
南洋楹溃疡病病原菌生物学特性研究
2017-12-14 05:54:08王伟王新荣曾炳山刘英范春节纪春艳
中国森林病虫 2017年6期
王伟,王新荣,曾炳山,刘英,范春节,纪春艳
(1.华南农业大学农学院/广东省微生物信号与作物病害重点实验室,广东 广州 510642; 2.中国林业科学院热带林业研究所,广东 广州 510000)
南洋楹溃疡病病原菌生物学特性研究
王伟1,王新荣1,曾炳山2,刘英2,范春节2,纪春艳1
(1.华南农业大学农学院/广东省微生物信号与作物病害重点实验室,广东 广州 510642; 2.中国林业科学院热带林业研究所,广东 广州 510000)
研究了不同培养基、碳氮源、温度、pH及光照条件对南洋楹溃疡病病原菌可可毛色二孢Lasiodiplodiatheobromae菌丝生长的影响。结果表明:该菌的最适培养基为PDA,最适碳源、氮源分别是葡萄糖和蛋白胨,菌丝最适生长温度为25 ℃,最适pH为5~7,在12∶12光暗交替条件下生长最好。
南洋楹;溃疡病;可可毛色二孢;生物学特性
南洋楹Falcatariamoluccana是含羞草科Mimosaceae南洋楹属的常绿乔木,自然分布于马来西亚马六甲和印度尼西亚,是世界著名的热带速生树种之一。南洋楹木材纤维丰富,韧性强,易加工,是人造板、制浆造纸的优良原料。南洋楹树形优美,根系富含根瘤菌,固氮力强,是风景园林、改良土壤的优良树种[1]。
南洋楹人工林具有经营周期短,经济效益显著等特点。目前,南洋楹人工林在广东增城等地已形成规模化种植,促进了当地林农的增收致富。国内对于南洋楹的研究大多集中在引种、栽培、优良种源/家系的选育上[2-5],有关南洋楹病害的研究很少报道。张景宁等[6]报道了南洋楹丛枝病的发生。2013年作者在广东增城惠东及博罗的南洋楹种植区发现了一种溃疡病,严重影响南洋楹的生长及品质。经调查,该病害主要为害1~2 a生南洋楹植株,感病枝条呈现棕褐色或黑褐色小斑,干部常有黑褐色的凹陷溃疡大病斑,感病植株生长不良,有的落叶、顶枯,严重时甚至死亡。作者对采集的感病样本进行了病原菌分离培养、纯化及鉴定,分别通过对南洋楹树苗(苗龄2月)及南洋楹树木(1 a生)接种代表优势病原菌菌株(BL1331和HD1332)的科赫法则致病性验证,结合菌株(BL1331和HD1332)的形态鉴定、分子鉴定和基于核糖体内转录间隔区(ribosomal internal transcribed spacer region,ITS)、β-微管蛋白(β-tubulin)及延长因子(elongation factor 1-α,EF 1-α)的序列比较和多基因系统进化树的构建分析,首次系统报道了由可可毛色二孢Lasiodiplodiatheobromae引起南洋楹溃疡病的发生[7]。本文作者对南洋楹溃疡病优势病原菌可可毛色二孢的生物学特性进行研究,以期对该病害的科学防控提供依据。
