
1.8 实时荧光定量PCR分析
为确定转化的最佳时间,在原生质体中转入GFP载体,分别提取转化10、12、14、16、18、20、22、24 h的产物,300×离心5 min,弃上清,加入500 μl TRIzol,涡旋15 s,温室静置3 min。加入150 μl氯仿,涡旋15 s,静置2 min,4℃,12 000×离心5 min,吸取水相到新的离心管,加入250 μl预冷的异丙醇,混匀后温室放置3 min,4℃,12 000×离心5 min,弃上清,加入1 ml 75%乙醇,简单混匀后4℃,12 000×离心2 min,去除残留酒精,加入20 μl RNase水溶解沉淀。分别取1 μg RNA用SMARTScribeTMReverse Transcriptase试剂盒(TakaRa Clontech)进行逆转录,反应条件:30℃ 10 min,42℃ 20 min,99℃ 5 min,4℃ 5 min,瞬间离心后,将样品浓度稀释至300—500 ng,以此为模板做qPCR定量分析,设计PCR引物GFP-F:GACGACGGCAACTACAAGAC、GFP-R:TCGGCCATGATATA GACGTT,产物为163 bp,管家基因作为对照。使用AceQ qPCR SYBR@Green MasterMix试剂盒(Vazyme),反应条件:95℃ 5 min,95℃ 10 s,60℃ 30 s,40个循环。
2 结果
2.1 瞬时表达载体的鉴定
将目的片段分别与GFP进行组装,如图2所示,选用RⅠ、Ⅰ两个限制性内切酶将载体线性化,选用这两个酶切位点插入目的片段不会导致移码,保证目标蛋白的正确表达。经PCR检测后结果如图3所示,除了Pik1-H4-GFP有一个扩增片段不正确以外,其余都正确,挑取PCR结果正确质粒送去公司测序,结果表明组装成功。
2.2 水稻原生质体游离
选取茁壮、叶鞘较硬的幼苗,用手术刀片将叶鞘切割成小段(图4)。通过血球计数板统计获得不同游离时间细胞的数目,游离的最佳时间为4—6 h,在3—4 h细胞游离数目增长速度最快,4—6 h细胞数量趋于平稳,6 h以后细胞总量呈现下降趋势,特别是7 h以后,显微下细胞碎片增多,细胞死亡速度加快(图5)。随着游离时间的增加,细胞总数目增多,导致供氧不足,初始的游离细胞活力下降,加上游离过程中的机械碰撞使细胞膜破碎,从而导致总体细胞数目呈现下降趋势。
2.3 实时荧光定量PCR分析最佳转化时间
GFP蛋白在蓝色波长范围的光线激发下,发出绿色萤光。将GFP载体转化到原生质体中,28℃,经过14—16 h的暗培养后,吸取5 μl在荧光显微镜下进行检测,可以看到GFP蛋白发出的荧光(图6)。从图中可以看出,GFP载体有较高的转化效率,经过暗培养后,大部分细胞还保持有较完整的形态,较高的生命活力。图中也存在一部分细胞碎片,说明存在一部分的细胞死亡,有一定比例的损失。
Ⅰ: AvrPik; Ⅱ: Pik1-H4; Ⅲ: Pik2-H4

M: 1 kb Marker; 1-6: Pik1-H4-GFP; 7-12: Pik2-H4-GFP; 13-18: AvrPik-GFP

a:水稻叶鞘游离过程图Process of rice leaf sheath protoplasts;b:水稻叶鞘原生质体图Observation of protoplasts isolation from leaf sheath
为探索原生质体转化的最佳时间,通过转化GFP质粒,设置不同的转化时间,通过提取原生质体的总RNA,用实时荧光定量技术估测的相对表达量,获得最佳转化时间为14—16 h相对表达量最高的转化时间为16 h,之后逐渐下降(图7)。
2.4 Pik1-H4、Pik2-H4及AvrPik-H4的亚细胞定位
大部分基因产物与特定的细胞器有一定的关联,可通过亚细胞定位来探索该蛋白的功能以及蛋白相互作用的网络。将瞬时表达载体转染到水稻的原生质体中,通过激光共聚焦扫描显微镜63×水镜观察到目标蛋白在水稻细胞内的具体表达部位。由图8可知AvrPik-H4主要定位于细胞膜,Pik1-H4主要定位于内质网,Pik2-H4主要定位于质体。AvrPik-H4蛋白被定位于水稻细胞膜上,说明是一种膜蛋白或者积累于膜上的蛋白,通过某种形式运输到宿主细胞作为激发子触发一系列反应。从定位结果初步推定Pik1-H4可能主要参与AvrPik蛋白的识别反应以及信号传递的作用,Pik2-H4主要起到改变能量的传输方式及调控下游抗病引发过敏性坏死的作用。相比烟叶、洋葱表皮细胞的瞬时转化体系,水稻叶鞘细胞更具有说服力,有正确的蛋白合成系统,能引导目标蛋白的正确折叠。

