
中华绒螯蟹微孢子虫研究进展
2017-12-14 05:52:53刘洪岩赵彦华孙梦玲薛晖
水产养殖 2017年10期
刘洪岩,赵彦华,孙梦玲,薛晖
(江苏省淡水水产研究所,江苏 南京 210017)
中华绒螯蟹微孢子虫研究进展
刘洪岩,赵彦华,孙梦玲,薛晖
(江苏省淡水水产研究所,江苏 南京 210017)
微孢子虫是专性细胞内寄生的真核生物,有着广泛的宿主。中华绒螯蟹微孢子虫可以引起中华绒螯蟹微孢子虫病,因此而受到重视。本文简要综述了中华绒螯蟹微孢子虫的生物学特性,检测方法以及中华绒螯蟹微孢子虫病的研究现状,以期为有效防治河蟹微孢子虫病提供参考。
中华绒螯蟹;微孢子虫;研究进展
微孢子虫是很大的一个门类,最开始被认为是寄生虫,直至2002年才重新被划分到真菌界[1]。目前发现的小孢子虫目微孢子虫有1 200个,分别属于150个种[2]。在中国,对微孢子虫的研究主要集中在昆虫宿主,特别是蚕[3-4]和蜜蜂[5-6]。目前发现,只有少数几种微孢子虫,如 Ameson[7],Nadelspora[8]和 A-belspora[9]是以螃蟹为宿主的。2002年,王文等[10]通过电镜观察,确定了江苏泗洪的螃蟹中含有微孢子虫,并且确定了微孢子虫病发病的不同时期的特性以及中华绒螯蟹微孢子虫肝胰腺寄生的特性。2006年,王文等[11]通过透射电镜和扫描电镜进一步分辨出了河蟹微孢子虫与Partrick 2002年提出的河蟹微孢子虫的不同,从而建立了一个新的分支。2015年,江苏省河蟹养殖过程中,肝胰腺的白化病大规模暴发,丁正峰[12]从病蟹中检测到大量微孢子虫18s rDNA。
1 河蟹微孢子虫的生物学特性
1.1 河蟹微孢子虫的超微结构
河蟹微孢子虫的形状为椭圆形,短轴长度为0.8~1.2μm,长轴长度为1.5~1.9μm。孢子壁厚度为130 nm,由低电子密度的孢子内壁和高电子密度的孢子外壁组成,孢子内壁厚度为100~120 nm,孢子外壁厚度为20~30 nm。河蟹微孢子虫是单核的,核位于极质体和后极泡之间,在细胞中央的位置,由两到三层内质网极管以及极管包围。在孢子的前端,有一个钟形的锚定盘,锚定盘前端有凸起;孢子后端则包含这2~3个后极泡。河蟹微孢子虫主要特点是:单核,有一个极丝,一个极丝盘复合物,一个极质体以及后极泡形成发芽装置;极管圈数为7圈。见图1。
1.2 河蟹微孢子虫生活史
河蟹微孢子虫寄生于中华绒螯蟹的肝胰腺细胞内,生活史包括四个阶段:裂殖增殖期,孢子增殖期,孢子原细胞期和感染期。第一个时期是从成熟的孢子细胞质由极管进入宿主细胞开始的。孢原质进入宿主细胞以后发育形成椭圆形的裂殖子。接下来就是裂殖增殖期。这个阶段,裂殖子在宿主细胞内不断分裂,形成多个新的裂殖子,并且发育成为成熟的孢子。这两个时期都是在宿主细胞的细胞质中进行的。成熟的孢子由发芽装置弹射出,继续感染新的细胞[13],见图2。