1 材料与方法
1.1 病原菌株的分离纯化 病样从增城博罗及惠东两地林场里发病严重且具典型症状的南洋楹病树上采集,并进行病原菌的分离、纯化。在感病组织的病、健交界处取一小块组织,用75 %乙醇浸泡30 s,后浸泡于10 %次氯酸钠溶液2 min,无菌水冲洗5次,无菌滤纸吸干,接种于PDA培养基平板上,25 ℃培养,经单孢分离,获得单孢纯化菌株,选取从博罗分离获得的菌株BL1331和从惠东分离获得的菌株HD1332作为供试菌株进行生物学测定。
1.2 培养基配方 生物学测定采用下述6种培养基。PDA(马铃薯葡萄糖培养基):马铃薯200 g,葡萄糖15 g,琼脂20 g,补足水至1 000 mL;Czapek(查氏培养基):硝酸钠3 g,磷酸氢二钾1 g,硫酸镁0.5 g,氯化钾0.5 g,硫酸铁0.01 g,蔗糖30 g,琼脂20 g,补足水至1000 mL;OA(燕麦培养基):燕麦片20 g,葡萄糖15 g,琼脂20 g,补足水至1 000 mL;CMA(玉米粉琼脂培养基):玉米粉300 g,琼脂20 g,补足水至1 000 mL;Plant tissue(南洋楹汁液培养基):健康新鲜南洋楹枝叶200 g,用水熬煮20 min,过滤,加入琼脂20 g,补足水至1 000 mL;WA(水琼脂培养基):琼脂15 g,补足水至1 000 mL。
1.3 病原菌生物学特性测定
1.3.1 不同培养基对菌株菌丝生长的影响 将PDA平板上培养3 d的菌株沿菌落边缘用打孔器打取直径6 mm的菌饼,分别接种于PDA、Czapek、OA、CMA、Plant tissue、WA等6种平板培养基中央,置于28 ℃恒温培养箱中光暗交替培养,观察菌落特征,采用十字交叉法测量菌落直径,以两次测量的平均值代表该平皿的菌落直径,每个处理重复3次。
1.3.2 不同C源对菌株菌丝生长的影响 以查氏培养基为基础培养基,分别加入等量蔗糖、果糖、乳糖、葡萄糖、麦芽糖及可溶性淀粉作为碳源制成相应的平板培养基,以不加蔗糖的查氏培养基为碳源对照。打取直径6 mm的菌饼分别接种于上述培养基中央,28 ℃恒温光暗交替培养,观察菌落生长情况,采用十字交叉法测量菌落直径,每个处理重复3次。