图5 酶解时间对原生质体产量的影响

a:GFP荧光GFP filter;b:白光bright field

图7 不同转化时间GFP相对表达量

图8 AvrPik-H4、Pik1-H4及Pik2-H4亚细胞定位
2.5 Western blot验证
收集转化结束后原生质体,提取总蛋白,进行Western blot验证,结果如图9所示。融合蛋白比目标蛋白分子量增加30 kD左右,用Anti-GFP作为一抗进行孵育,其中泳道1是没有转入质粒的细胞总蛋白作为阴性对照,泳道2转入GFP质粒作为阳性对照,泳道3—5是目的片段与GFP的融合蛋白,与Marker进行对照结果表明正确,为亚细胞定位的准确性提供了有力证据。

1:阴性对照Negative control;2:阳性对照Positive control; 3:Pik1-H4-GFP;4:Pik2-H4-GFP;5:AvrPik-H4-GFP
3 讨论
水稻作为单子叶模式植物被广泛地应用于分子功能、遗传进化研究,是探索基因组学和比较基因组学的有效工具[24]。利用水稻转基因植株进行基因功能研究存在周期长、生物安全性问题,而水稻瞬时表达体系能在短时间内实现目标基因的高水平表达,且保留原有的合成、修饰及转运蛋白的途径,更有益于后续接近真实情况抗病相关蛋白的筛选。目前,水稻系统的瞬时表达主要采用原生质体表达系统[25]、农杆菌侵染[26]方法,相对来说原生质体的方法更便捷高效。水稻叶片表面的蜡质层含有10%的硅胶[27],不利于被纤维素酶降解,而水稻幼苗叶鞘的硅胶含量较低[28],适合做原生质体游离的材料。本研究利用1/2 MS培养基25℃种植水稻幼苗,得到了较好的游离效果。相对于土壤栽培,培养基种植能提供更丰富均衡的营养,提供了更高质的游离材料。PEG与二价阳离子共价结合时能介导DNA发生有效沉淀达到转化的目的。有研究表明载体分子量越大转化效率越低,Bart等[29]研究表明12 kb质粒转化效率为25%—30%;YANG等[13]研究表明5.9 kb质粒转化效率达到70%左右,3 kb的GFP质粒转化效率达到90%以上;段炼等[30]用蔗糖密度梯度法纯化水稻原生质体,质粒转化浓度为0.7 μg∙μL-1时转化效率达到60%—70%。本研究中,4.7 kb的GFP载体转化效率可达95%以上,融合GFP的目的基因载体大小分别为5、7.7、8.2 kb,GFP空载体与AvrPik-H4-GFP的转化效率相对稍高,但并无太大差别。相对以前的报道,本研究采用离心的方法对细胞进行收集,相对蔗糖密度梯度纯化法简便高效,利用实时荧光定量PCR法得到最佳转化时间为14—16 h,获得了较高的转化效率。Western blot验证结果显示,Pik1-H4-GFP与AvrPik-H4-GFP的表达量最高,说明转化效率与质粒分子大小无显著的线性关系,蛋白的表达量则与蛋白本身的功能性质有关。PEG介导的原生质体转化对DNA属于无选择性吸收,要同时保证质粒的高浓度及高质量才能获得较高的转化效率。
位于水稻第11号染色体,由两个相邻的NBS-LRR基因Pik、Pik组成,NBS-LRR类是水稻抗稻瘟病基因中最常见的编码结构域[31]。位点存在7个等位基因(、、、、、、),对稻瘟病菌都具有广谱抗性[32-33]。利用水稻瞬时表达系统对Pik-H4及AvrPik-H4蛋白的成功表达以及亚细胞定位对蛋白的功能研究有非常重要的意义,可为揭示靶蛋白介导的抗性通路提供依据。本研究表明在H4幼苗接种GD0193稻瘟病菌24 h游离的原生质体中转入无毒蛋白AvrPik-H4后,在短时间内大部分水稻细胞聚集成团呈现胶稠透明状发生过敏性坏死,而对照品种中二软占则不会有此现象,说明介导的免疫反应是通过迅速而高效地引起宿主过敏性坏死实现的。该现象为目标基因在水稻叶鞘原生质体中的成功表达提供了有力证据。