图1 微孢子虫模式图[11]
1.2.1 裂殖增殖期 这个时期从微孢子虫的原生质进入宿主细胞便开始了。原生质的细胞核由单核分裂成为双核,四核,进而形成多核,这时的细胞是椭圆形的,具有很大的核区。随后,原生质的质膜内陷,把细胞核包裹起来,形成新的原生质。微孢子虫的原生质会进行多次这样的分裂,从而形成多个原生质体。这些原生质体开始排列成莲花形,随后,分裂成为4个或者8个新的原生质。这种原生质体继续发育,在寄生虫样空泡中发育成为单层膜包围的原生质体。这时的原生质体富含游离的核糖体,细胞核为均匀的颗粒状,质膜周围富含线粒体。
1.2.2 孢子发生期 这一阶段是由裂殖体向孢子母细胞发育的时期。孢子母细胞是成熟的、不再继续分裂的孢子。孢子母细胞是单核的,由多核的原生质分裂团分裂产生。电子显微镜观察显示,当裂殖体的质膜外面有一层致密的高电子层时,就发育为成熟的孢子,即孢子母细胞。孢子母细胞和裂殖体相比,细胞质的电子密度也有所增加,因为在孢子母细胞中,有大量的核糖体和内质网形成。
中华绒螯蟹微孢子虫之所以成为新的一类,是因为与 Endoreticulatus wdelis[14],E.schuberg[15],E.bombycis[16-17]and Endoreticulatus sp.Taiwan[18]相比,有着以下三点明显的不同:①中华绒螯蟹微孢子虫比较小,(1.0±0.2)μm×(1.7±0.2)μm;②孢子成熟和孢子生殖同时在宿主的细胞质中进行,这与网虫属Ocinaralida是不同的,后者表现出不同发育阶段的寄生虫空泡[17];③中华绒螯蟹微孢子虫的极丝,核膜,细胞核在早期的孢子发育阶段都是可见的,而这些在Endoreticulatus中则是很难发现的。
2 河蟹微孢子虫的检测方法
目前,河蟹微孢子虫还没有被提纯出来,现阶段的检测方法主要集中在组织病理切片和分子检测手段上面。
2.1 河蟹微孢子虫的组织病理学切片检测
HE染色是观察病理的主要方法,可以直观的看到微孢子虫的含量以及对组织的破坏程度。目前主要有马松三色法和HE染色法两种[12]。前者可以根据染料渗入程度分辨不同的粒子,后者则是根据嗜酸性和嗜碱性来区分不同粒子的。
2.2 电镜切片观察
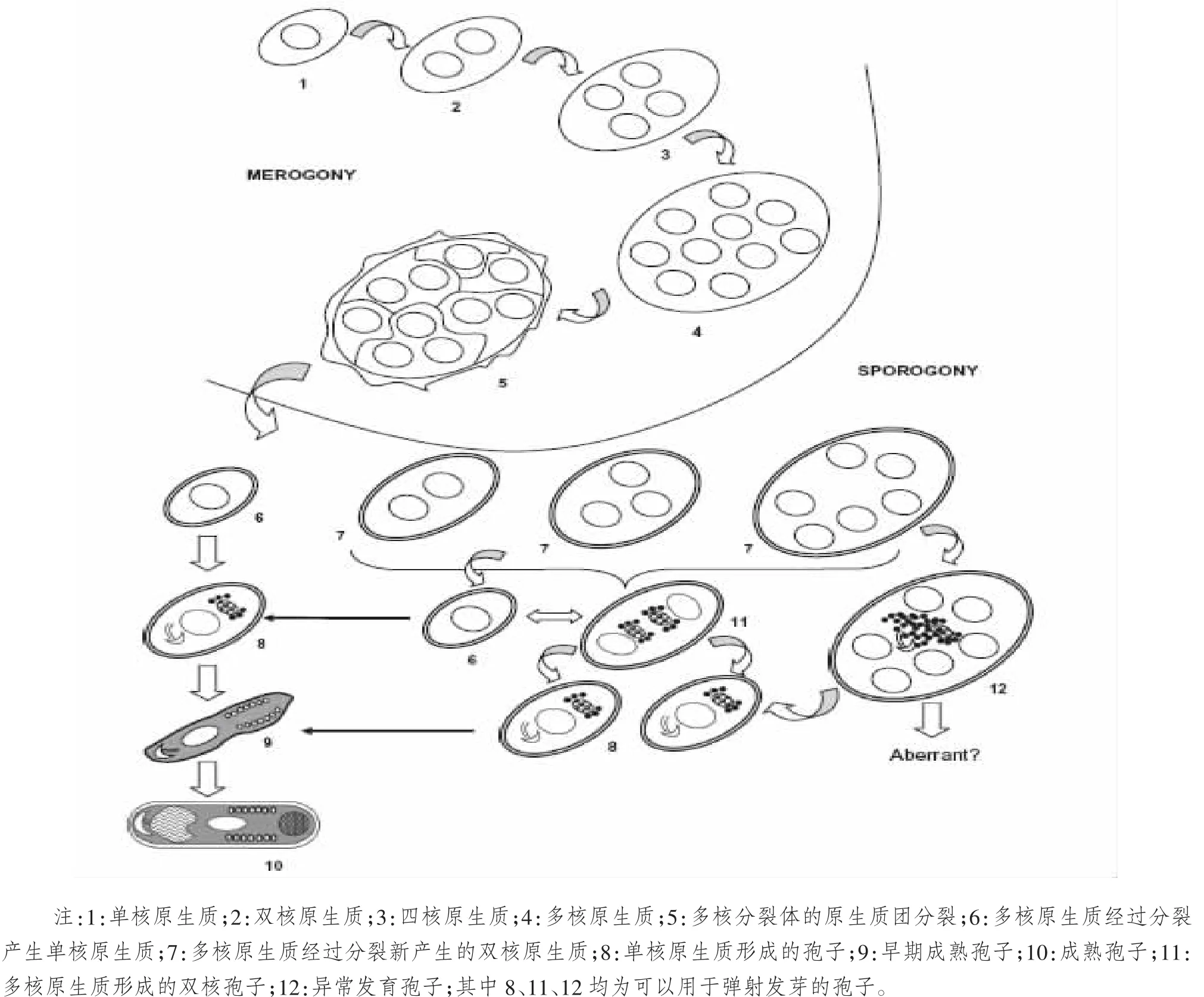
图2 河蟹微孢子虫的生活史[13]

图3 白化肝胰腺组织化学切片[21]
成熟的河蟹微孢子虫主要由孢子壁、孢原质、发芽装置和孢子细胞器等组成。孢子壁厚而坚硬,由3层结构组成,由外向内依次为刺突状外层孢子外壁、透明薄层孢子内壁和纤维性内层的原生质膜。孢子壁厚度为130 nm,其中,孢子内壁厚度为100~120 nm,孢子外壁厚度为20~30 nm。孢原质的前部是薄膜层叠成的极膜层,孢原质的后部有沿孢子短轴方向稍伸长的孢子。发芽装置主要由孢子壁内侧螺旋状盘绕的极管、若干叠成片状的薄膜层、结构相对松散的极膜层和由多层膜包围成的后极泡构成。孢子细胞质内可见有内质网、极膜层结构、高尔基体和线粒体等细胞器。河蟹微孢子虫的成熟孢子大小为(1.7±0.2)μm ×(1.0±0.2)μm,呈椭圆形,极管前端与孢子纵轴呈平行状,后端沿孢子壁与纵轴形成螺旋状卷曲,极丝的极管圈数一般为7~8 圈,见图 4。
