
水稻分蘖角基因TAC1的育种应用价值分析
2017-12-02 03:19陈宗祥冯志明王龙平冯凡张亚芳马玉银潘学彪左示敏
中国水稻科学 2017年6期
陈宗祥 冯志明 王龙平 冯凡 张亚芳 马玉银 潘学彪 左示敏,*
(1扬州大学 植物功能基因组学教育部重点实验室/江苏省粮食作物现代产业技术协同创新中心,江苏 扬州 225009; 2扬州职业大学,江苏 扬州225000;*通讯联系人,E-mail: smzuo@yzu.edu.cn)
水稻分蘖角基因TAC1的育种应用价值分析
陈宗祥1冯志明1王龙平1冯凡1张亚芳1马玉银2潘学彪1左示敏1,*
(1扬州大学 植物功能基因组学教育部重点实验室/江苏省粮食作物现代产业技术协同创新中心,江苏 扬州 225009;2扬州职业大学,江苏 扬州225000;*通讯联系人,E-mail: smzuo@yzu.edu.cn)
【目的】分蘖角度是水稻重要株型性状,合理的分蘖角度是培育理想株型、达到高产育种的一个关键因素。【方法】以控制水稻分蘖角度增大的显性主效数量基因TAC1为研究对象,构建不同品种背景下的TAC1近等基因系,通过单本栽插、多本栽插/高产栽培以及纹枯病菌接种鉴定,分析TAC1对其他农艺性状、纹枯病抗性、产量及品质的影响。【结果】同一背景近等基因系之间比较,TAC1使水稻品系分蘖角度增加,有利于减轻纹枯病危害,对其他农艺性状无不利影响。在高产栽培条件下,不同背景TAC1系有效穗数均多于tac1系;籼稻特青背景下,TAC1系结实率、千粒重及单株产量均高于tac1系;美国稻Lemont背景下,TAC1系结实率、千粒重及单株产量均低于tac1系;粳稻武陵粳1号、镇稻88背景下,TAC1系结实率和千粒重均略低于tac1系,单株产量略高于tac1系,差异不显著。加工及外观品质方面,特青TAC1系优于tac1系,武陵粳1号、镇稻88以及Lemont背景TAC1系较tac1系有劣化趋势,差异均不显著。【结论】叶片较长的籼稻品种,适宜的分蘖角度范围较窄,叶片较小的粳稻品种,适宜的分蘖角度范围较宽。适当增加水稻品种的分蘖角度,有利于减轻纹枯病危害。TAC1可用于株型紧凑型籼稻品种及粳稻品种的株型改良。
分蘖角度;理想株型;纹枯病;产量