瞬时表达系统常用于靶蛋白的亚细胞定位,如农杆菌侵染烟草[34]、洋葱[35]及拟南芥表皮瞬时表达[36]。介于异源表达系统蛋白修饰、转运存在差异可能出现的错定位,同源表达系统的亚细胞定位结果更接近真实情况[37]。Zhai等[38]利用水稻原生质体表达系统定位Pikh-1、Pikh-2及AvrPik-h均在细胞质和细胞核。靶基因的亚细胞定位对基因功能的研究有重要意义,本研究中将目标蛋白分别与GFP蛋白融合表达,利用水稻叶鞘原生质体瞬时表达系统进行亚细胞定位,观察到Pik1-H4主要定位于内质网,Pik2-H4主要定位于质体,AvrPik-H4主要定位于细胞膜。Pikh是Pik-H4的等位基因,二者定位结果出现差异的原因有两个:(1)CDS碱基序列存在差异。Pikh-1与Pik1-H4的CDS序列存在两个碱基的差异,Pikh-2与Pik2-H4的CDS序列则完全相同,AvrPikh与AvrPik-H4的CDS序列存在一个碱基的差异,可能改变其在细胞内的定位;(2)病原菌入侵时靶蛋白定位发生改变。本研究中的游离材料取自于接种稻瘟病菌24 h后的幼苗,有研究表明一些抗病相关基因会随着病原菌的入侵而改变细胞中的定位,由细胞核流向细胞质,如定位于叶绿体的NRIP1蛋白识别病原菌效应因子后,会从叶绿体流向细胞质及细胞核[39]。有研究表明与的CC结构域互作产生相应的信号转导后,使宿主产生相应的抗性[38]。初步推测Pik1-H4主要起到一个无毒蛋白与宿主抗病的衔接作用,识别外源物质入侵及启动防御信号传递给Pik2-H4,能够受到病原物的诱导而表达。Pik2-H4接收到信号以后通过调配代谢物的合成以及物质运输方式启动宿主的防御模式,加厚被侵染细胞的细胞壁或者直接启动自杀机制阻止相邻细胞被侵菌丝侵入。AvrPik在稻瘟病菌孢子的细胞膜上主要起到信号传递的作用,便于被宿主识别。
4 结论
水稻叶鞘原生质体游离的最佳时间为4—6 h,最适转化时间为14—16 h,FAD染色发现得到的细胞具有较高的活力,检测GFP荧光观察到较高的转化效率,通过亚细胞定位观察及Western blot验证目标蛋白的表达,为Pik-H4与无毒蛋白互作机制的研究打下了基础。推测Pik1-H4可能主要参与AvrPik-H4蛋白的识别反应,Pik2-H4主要调控下游抗病反应。
[1] FISHER M C, HENK D A, BRIGGS C J, BROWNSTEIN J S, MADOFF L C, MCCRAW S L, GURR S J. Emerging fungal threats to animal, plant and ecosystem health., 2012, 484(7393): 186-194.
[2] NANJAREDDY K, ARTHIKALA M, BLANCO L, ARELLANO E, LARA M. Protoplast isolation, transient transformation of leaf mesophyll protoplasts and improved-mediated leaf disc infiltration of: tools for rapid gene expression analysis., 2016, 16: 53.
[3] XIAO W, YANG Q, WANG H, GUO T, LIUY, ZHU X, CHEN Z. Identification and fine mapping of a resistance gene toin a space-induced rice mutant., 2011, 28(3): 303-312.
[4] 孙琴, 孙进华, 王树军, 李焕苓, 王家保. 荔枝16-2基因启动子的克隆与瞬时表达分析. 热带作物学报, 2016, 37(4): 736-741.
SUN Q, SUN J H, WANG S J, LI H L, WANG J B. Cloning and transient expression assay of16-2 gene promoter from litchi., 2016, 37(4): 736-741. (in Chinese)
[5] ZHANG Y, LIANG Z, ZONG Y, WANG Y, LIU J, CHEN K, QIU J, GAO C. Efficient and transgene-free genome editing in wheat through transient expression of CRISPR/Cas9 DNA or RNA., 2016, 7: 12617.