1.3.3 不同N源对菌株菌丝生长的影响 以查氏培养基为基础培养基,分别加入等量的硫酸铵、硝酸铵、氯化铵、蛋白胨、牛肉膏、尿素以代替硝酸钠作为氮源制成相应的平板培养基,以不加硝酸钠为氮源对照。打取直径6 mm的菌饼分别接种于上述培养基中央,28 ℃恒温光暗交替,观察菌落生长情况,采用十字交叉法测量菌落直径,每个处理重复3次。
1.3.4 不同温度对菌株菌丝生长的影响 以查氏培养基为基础培养基,打取直径6 mm的菌饼接种至平板的中央,分别置于5,10,15,20,25,30,35,40 ℃中光暗交替培养,观察菌落生长情况,采用十字交叉法测量菌落直径,每个处理重复3次。
1.3.5 不同光照对菌株菌丝生长的影响 以查氏培养基为基础培养基,打取直径6 mm的菌饼接种至平板的中央,分别置于24 h光照(光照强度为300 lx)、24 h黑暗及12∶12(L∶D)3种光照条件下,28 ℃恒温培养,观察菌落生长情况,采用十字交叉法测量菌落直径,每个处理重复3次。
1.3.6 不同pH对菌株菌丝生长的影响 用1 mol/L的HCl和NaOH调配pH为3,4,5,6,7,8,9,10,11,12的查氏培养基,打取直径6 mm的菌饼接种至平板的中央,28 ℃恒温培养,观察菌落生长情况,十字交叉法测量菌落直径,每个处理重复3次。
1.4 数据分析 试验数据采用SPSS软件Duncan新复极差法进行统计分析。
2 结果与分析
2.1 分离菌株的形态特征 供试菌株BL1331和HD1332在PDA培养基平板上生长速度较快,气生菌丝发达,菌落初期为白色,绒毛状,边缘整齐,培养后期菌丝逐渐变为灰色,背面呈黑色,菌丝体呈束状耸立(图1)。供试菌株的分生孢子未成熟时单细胞,无色,薄壁;成熟时暗褐色,双细胞,椭圆形至卵圆形,厚壁(图2)。
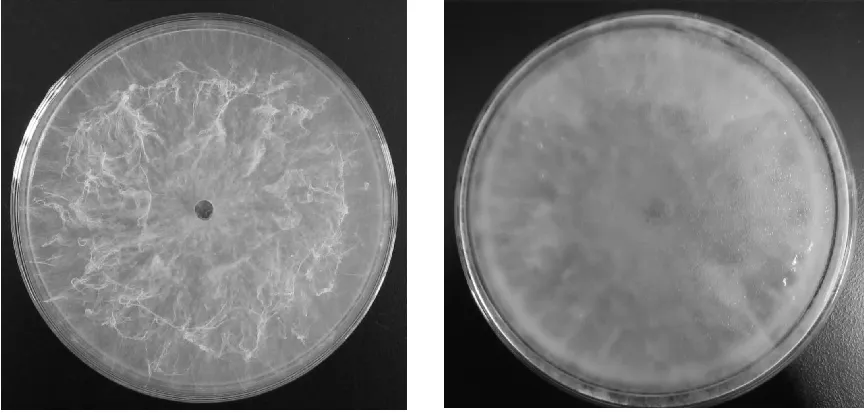
图1 菌株BL1331在PDA平板上的菌落形态(左:初期;右:后期)
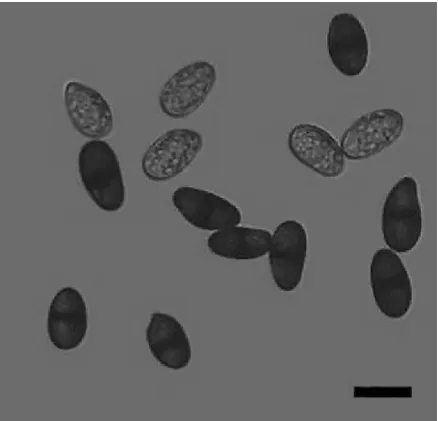
图2 菌株BL1331的分生孢子(标尺20 μm)
2.2 不同培养基对供试菌株菌丝生长的影响 病原菌菌株BL1331和HD1332在不同培养基上的菌丝生长存在一定差异。培养48 h后,两个菌株在PDA上生长速度最快,菌丝茂盛,菌落直径明显大于在其它培养基上的菌落直径。在查氏、玉米粉、燕麦及南洋楹汁液培养基上菌丝的生长速度依次递减,在水琼脂培养基上生长最慢,菌落最小(表1)。

表1 不同培养基对菌株菌丝生长的影响
注:数值为3个重复的平均值,同列中不同小写字母和大写字母分别表示5%和1%的差异显著水平。