分蘖角度是衡量植株松紧程度的指标之一,是水稻重要株型性状,对水稻的冠层结构、光合效率、物质生产和抗病性等都有重要的影响[1]。分蘖角度尽管受环境影响,但主要还是由遗传因素决定[2]。针对不同材料的研究发现,水稻分蘖角度既有表现为由主基因控制的质量性状,也有表现为由多基因控制的数量性状。作为质量性状,最典型的是Takahashi等[3-4]发现的控制水稻分蘖平卧生长的la基因和直立生长的er基因;Kinoshita等[5]报道了具有多效性的畸形矮秆基因d20也能使植株株型分散。李培金等[6]通过对水稻散生突变体la(k)进行遗传分析发现,该散生表型受一隐性核基因控制,与已报道的水稻散生基因la等位,将此突变体命名为la-2,而将la重新命名为la-1,在对la-1精细定位在的基础上,图位克隆了lazy1基因。Tan等[7]克隆了控制野生稻匍匐生长习性的基因 PROG1(PROSTRATE GROWTH1)。Yamamoto等[8]对籼粳回交后代进行分析,认为源于籼稻Kasalath的显性单基因Spk(t)控制株型,增加了水稻的分蘖角度,使水稻呈散生性状。对人工诱变产生的散生突变体研究也表明,tag1[9]和tac2[10]的散生性状都由1对隐性主效基因控制。作为数量性状的研究报道也较多,Li等[11]、Yan 等[12]、钱前等[13]、沈圣泉等[14]、余传元等[15]、赵春芳等[16]利用不同的遗传群体,在不同染色体上定位了多个分蘖角度 QTL。Dong等[2]对 529份亚洲栽培稻种质资源进行全基因组测序得到大量的SNP分子标记,并依次对这些材料开花期的分蘖角度进行全基因组关联分析(genome wide association study,GWAS),共检测到30个控制栽培稻分蘖角度的QTL,并通过突变体表型与基因型的共分离鉴定,成功分离了位于水稻第3染色体上qTA3的候选基因TAC3。多个研究小组都在水稻第9染色体长臂上定位到了控制分蘖角度的主效QTL。Yu等[17]利用一个株型松散的籼稻品种 IR24为遗传背景、渗入了少量粳稻品种Asominori染色体片段的株型紧凑渗入系 IL55为材料,通过图位克隆分离到了该主效QTL,命名为TAC1。TAC1是控制水稻分蘖角度增加的主效数量基因,具有显性效应,是目前少数几个被克隆的控制分蘖角度的主效QTL。
合理的分蘖角度是培育理想株型、达到高产育种的一个关键因素[18]。高产水稻群体要求株型松散适中,前人定位的控制水稻散生和直立两种极端类型的分蘖角度基因基本没有实际育种利用价值,而那些具有一定效应的主效数量基因在株型改良中可能具有重要价值。我们在进行抗纹枯病分子育种过程中,获得不同遗传背景下的分蘖角主效QTL TAC1的近等基因系[19]。本研究利用这些近等基因系分析不同分蘖角度对水稻产量及品质性状的影响,为进一步合理利用TAC1提供参考。
1 材料与方法
1.1试验材料