2.3 常规PCR检测和荧光PCR检测
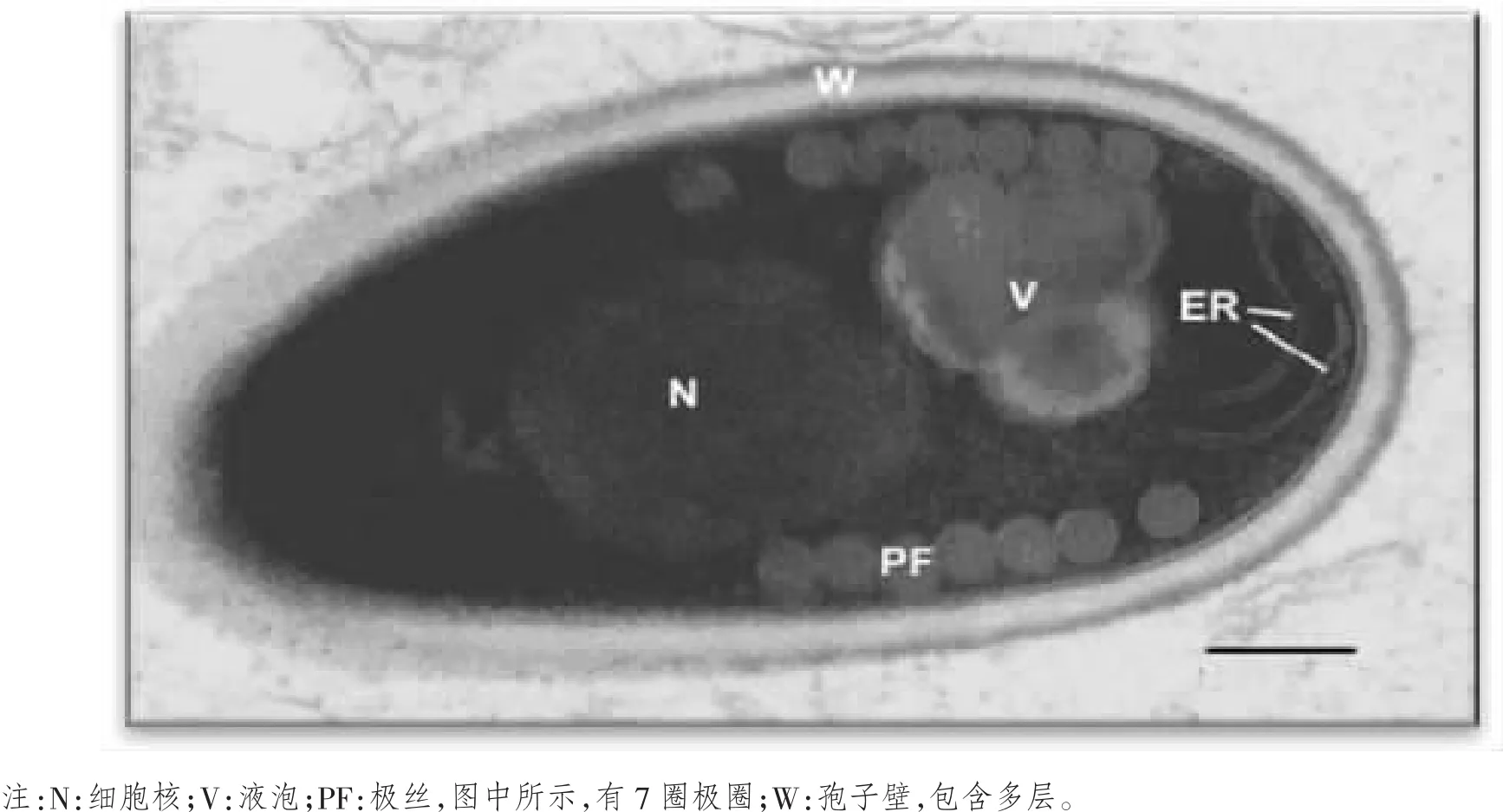
图4 河蟹微孢子虫纵轴切面的透射电镜照片[11]
检测方法目前采用PCR方法,检测河蟹微孢子虫时,主要针对微孢子虫16sRNA基因来设计特异性引物[19]。丁正峰利用巢式PCR扩增出931 bp的片段,经比对确认为河蟹微孢子虫18sRNA。
常规PCR的检测,对于低浓度的微孢子虫检出率不高,经过笔者实验,常规PCR只能检测到102以上的微孢子虫。因此,需要更加灵敏的检测方法。荧光定量PCR满足了这一需求。荧光定量PCR的检出线为101,并且能够确定微孢子虫的含量,确定微孢子虫在河蟹体内的分布情况。为微孢子虫的检测提供了更加灵敏的方法,也为后续研究提供了一种新途径。
2.4 原位杂交
原位杂交是指将特定标记的已知顺序核酸为探针与细胞或组织切片中核酸进行杂交,从而对特定核酸顺序进行精确定量定位的过程。丁正峰[16]设计 了 两 对 引 物 W3F (50-CTGTGAGGCTATTGTTGGGC-30)and W3R(50-TACTGTGCTCCCTGTCCATT-30),对白化的肝胰腺进行了原位杂交。对比HE染色和直接显微镜观察的结果,原位杂交能够更加灵敏地显示出低浓度的微孢子虫。
3 螃蟹微孢子虫病
早 在 1970年 ,Sprague就 指 出 ,Ameson michaelis是引起墨西哥湾,亚特兰大的海蟹肌肉溶解的病因。这种微孢子虫存在于海蟹的肌肉中,会使螃蟹的肌肉成为棉花样,并且失去原有的味道,被称为“棉花蟹”。Ameson michaelis[3]的传染性很高,但是并没有引起“棉花蟹”的大规模传播。
1987年,另外一种螃蟹微孢子虫被发现。Azevedo发现的这种寄生在岸蟹肝胰腺中的微孢子虫叫做Abelspora portucalensis,这种微孢子虫是单核的,以二分式的裂殖方式繁殖的。每个微孢子虫可以裂殖为两个,两个再分为四个,这一过程是在微孢子虫囊膜中进行的[9]。1994年,Olson等[8]在太平洋大蟹中发现了微孢子虫Nadelspora canceri。2007年,Stentiford G D.在寄生蟹中发现了微孢子虫Enterospora[20]。