2.3 不同碳源对供试菌株菌丝生长的影响 病原菌菌株BL1331和HD1332在所有供试的碳源培养基上均能够生长,但对不同碳源的利用能力存在一定差异(图3)。在含有葡萄糖和蔗糖的培养基上菌丝生长较快,培养48 h时菌株BL1331菌落直径分别为对照的1.93和1.91倍,而菌株HD1332菌落直径分别为对照的1.92和1.76倍。在含有乳糖、麦芽糖和可溶性淀粉的培养基上生长速度次之,在果糖上生长速度较慢,在不加碳源的对照上生长速度最慢。结果表明,葡萄糖和蔗糖是菌丝生长的最佳碳源。
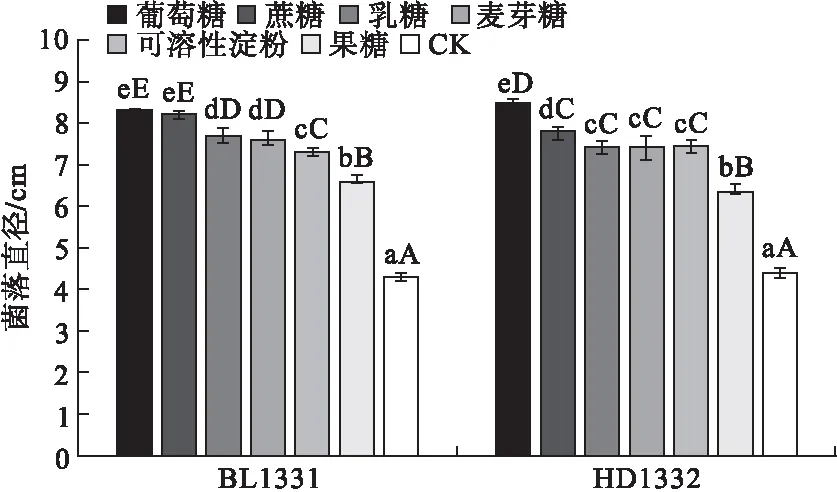
图3 不同碳源对菌株菌丝生长的影响
2.4 不同氮源对供试菌株菌丝生长的影响 病原菌菌株BL1331和HD1332对不同氮源的利用能力存在一定差异(图4)。培养48 h时,两个菌株在含有蛋白胨的培养基上菌丝生长最快,在含有硝酸铵的无机氮源培养基上菌丝的生长速度较慢,而在含有尿素的培养基上几乎不生长。结果表明,蛋白胨是菌丝生长的最佳氮源,而尿素不利于菌株生长。不同氮源培养基上菌落形态也稍不同,在含有蛋白胨、牛肉膏等有机氮源的培养基上菌落较致密,菌丝茂盛,而在硝酸铵为氮源的培养基上菌丝稀疏(图5)。
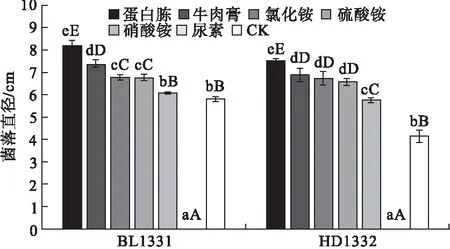
图4 不同氮源对菌株菌丝生长的影响
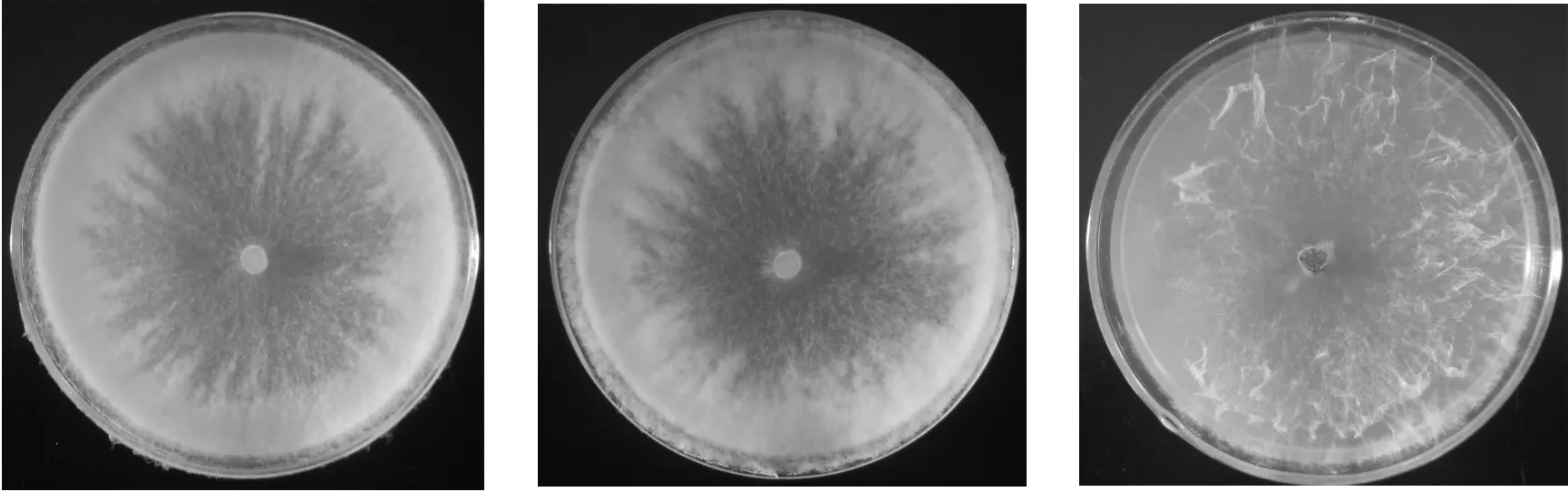
图5 不同氮源培养基上菌株BL1331的菌落形态(左:蛋白胨;中:牛肉膏;右:硝酸铵)
2.5 不同温度对供试菌株菌丝生长的影响 温度对病原菌菌株BL1331和HD1332的菌丝生长都具有显著影响。两个菌株在10~35 ℃的范围内均能够生长,在25~30 ℃范围内菌丝生长较快,显著高于其它温度区域。在15 ℃和35 ℃生长速度较慢,在10 ℃条件下生长缓慢,在5 ℃和40 ℃的条件下不生长(表2)。结果表明,最适宜该菌菌丝生长的温度范围在25~30 ℃,低于5 ℃和高于40 ℃均不能生长。
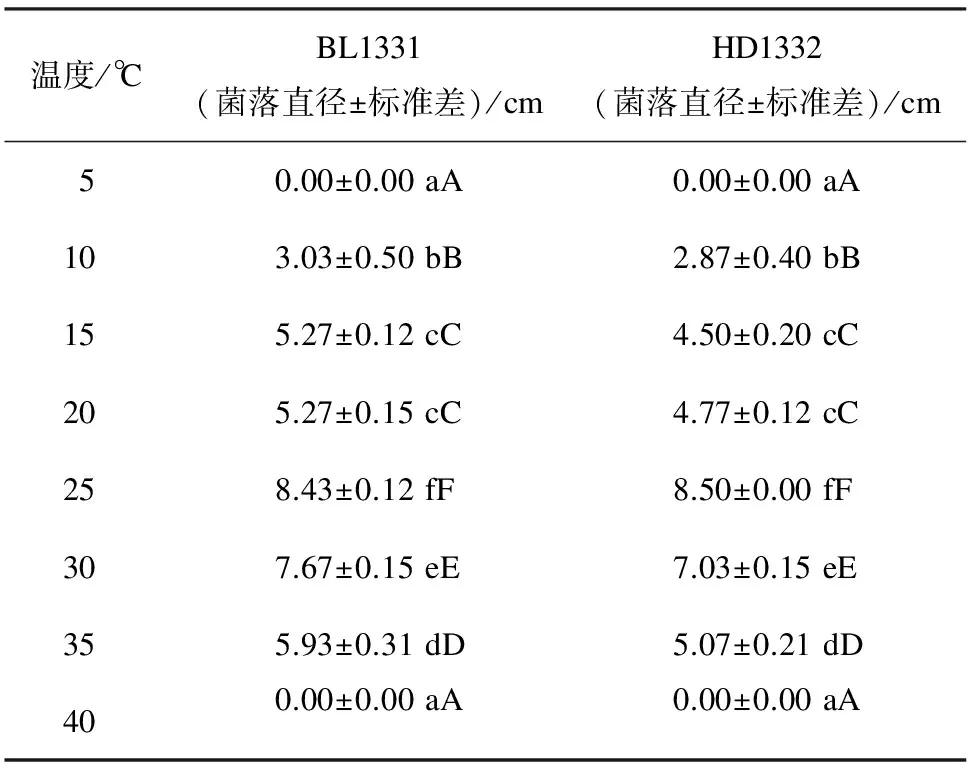
表2 不同温度对菌株菌丝生长的影响
注:数值为3个重复的平均值,同列中不同小写字母和大写字母分别表示5%和1%的差异显著水平。
2.6 不同光照对供试菌株菌丝生长的影响 不同光照条件对病原菌菌丝的生长具有影响,菌株BL1331和HD1332在完全光照和光暗交替的条件下生长较为快速,在完全黑暗条件下生长较慢(表3)。结果表明,光暗交替最有利于病原菌的菌丝生长。

表3 光照对菌株菌丝生长的影响(PDA,25 ℃)