试验材料包括两部分,一部分是本课题组通过分子标记辅助选择获得的同一背景下携带TAC1基因或tac1基因的4组近等基因系,依据品种的遗传背景分别命名为特青-TAC1和特青-tac1,Lemont-TAC1和 Lemont-tac1,武陵粳 1号-TAC1和武陵粳1号-tac1,镇稻88-TAC1和镇稻88-tac1;另一部分材料由中国农业大学孙传清教授提供,包括籼稻品种 IR24(携带 TAC1,命名为 IR24-TAC1)及其遗传背景下的 TAC1基因 RNA干扰系(IR24-TAC1Ri)、日本晴(携带tac1,日本晴-tac1)及其背景下的TAC1基因超表达系(日本晴-TAC1ox)。
1.2试验方法
1.2.1 田间试验设计
试验在扬州大学校内试验田进行。试验材料 5月上旬播种,6月上旬移栽。设产量试验圃和纹枯病抗性鉴定圃。产量试验部分,每材料栽8行,每行50株,株行距13.3 cm×25 cm,单本栽插,随机区组设计,3次重复,常规肥水管理。纹枯病抗性鉴定部分,每材料栽3行,每行12株,株行距13.3 cm×25 cm,单本栽插,随机区组设计,区组内各实验材料小区采用完全随机区组排列,3次重复,偏施氮肥,田间保持水层以利于发病。
在单本栽插试验基础上,选择近等基因系间农艺性状更接近的4组材料,进行多本栽插产量比较试验。每材料栽8行,每行50穴,穴行距13.3 cm×25 cm,每穴2~3苗,随机区组设计,3次重复。按高产栽培要求进行肥水管理和病虫害防治,施总氮量(折合纯 N)为 285 kg/hm2,氮(以纯 N 计)、磷(折合P2O5)、钾肥(折合K2O)施用比例为10∶3∶5。其中氮肥按m基肥∶m分蘖肥∶m穗肥=4∶3∶3比例施用,磷肥全部基施;钾肥50%作基肥,50%于拔节期施用。肥料以东溪牌45%复合肥为主,配以中煤牌46.4%大颗粒尿素和俄罗斯进口的硫酸钾肥。
1.2.2 性状调查
在单本栽插产量试验圃,移栽活棵后定点调查基本苗,以后每隔5 d调查各试验小区茎蘖数,每点10株,10株平均值代表该小区茎蘖数,至茎蘖数明显下降为止,以“100×(高峰苗-基本苗)/基本苗”计算分蘖率。记载各小区抽穗期,齐穗时用量角器测量各小区的分蘖角度(1株中开张度最大的两个茎秆之间的夹角)[20],收获前测量各试验材料株高,每区均测量10株,以10株的平均值代表该小区性状值。
收获前先普查有效穗数,即连续计数200株有效穗数,计算单株平均有效穗,选择有效穗数与普查有效穗相当的单株 20株,混合收获、脱粒,考查产量性状,项目包括有效穗数、每穗粒数、结实率、千粒重、单株产量等。以“100×有效穗/高峰苗”计算成穗率。
多本栽插产量试验圃,茎蘖动态、产量及其构成因素考查方法同上。晒干扬净的种子按照《GB/T 17891-1999》标准进行加工品质和外观品质分析。