但是,河蟹微孢子虫和上述的都不是一种。河蟹微孢子虫的孢子时期都是单核的,多核的孢子母细胞原生质体以莲花形式的分裂方式,分为4个或者8个单核孢子,这些孢子在寄生虫样的空泡中成熟。这种特征跟内网虫属比较相似,因此2007年,王文[11]建立了Endoreticulatus eriocheir sp.nov这一分支。2015年,江苏暴发了严重的河蟹肝胰腺白化病,丁正峰[12]在发病的河蟹肝胰腺中,发现了大量的微孢子虫,经过18sRNA的鉴定,确认为Endoreticulatus eriocheir sp.nov.。河蟹微孢子虫病的主要病症为肝胰腺白化,水化,进而消失,不会引起螃蟹的死亡,但是会严重影响螃蟹的品质。
目前,对河蟹微孢子虫和河蟹肝胰腺白化之间的关系,争议颇多。2016年,丁正峰[21]利用18sRNA设计引物,对白化的肝胰腺进行的原位杂交,在荧光显微镜下,可以看到阳性杂交信号。并且文中还指出,原位杂交能发现组织化学和普通显微镜观察所不能发现的,早期的、含量低的河蟹微孢子虫,见图5。
4 结语
河蟹微孢子虫是一种真核生物,内虫网属,生活史包括四个阶段:裂殖增殖期,孢子增殖期,孢子原细胞期和感染期。河蟹微孢子虫为单核,多核的孢子母细胞原生质经过莲花式的分裂,分裂为4个或者8个孢子。
河蟹微孢子虫病可以引起河蟹肝胰腺的白化,水化,最终肝胰腺消失,给螃蟹养殖也造成了很大的损失。目前,对于该病的研究集中在检测方法的建立上,现在已经能够成熟地利用PCR和荧光PCR检测河蟹微孢子虫的含量,用组织化学的方法直观地检测到微孢子虫,但是,对于感染途径等流行病学调查还没有相关文献报道,也没有特效药可以治疗。
今后,河蟹微孢子虫病的研究方向应主要集中在传染途径的确定以及防治特效药的研究方面。
[1]Peer Y V,Ali A B,Meyer A.Microsporidia:accumulating molecular evidence that a group of amitochondriate and suspectedly primitive eukaryotes are just curious fungi[J].Gene 2000,(246):1-8.
[2]Patrick,J K,Naomi M.Microsporidia:biology and evolution of highly reduced intracellular parasites[J].Annu.Rev.Microbiol,2002(56):93-116.
[3]贡成良.家蚕微孢子虫研究[J].江苏蚕业,1999(1):6-13.
[4]高永珍,黄可威.家蚕病原性微孢子虫的超微结构研究[J].蚕业科学,1999,25(3):163-169.
[5]刘锋,王强,代平礼,等.蜜蜂微孢子虫在中国的自然种系构成初探[J].应用昆虫学报,2008,45(6):963-966.
[6]欧阳红燕,刘玉梅,刘彩珍.蜜蜂微孢子虫病研究进展[J].养蜂科技,2002(6):17-19.