2.7 不同pH对菌株BL1331和HD1332菌丝生长的影响 供试菌株的菌丝在pH 4~11的范围内均能够生长。菌丝在pH为5~7的范围内生长最快,在pH 7~11的范围内生长速度有所减缓(图6),说明微偏酸条件更有利于该病原菌的生长。

图6 不同pH对菌株菌丝生长的影响
3 结论与讨论
本次试验研究结果表明:最适合该病原菌菌丝生长的培养基为PDA,最适碳、氮源分别为葡萄糖和蛋白胨。在供试碳源中,低聚糖(如葡萄糖和蔗糖)有利于病原菌的营养生长。在供试氮源中,有机氮源有利于病原菌的营养生长,而无机氮源相对较差,尿素则难以利用,与沙田柚果腐病(citusmaximafruit rot)菌的研究结果一致[8]。病菌在pH 4~11的范围内都能够生长,最适pH为5~7,在12∶12光暗交替条件下生长最好,在全光照、全黑暗条件下均能生长,表明该病菌对环境的适应性较强。在5 ℃条件下,病原菌菌丝不能生长。在10 ℃条件下,菌丝生长十分缓慢,表明低温可减缓该病害的发生。最适合病原菌菌丝生长的温度为25~30 ℃,同毛葡萄穗轴褐腐病(vitisheyneanabrown rachis rot)病原菌菌丝生长的最适温度范围一致[9]。在25~35 ℃的条件下,病原菌菌丝生长较好,表明高温较低温更适宜病原菌菌丝的生长,在高温条件下可能仍具有较强的存活力而有利于病害的流行蔓延,此特点与林间高温多雨时发病较重相一致。在生产实践中,通过控制环境条件减少病原菌,从而减缓病害的发生,也能够为生长季节预测病害的发生和及时防控提供基础的依据。
可可毛色二孢主要分布在热带、亚热带地区,寄主广泛,能够引起500多种植物病害,导致溃疡、枝枯、坏死、果腐等症状[10]。它是一类弱寄生菌,Jami等研究报道[11]逆境胁迫条件有利于其成功侵染植物。南洋楹溃疡病是近年南洋楹种植业的重要病害,其发病流行规律尚不明确。我们在增城博罗和惠东南洋楹林场的病害调查初步发现,林间虫害频发、温暖多雨的环境,尤其是台风后造成林木风折伤口的条件下,病害易严重发生。有必要在南洋楹溃疡病病原菌生物学特性的研究基础上,系统开展该病害发生、发展及流行规律的研究,结合科学种植、改变环境,药剂筛选等综合防控措施,积极有效地控制南洋楹溃疡病的进一步发生及蔓延。
[1] SIREGAR U J,RACHMI A,MASSIJAYA M Y,et al.Economic analysis of sengon (Paraserianthesfalcataria) community forest plantation,a fast growing species in East Java,Indonesia[J].Forest Policy and Economics,2007,(9):822-829.