1.2.3 纹枯病菌接种及病情调查
纹枯病抗性鉴定圃,采用牙签嵌入法接种纹枯病病菌[21],接种于7月16日进行,每个小区接种中间1行的中间10株,每株接种3个茎秆,两边各1行和两端各1株作为保护行。记载各小区抽穗期,抽穗后 30 d左右采用左示敏等[22]改进的 0~9级病级评价标准调查纹枯病病级。
1.2.4 数据分析
应用Excel 2010软件整理数据,DPS 14.5软件对数据进行分析处理。
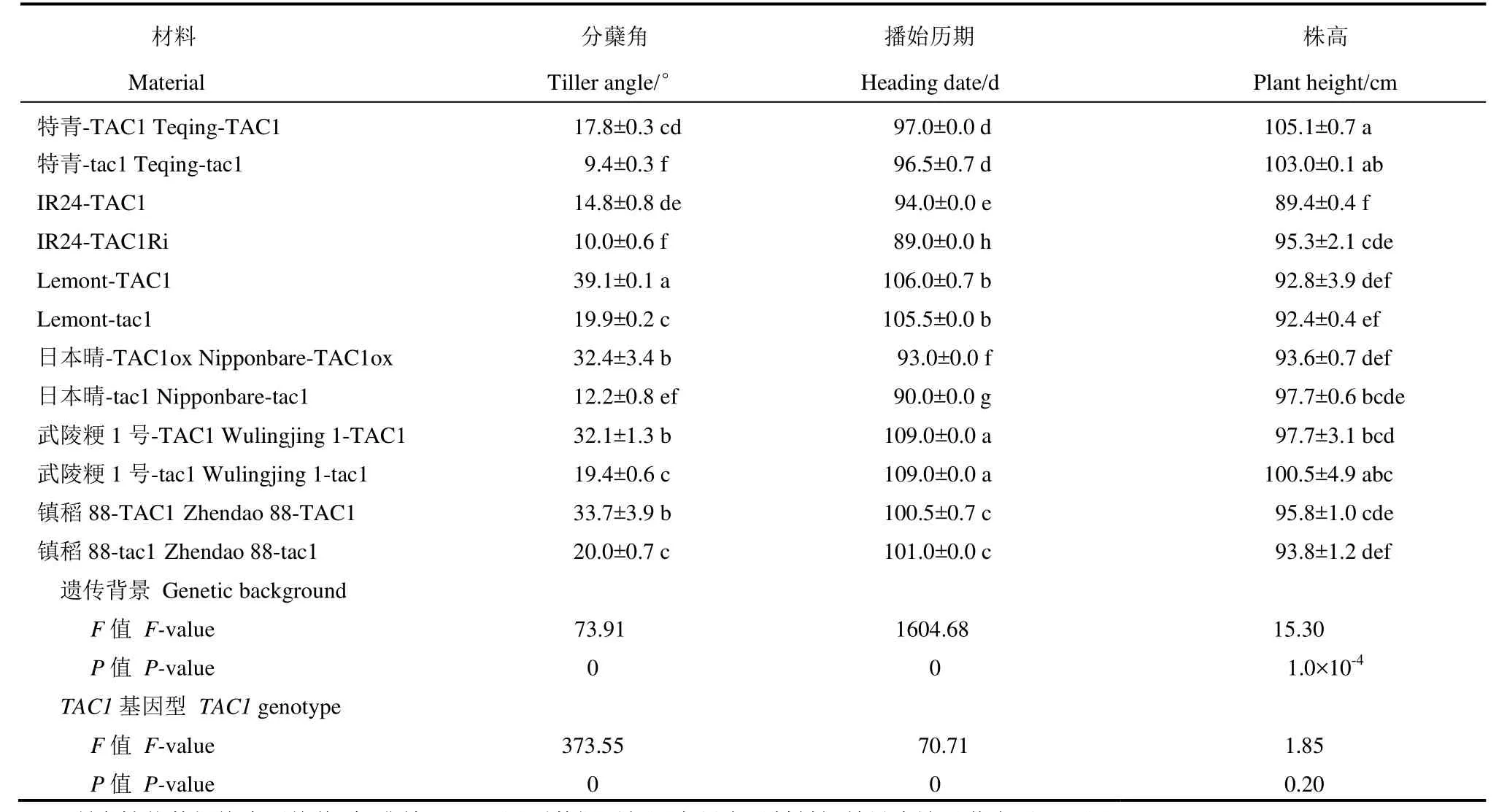
表1 供试水稻材料的基本农艺性状Table 1. Information of the basic agronomic traits of the rice lines tested in the study.
2 结果与分析
2.1试验材料基本性状
按照遗传背景、TAC1基因型二因素试验统计分析,所调查的3个性状,遗传背景间均存在极显著差异(表1)。分蘖角度性状基因型间差异达显著水平,同一背景的两个系比较,带有TAC1的品系其分蘖角度均显著大于带有tac1的系,说明TAC1基因效应在本试验材料中得以表达;不同背景比较,特青-TAC1的分蘖角度与武陵粳 1号-tac1、镇稻88-tac1以及Lemont-tac1基本一致,显著小于武陵粳1号- TAC1、镇稻88-TAC1以及Lemont-TAC1的分蘖角度,大于特青-tac1的分蘖角度,表明TAC1的表达还受遗传背景影响。生育期性状在不同分蘖角度材料间的差异也达显著水平,以 IR24和日本晴为背景的2组材料,大角度品系的播始历期显著长于小角度品系,另外4组材料的生育期在大小角度品系间的差异则不显著。株高性状在不同分蘖角度材料间的差异不显著。通过分子标记辅助选择获得的4对近等基因系,两两之间株高和生育期性状差异较小,说明TAC1的表达对这两个性状基本没有影响;IR24和日本晴背景分蘖角度有差异的两个系之间,株高和生育期也有较大差异,这可能是相关转基因系中存在组培变异或其他未知因素所致。

表2 供试水稻材料纹枯病病级方差分析表Table 2. ANOVA of sheath blight scores of the rice lines tested in the study.

表3 供试水稻材料纹枯病病级多重比较Table 3. Multiple comparison of sheath blight scores of the rice lines tested in the study.