[6] ANDERSSON M, TURESSON H, NICOLIA A, FALT A, SAMUELSSON M, HOFVANDER P. Efficient targeted multiallelic mutagenesis in tetraploid potato () by transient CRISPR-Cas9 expression in protoplasts., 2016, 36(1): 117-128.
[7] HAYASHI S, WAKASA Y, OZAWA K, TAKAIWA F. Characterization of IRE1 ribonuclease-mediated mRNA decay in plants using transient expression analyses in rice protoplasts., 2016, 210(4): 1259-1268.
[8] BAI Y, HAN N, WU J, YANG Y, WANG J, ZHU M, BIAN H. A transient gene expression system using barley protoplasts to evaluate microRNAs for post-transcriptional regulation of their target genes., 2014, 119(1): 211-219.
[9] ZHAO F L, LI Y J, HU Y, GAO Y R, ZANG X W, DING Q, WANG Y J, WEN Y Q. A highly efficient grapevine mesophyll protoplast system for transient gene expression and the study of disease resistance proteins., 2016, 125(1): 43-57.
[10] CAO J, YAO D, LIN F, JIANG M. PEG-mediated transient gene expression and silencing system in maize mesophyll protoplasts: a valuable tool for signal transduction study in maize., 2014, 36(5): 1271-1281.
[11] IM J H, YOO S D. Transient expression inleaf mesophyll protoplast system for cell-based functional analysis of MAPK cascades signaling//Komis G, ŠAMAJ J.. Humana Press, 2014: 3-12.
[12] 赵文婷, 魏建和, 刘晓东, 高志晖. 植物瞬时表达技术的主要方法与应用进展. 生物技术通讯, 2013, 24(2): 294-300.
ZHAO W T, WEI J H, LIU X D, GAO Z H. Advance of the main methods and applications of plant transient expression system., 2013, 24(2): 294-300. (in Chinese)
[13] YANG J W, FU J X, Li J, CHENG X L, LI F, DONG J F, LIU Z L, ZHUANG C X. A novel co-immunoprecipitation protocol based on protoplast transient gene expression for studying protein-protein interactions in rice., 2014, 32(1): 153-161.
[14] PLANCHAIS S, CAMBORDE L, JUPIN I. Protocols for studying protein stability in anprotoplast transient expression system//LOIS L M, MATTHIESEN R., 2016, 1450: 175-194.
[15] YAO L, LIAO X, GAN Z, PENG X, WANG P, LI S, LI T. Protoplast isolation and development of a transient expression system for sweet cherry (L.)., 2016, 209: 14-21.
[16] CHEN Q, DENT M, HURTADO J, STAHNKE J, MCNULTY A, LEUZINGER K, LAI H. Transient protein expression by agroinfiltration in lettuce//MACDONALD J, KOLOTILIN I, MENASSA R.. Humana Press, 2016: 55-67.
[17] NICOLIA A, PROUX-WERA E, AHMAN I, ONKOKESUNGA N, ANDERSSON M, ANDEASSONB E, ZHU L H. Targeted gene mutation in tetraploid potato through transient TALEN expression in protoplasts., 2015, 204: 17-24.
[18] PITINO M, ARMSTRONG C M, CANO L M, DUAN Y. Transient expression ofLiberibacter Asiaticus effector induces cell death in., 2016, 7: Article 982.
[19] 鹿连明, 秦梅玲, 王萍, 兰汉红, 牛晓庆, 谢荔岩, 吴祖建, 谢联辉. 利用免疫共沉淀技术研究RSV、CP、SP和NSvc4蛋白的互作. 农业生物技术学报, 2008, 16(5): 891-897.
LU L M, QIN M L, WANG P, LAN H H, NIU X Q, XIE L Y, WU Z J, XIE L H. Studies on the interactions between RSV CP, SP and NSvc4 proteins using co-immunoprecipitation technology., 2008, 16(5): 891-897. (in Chinese)
[20] 王华忠, 陈雅平, 陈佩度. 植物瞬间表达系统与功能基因组学研究. 生物工程学报, 2007, 23(3): 367-374.
WANG H Z, CHEN Y P, CHEN P D. Plant transient expression system in functional genomics., 2007, 23(3): 367-374. (in Chinese)
[21] ZHANG Y, SU J, DUAN S, AO Y, DAI J, LIU J WANG P, LI Y, LIU B, FENG D, WANG J, WANG H. A highly efficient rice green tissue protoplast system for transient gene expression and studying light/chloroplast-related processes., 2011, 7: 30.