[7]Sprague V.Some protozoan parasites and hyperparasites in marine decapod Crustacea.In:Snieszko,S.F.(Ed.),A Symposium on Diseases of Fishes and ShellWshes[J].American Fisheries Society Special Publication,1970(5):416-429.
[8]Olson R,Tiekotter E,Reno P.Nadelspora canceri(n.g,n.sp.):an unusual microsporidian parasite of Dungeness crab(Cancer magister)[J].Eukaryot.Microbiol,1994(41):337-347.
[9]Azevedo C.Fine structure of the microsporidan Abelspora portucalensis gen.n.,sp.n. (Microsporida)parasite of the hepatopancreas of Carcinus maenas(Crustacea,Decapoda)[J].Invertebr.Pathol,1987(49):83-92.
[10]Wang W,Gu Z.Rickettsia-like organism associated with tremor disease and mortality of the Chinese mitten crab,Eriocheir sinensis[J].Dis.Aquat.Org,2002(48):149-153.
[11]Wang W,Chen J.Ultrastructural study on a novel microsporidian,Endoreticulatus eriocheir sp.nov.(Microsporidia,Encephalitozoonidae),parasite of Chinese mitten crab,E-riocheir sinensis(Crustacea,Decapoda)[J].Journal of invertebrate pathology,2007(94):77-83.
[12]Ding Z,Meng Q,Liu H.First case of hepatopancreatic necrosis disease in pond-reared Chinese mitten crab,Eriocheir sinensis,associated with microsporidian[J].Journal of fish diseases.doi:10.1111/jfd.12437.