2.2分蘖角基因TAC1对纹枯病抗性的影响
按照遗传背景、分蘖角基因型二因素试验统计分析结果表明,遗传背景之间以及分蘖角基因型之间纹枯病病级差异均达极显著水平(表2)。同一遗传背景两个系比较(表3),TAC1系的纹枯病病级均小于tac1系,其中以特青、Lemont、武陵粳1号为背景的两个系之间纹枯病病级差异达显著水平,说明TAC1基因可使分蘖角度增大,并因此减轻水稻纹枯病的危害度。
2.3分蘖角基因TAC1对群体结构的影响
采用随机区组设计二因素试验统计分析方法对供试材料部分群体构成因素进行统计分析。结果显示(表4),单本栽插条件下,遗传背景之间基本苗、高峰苗、分蘖率和成穗率的差异均达极显著水平,这种差异反映遗传背景的基本特性,如 Lemont分蘖能力较弱,而IR24、武陵粳1号、镇稻88分蘖能力较强。同一背景下的不同分蘖角度基因型材料间所调查的4个性状差异均不显著,携带TAC1基因的材料分蘖率总体略高于携带tac1的基因品系,不同遗传背景间表现相同趋势。
多本栽插条件下,遗传背景之间基本苗、高峰苗、分蘖率和成穗率的差异也均达极显著水平(表5)。同一遗传背景下的不同分蘖角基因型材料之间,高峰苗差异达显著水平,基本苗、分蘖率和成穗率的差异不显著;同一遗传背景下的不同分蘖角度品系中,TAC1系的高峰苗总体高于 tac1系,TAC1系的分蘖率、成穗率均略高于tac1系,不同遗传背景间的表现趋势相同。以上结果表明分蘖角基因TAC1有利于分蘖发生和成穗。
2.4分蘖角基因TAC1对经济性状的影响
单本栽插条件下,遗传背景之间单株产量及其构成因素的差异均达极显著差异(表6),这种差异反映了遗传背景的基本特性,如以武陵粳1号、镇稻88为遗传背景的材料产量较高,而以 Lemont、日本晴为遗传背景的材料产量较低。同一遗传背景下,不同分蘖角基因型材料之间在单株产量及其构成因素上均差异不显著,表明近等基因系之间除了目标性状有差异外,其他产量及其构成因素表现基本一致。

表4 单本栽插各试验材料群体构成Table 4. Population characteristics of rice lines tested in one-plant one-point transplantation condition.
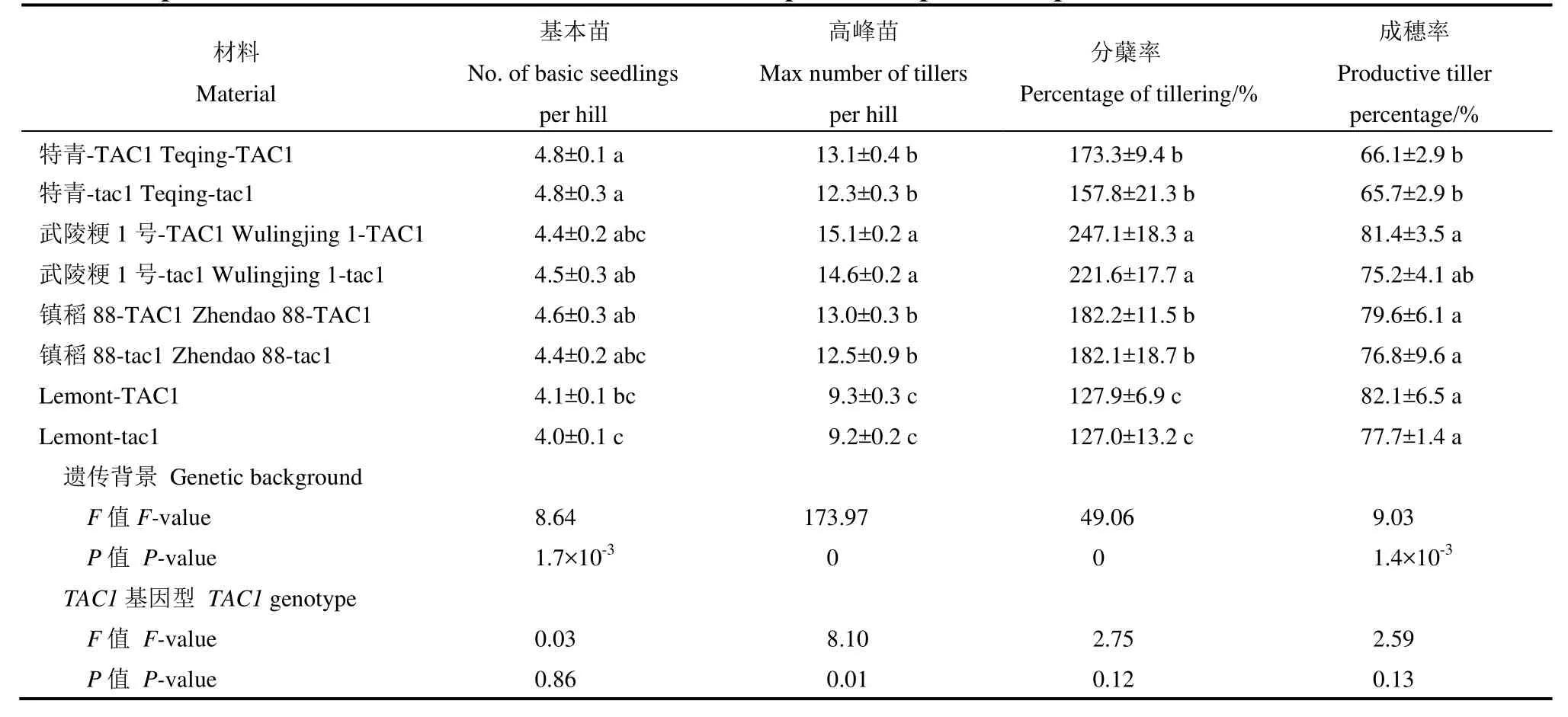
表5 多本栽插各供试材料群体构成Table 5. Population characteristics of rice lines tested in multi-plants one-point transplantation condition.