[22] KIM N, MOON S, MIN M K, CHOI E, KIM J, KOH E, YOON I, BYUN M, YOO S, KIM B. Functional characterization and reconstitution of ABA signaling components using transient gene expression in rice protoplasts., 2015, 6: Article 614.
[23] LIN Q P, ZHOU Z J, LUO W B, FANG M, LI M, LI H. Screening of proximal and interacting proteins in rice protoplasts by proximity-dependent biotinylation., 2017, 8: Article 749.
[24] CANTRELL R P, REEVES T G. The cereal of the world’s poor takes center stage., 2002, 296(5565): 53.
[25] 郭萍, 武瑶, 李嘉, 方荣祥, 贾燕涛. 利用水稻原生质体快速分析miRNA靶标RNA. 生物工程学报, 2014, 30(11): 1751-1762.
GUO P, WU Y, LI J, FANG R X, JIA Y T. Efficient transient expression to analyze miRNA targets in rice protoplasts., 2014, 30(11): 1751-1762. (in Chinese)
[26] ANDRIEU A, BREITLER J C, SIRE C, MEYNARD D, GANTET P, GUIDERDONI E. An in planta,-mediated transient gene expression method for inducing gene silencing in rice (L.) leaves., 2012, 5: 23.
[27] ISLAM M A, DU H, NING J, YE H, XIONG L. Characterization of-homologous genes in rice involved in leaf wax accumulation and drought resistance., 2009, 70(4): 443-456.
[28] MA J F, YAMAJI N. Silicon uptake and accumulation in higher plants., 2006, 11(8): 392-397.
[29] BART R, CHERN M, PARK C, BARTLEY L, RONALD P C. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts., 2006, 2: 13.
[30] 段炼, 钱君, 郭小雨, 朱英. 一种快速高效的水稻原生质体制备和转化方法的建立. 植物生理学报, 2014, 50(3): 351-357.
DUAN L, QIAN J, GUO X Y, ZHU Y. A rapid and efficient method for isolation and transformation of rice protoplast., 2014, 50(3): 351-357. (in Chinese)
[31] MARTIN G B, BOGDANOVE A J, SESSA G. Understanding the functions of plant disease resistance proteins., 2003, 54(1): 23-61.
[32] ASHIKAWA I, HAYASHI N, ABE F, WU J, MATSUMOTO T. Characterization of the rice blast resistance gene Pik cloned from Kanto51., 2012, 30(1): 485-494.
[33] CHEN J, PENG P, TIAN J, HE Y, ZHANG L, LIU Z, YIN D, ZHANG Z., a rice blast resistance allele consisting of two adjacent NBS-LRR genes, was identified as a novel allele at thelocus., 2015, 35(5): 117
[34] 霍琳, 及晓宇, 王玉成. 农杆菌介导的烟草瞬时表达试验条件优化. 分子植物育种, 2016, 14(1): 80-85.
HUO L, JI X Y, WANG Y C. Transient expression conditions of tobacco transformation mediated by., 2016, 14(1): 80-85. (in Chinese)
[35] 刘海燕, 冯冬茹, 刘兵, 何炎明, 王宏斌, 王金发. 农杆菌介导的MpASR蛋白在洋葱表皮细胞的定位研究. 热带亚热带植物学报, 2009, 17(3): 218-222.
LIU H Y, FENG D R, LIU B, HE Y M, WANG H B, WANG J F. Studies on subcellular localization of MpASR in onion epidermal cells mediated by., 2009, 17(3): 218-222. (in Chinese)
[36] ROSAS-DIAZ T, CANA-QUIJADA P, AMORRIM-SILVA V, BOTELLA M, LOZANO-DURAN R, BEJARANO E.plants as a suitable and efficient system for transient expression using., 2017, 10(2): 353-356.
[37] MARION J, BACH L, BELLEC Y, MEYER C, GISSOT L, FAURE J. Systematic analysis of protein subcellular localization and interaction using high-throughput transient transformation ofseedlings., 2008, 56(1): 169-179.
[38] ZHAI C, ZHANG Y, YAO N, LIN F, LIU Z, DONG Z, WANG L, PAN Q. Function and interaction of the coupled genes responsible forencoded rice blast resistance., 2014, 9(6): e98067.
[39] CAPALN J L, KUMAR A S, PARK E, PADMANABHAN M, HOBAN K, MODLA S, CZYMMEK K, DINESH-KUMAR S. Chloroplast stromules function during innate immunity., 2015, 34(1): 45-57.