[13]Stentiford G D.Hepatospora eriocheir(Wang and Chen,2007)gen.et comb.nov.infecting invasive Chinese mitten crabs(Eriocheir sinensis)in Europe[J].Journal of Invertebrate Pathology,2011(108):156-166.
[14]Brooks W M,Becnel JJ,Kennedy G G.Establishment of Endoreticulatus n.g.for Pleistophora fidelis(Hostounskamp;Weiser,1975) (Microsporida:Pleistophoridae)based on the ultrastructure of a microsporidium in the Colorado potato beetle,Leptinotarsa decemlineata(Say)(Coleoptera:Chrysome)[J].Journal of Eukaryotic Microbiology,2010,35(4):481-488.[15]Cali A,Garhy M E.Ultrastructural Study of the Development of Pleistophora schubergi Zwelfer,1927 (Protozoa,Microsporida)in Larvae of the Spruce Budworm,Choristoneura fumiferana and Its Subsequent Taxonomic Change to the Genus Endoreticulatus[J].Journal of Eukaryotic Microbiology,1991,38(3):271-278.
[16]Wan J,Zhang L,Chen Z,et al.Study of a pathogenic microsporidium SCM 7(Endoreticulatus sp)isolated from the larvae of silkworm,Bombyx mori[J].Acta Sericologica Sinica,1995,21(3):168-172.
[17]Zhang L,Du Y,Wang Y.A new species of Endoreticulatus(Protozoa,Microspore)from the larva of silkworm,Bombyx mori.L[J].Journal of Southwest Agricultural University,1995(17):363-367.
[18]Wang CY,Solter L F,Sui W H,et al.An Endoreticulatus,species from Ocinara lida,(Lepidoptera:Bombycidae)in Taiwan[J].Journal of Invertebrate Pathology,2005,89(2):123.
[19]Vossbrinck,C.R.,Debrunner-Vossbrinck,B.A.Molecular phylogenyof the Microsporidia:ecological,ultra structural and taxonomic considerations[J].Folia Parasitol,2005(52):131-142.
[20]Stentiford G D.Enterospora sp.an intranuclear microsporidian infection of hermit crab Eupagurus bernhardus[J].Diseases of Aquatic Organisms,2007,75(1):73-78.
[21]Ding Z F,Chen JQ,Lin J,et al.Development of In situ hybridization and real-time PCR assays for the detection of Hepatospora eriocheir,a microsporidian pathogen in the Chinese mitten crab Eriocheir sinensis [J].Journal of Fish Diseases,2016(40):919-927.
Research progress on the Endoreticulatus eriocheir sp.nov.(Microsporidia,Encephalitozoonidae)
Liu Hongyan,Zhao Yanhua,Sun Mengling,Xue Hui
(Freshwater Fishery Research Institute of Jiangsu Province,Nanjing 210017,China)
Microsporidia are an extremely large and diverse group of microbial eukaryotes and have wide host area.For emerging pathogen in Eriocheir sinensis,more attention had been put in it.In order to provide a reference for controlling microsporidian infection effectively,the structure,detecting technology and progress of studying on the Endoreticulatus eriocheir sp.nov.are reviewed in this paper.
Eriocheir sinensis;microsporidia;research progress
S941.5
A
1004-2091(2017)10-0042-06
10.3969/j.issn.1004-2091.2017.10.009
江苏省科技计划现代农业项目(BE2016392)
刘洪岩(1986-),女,研究实习员,主要从事水产病害研究.E-mail:njsfdxliuhongyan@163.com
薛晖(1968-),男,研究员级高级工程师.E-mail:jsxuehui@163.com
图5 河蟹白化肝胰腺原位杂交
2016-12-26)
猜你喜欢
食用菌(2017年5期)2017-10-19 03:02:28
特产研究(2016年3期)2016-04-12 07:16:32
首都医科大学学报(2015年4期)2015-12-16 13:00:08
大连工业大学学报(2015年4期)2015-12-11 04:06:50
特产研究(2015年1期)2015-04-12 06:36:11
中国当代医药(2015年17期)2015-03-01 02:03:43
郑州大学学报(医学版)(2015年1期)2015-02-27 14:50:35
食品工业科技(2014年23期)2014-03-11 18:19:06
无机化学学报(2014年12期)2014-02-28 17:33:53
无机化学学报(2014年7期)2014-02-28 17:32:11