多本栽插的高产栽培条件下,遗传背景之间单株产量及其构成因素也均存在极显著差异(表7)。不同分蘖角基因型材料之间有效穗数、结实率、千粒重差异达显著至极显著水平。同一背景中的不同角度品系间比较,TAC1系的有效穗数总体多于tac1系,不同背景间的表现区试相同;除特青背景外,其余3组背景材料中,tac1系的结实率和千粒重总体均高于TAC1系。总体而言,分蘖角基因型材料之间在每穗粒数和单株产量上的差异均不显著,但就不同背景而言,特青、武陵粳1号和镇稻88三个背景的TAC1系单株产量均高于tac1系,其中特青背景中两个系之间的产量差异达显著水平,Lemont背景中的单株产量 TAC1系显著小于 tac1系。

表6 单本栽插各供试水稻材料的经济性状Table 6. Multiple comparison of plant yield and its components among rice lines tested in one-plant one-point transplantation condition.

表7 多本栽插各供试水稻材料的经济性状Table 7. Multiple comparison of plant yield and its components among rice lines tested in multi-plants one-point transplantation condition.

表8 多本栽插各供试水稻材料加工及外观品质Table 8. Multiple comparison of grain processing and appearance qualities among rice lines tested in multi-plants one-point transplantation condition.
2.5分蘖角基因TAC1对加工及外观品质性状的影响
多本栽插高产栽培条件下,不同遗传背景材料之间在出糙率、整精米率、垩白率和垩白度上的差异均达显著至极显著水平(表8),而不同分蘖角基因型材料之间在这些性状上均差异不显著,表明在本研究条件下,分蘖角基因TAC1对稻米加工及外观品质影响较小。
3 讨论
钱前等[13]认为,水稻分蘖角度与产量呈负相关,即分蘖角度过大,影响群体产量构成,使产量降低。梁彦等[23]认为,合适的分蘖角度还有利于促进水稻的光合作用,提高抗倒伏能力,进而提高水稻的单株产量和收获指数;同时,分蘖角度直接决定水稻的种植密度,对田间群体产量的影响更大。可见,合理的分蘖角度对提高产量有重要意义,然而,多大的分蘖角度为合理,并没有明确的结论。一方面前人的研究多以自然品种为对象,这些材料不仅分蘖角度有差异,其他性状也各不相同,这些各不相同的性状对分蘖角的效应难免产生影响;另一方面,不同的研究者对分蘖角的定义不同、测量时期不一样[1,24],使得不同的研究成果间缺乏可比性。本研究采用的材料为几个不同遗传背景下的分蘖角基因TAC1近等基因系,在单本栽插条件下,同一背景近等基因系之间除了分蘖角度有差异外,其他性状基本一致,一方面表明分蘖角基因TAC1对水稻其他农艺性状无不利影响,另一方面也排除了其他农艺性状对分蘖角效应评价结果的干扰。