[2] 郑永光,周小珍,陈红跃,等.南洋楹优良种源/家系选择的研究[J].华南农业大学学报,2004,25(3):29-33.
[3] 韦如萍,胡德活,郑永光,等.南洋楹种源家系试验[J].中南林业科技大学学报(自然科学版),2007,27(1):64-69.
[4] 韦如萍,晏姝,胡德活,等.11年生南洋楹引种试验及家系选择[J].中南林业科技大学学报,2012,32(5):19-24.
[5] 刘英,曾炳山,裘珍飞,等.基质对南洋楹组培苗移植成活的影响[J].中南林业科技大学学报,2013,33(11):29-33.
[6] 张景宁,吴爱萍,程伟文,等.南洋楹丛枝病病原的研究[J].广东林业科技,1994,(2):24-26.
[7] JI C Y,CHEN C J,WANG X R,et al.A report on canker disease ofFalcatariamoluccanacaused byLasiodiplodiatheobromaein China[J].Crop Protection,2017,91:89-92.
[8] 董章勇,罗梅,宾淑英,等.沙田柚果腐病病原菌的鉴定与生物学特性[J].中国农学通报,2013,29(22):125-128.
[9] 史国英,胡春锦,罗掉爱,等.毛葡萄穗轴褐腐病病原菌鉴定及其生物学特性[J].植物病理学报,2010,40(3):242-249.
[10]URBEZ-TORRES J R,LEAVITT G M,VOEGEL T M,et al.Identification and distribution ofBotryosphaeriaspp.associated with grapevine cankers in California[J].Plant Disease,2006,90:1490-1503.
[11]JAMI F,SLIPPERS B,WINGFIELD M J,et al.Greater Botryosphaeriaceae diversity in healthy than associated diseasedAcaciakarrootree tissues[J].Australasian Plant Pathology,2013,42:421-430.
(责任编辑 李计顺)
BiologicalcharacteristicsofthepathogenLasiodiplodiatheobromaefromthecankerdiseaseinFalcatariamoluccana/
WANG Wei,et al.
(College of Agriculture,South China Agricultural University,Guangdong Provincial Key Laboratory for Microbial Signals and Crops Disease Control,Guangzhou 510642,China)
Biological characteristics ofLasiodiplodiatheobromaecausingFalcatariamoluccanastem canker disease were studied.The results showed that the optimal media for mycelium growth was PDA.The most suitable carbon and nitrogen source were glucose and peptone.The optimum temperature was 25℃.The most suitable pH value was from 5.0 to 7.0,and the most suitable light-dark cycle was 12∶12 h.
Falcatariamoluccana;canker disease;Lasiodiplodiatheobromae;biological characteristics
2016-11-17;
2016-12-18
国家林业公益性行业科研专项(No.201304113)
王伟(1992—),男,山西太原人,在读研究生,研究方向为植物病理学,E-mail:565422947@qq.com
纪春艳,博士,从事植物病理学研究,E-mail:jcy3210@163.com。
S763.15
A
1671-0886(2017)06-0014-04
猜你喜欢
中老年保健(2022年7期)2022-09-20 01:07:16
陕西农业科学(2022年2期)2022-03-28 02:41:54
华人时刊(2021年13期)2021-11-27 09:18:54
浙江农业学报(2021年5期)2021-05-31 07:29:08
上海农业学报(2017年4期)2017-04-10 12:40:22
广东饲料(2016年5期)2016-12-01 03:43:21
中国酿造(2016年12期)2016-03-01 03:08:11
华侨华人历史研究(2016年2期)2016-02-27 14:58:37
浙江人大(2014年5期)2014-03-20 16:20:25
中国酿造(2014年9期)2014-03-11 20:21:03