(责任编辑 岳梅)
Construction of Rice Leaf sheath Protoplast Transformation System and Transient Expression of Pik-H4 and AvrPik-H4 Proteins
LIU Wei, LIU Hao, DONG ShuangYu, GU FengWei, CHEN ZhiQiang, WANG JiaFeng, WANG Hui
(National Engineering Research Center of Plant Space Breeding, South China Agricultural University, Guangzhou 510642)
The objective of this study is to obtain the suitable digestion and transformation time of protoplasts of rice sheath, improve the efficiency of transient expression, the target gene can be detected at the protein level and expressed in large quantities. To explore the feasibility of transient expression of rice blast resistance protein Pik1-H4, Pik2-H4 and avirulence protein AvrPik-H4 in protoplasts of rice leaf sheath, and to analyze the function of above target genes.High blast resistance rice variety H4 and control variety Zhonger Ruanzhan were used as experimental materials. Rice seedlings were cultured with 1/2 MS medium at 25℃ for 7-10 d. The protoplasts were isolated by cellulase and macerozyme enzymatic action. The optimal time of digestion was obtained by counting the number of cells in 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 and 12 h using hemocytometer. The target genesPik-H4,Pik-H4andwere fused with GFP to construct the transient expression vector. The total RNA was extracted from transformed protoplasts in 10, 12, 14, 16, 18, 20, 22 and 24 h, respectively. Real-time quantitative PCR (qRT-PCR) was used to detect the relative expression ofto obtain the best transformation time,housekeeping gene was used as control andspecific amplification primers were designed. The method of subcellular localization of the target gene by laser confocal scanning microscopy was used to estimate the gene function. The total protein was extracted and anti-GFP was used as the primary antibody, verified that the target protein successful expression by Western blot.rice seedlings were grown better quality and vitality in constant temperature 1/2 MS medium with rich and balanced nutrients, compared with soil planted at room temperature. Digested time had a greater impact on protoplast isolation efficiency. The results showed that the best time to digest was 4-6 h. The number of cells grew fastest at 3-4 h, tended to be stable at 4-6 h, showed a downward trend after 6 h, cell death rate accelerated and observed debris increased in the microscopic cell after 7 h. By detecting the relative expression of, it was found that the most suitable time for transformation was 14-16 h, reached the highest value at 16 h, and then gradually decreased. Subsequently, fluorescence of the GFP protein was observed to be quenched by fluorescence microscopy. Subcellular localization observation of AvrPik-H4 protein was mainly located in the cell membrane, presumably this is a membrane protein that is transported by some form to the host cell as an exciton to trigger a series of reactions.is composed ofPik-H4andPik-H4, which is highly efficient broad-spectrum rice blast gene. Pik1-H4 and Pik2-H4 were mainly located in the endoplasmic reticulum and plastid, respectively. From the subcellular localization results, it was presumed that Pik1-H4 might be mainly involved in the recognition of Avr-Pik protein and signal transmission, Pik2-H4 mainly play a role in changing the energy transmission and regulation of downstream disease caused hypersensitive reaction. Western blot results showed that the target protein was successfully expressed and the molecular size was correct. The expression of Pik1-H4 and AvrPik-H4 was higher than that of Pik2-H4, indicating that the size of the molecular weight is not a key factor affecting the transforming efficiency.The protoplast transient expression system of rice leaf sheath has the characteristics of high efficiency and rapidity, the exploration of protoplast isolation and transformation time has laid a foundation for the extensive practice of rice transient expression system. The successful expression of the target gene has provided a valuable theoretical basis for the study of the interaction mechanism between Pik-H4 and Avr protein.
rice; protoplast; subcellular localization; Western blot
2017-06-12;
2017-07-20
国家重点研发计划(2017YFD0100100)、国家“863”计划(2012AA101201)、国家自然科学基金(31401722)、广东省省级科技计划(2016A020210064)
联系方式:刘维,Tel:18819266052;E-mail:1158397772@qq.com。通信作者王慧,Tel:020-85283237;E-mail:wanghui@scau.edu.cn
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
作物学报(2022年6期)2022-04-08
新农业(2020年21期)2020-11-19
农村百事通(2019年8期)2019-05-13
农民文摘(2019年7期)2019-01-14
浙江中医杂志(2019年5期)2019-01-06
江苏农业科学(2018年21期)2018-12-05
农机化研究(2018年2期)2018-06-05
食品工业科技(2014年23期)2014-03-11
作物研究(2014年6期)2014-03-01