本研究表明TAC1使分蘖角度增大,有利于减轻纹枯病危害,与之前的研究结果一致[19]。不同背景下携带TAC1基因系与携带tac1基因系之间的纹枯病病级差异不尽相同,以特青背景下的两者差异最大,达到1.12级,其他背景下的近等基因系间病级差异变幅为 0.26~0.53级。至于为何特青-TAC1的纹枯病病级明显小于特青-tac1,这应该是由于特青-TAC1携带的TAC1基因连锁片段上还包含一个抗纹枯病主效QTL qSB-9TQ。我们在构建不同背景qSB-9TQ近等基因系时,利用覆盖目标区间的 3个分子标记辅助选择,随着对qSB-9TQ进一步定位,发现在原定位区间还存在一个控制分蘖角度的基因 TAC1,并应用精细定位获得的新的分子标记分别对qSB-9TQ和TAC1进行了选择,武陵粳1号、镇稻88以及Lemont背景下,最终实现了qSB-9TQ和TAC1的分离,获得各带一个QTL的近等基因系,而特青背景下,qSB-9TQ和TAC1未能分开,导致特青-TAC1中同时还携带了抗纹枯病等位基因qSB-9TQ,而特青-tac1同时携带的是感病等位基因qsb-9tq[19],因此,特青-TAC1对纹枯病的抗性表现其实是qSB-9TQ和TAC1共同作用的结果。
在多本栽插、高产栽培条件下,TAC1系的高峰苗、分蘖率和成穗率均高于tac1系(表5),最终TAC1系的有效穗多于tac1系(表7),4个背景表现相同的趋势。近等基因系之间在结实率、千粒重及单株产量等方面的差异不同背景趋势并不相同,特青-TAC1比特青-tac1有优势,Lemont-tac1比Lemont-TAC1有优势,武陵粳1号及镇稻88两个背景近等基因系之间差异不明显,结合遗传背景(品种)和近等基因系的分蘖角度分析,更容易理解这一现象。特青和 Lemont叶片较大,农艺性状表现更优的特青-TAC1和Lemont-tac1的分蘖角度分别为17.8°和19.9°,分蘖角度过小(特青-tac1分蘖角度为9.4°),株型紧束导致株内郁蔽,分蘖角度过大(Lemont-TAC1的分蘖角度为39.1°),株型过于松散导致株间遮蔽严重,都使得结实率降低、千粒重下降最终产量降低。而粳稻品种武陵粳1号和镇稻88叶片相对较小,分蘖角度从19.4°增加至33.7°时,株间遮蔽影响相对较小,尽管结实率和千粒重有降低的趋势,但由于有效穗数增加,最终TAC1系的产量还略大于tac1系,但差异不明显。加工及外观品质方面,特青背景分蘖角变小以及Lemont、武陵粳1号和镇稻88背景的分蘖角增大,都有使品质劣化的趋势,但差异均不显著。
综合分蘖角度对产量、抗性及品种等方面的影响分析,籼稻等叶片较大的品种,相对于株型过于紧束(特青-tac1的分蘖角度为9.4°)和株型过于松散(Lemont-TAC1的分蘖角度为39.1°)的品系,分蘖角度在17.8°~19.9°比较适宜。而粳稻品种分蘖角度在19.4°~33.7°之间都比较合理,且在此范围内,分蘖角度增大,更有利于减轻纹枯病危害,增加有效穗数,进而提高产量。实际上,本研究利用的武陵粳1号和镇稻88两个品种,在粳稻中属于分蘖角度较大的一类,有相当数量粳稻品种,分蘖角度在10.0°~18.0°,这些品种尽管株型紧凑,但都有较高的产量潜力。由此说明,对于叶片较小的粳稻品种,适宜的分蘖角度范围可能较宽。
Yu等[17]研究认为,松散型籼稻品种都带有TAC1基因,而tac1基因在粳稻中广泛分布。本研究表明,TAC1系与tac1系比较,分蘖角度仅增加8.4°~13.7°,因此,将TAC1导入株型紧凑的籼稻品种或粳稻品种,适当增加水稻品种的分蘖角度,既可以提高其纹枯病抗性水平,又能保持产量水平不变甚至增产,具有较高的育种应用价值。当然,分蘖角度不同的品种,尤其是分蘖角度差异较大的品种,其适宜的栽培措施也不同,甚至会有较大差异,有必要根据不同的品种类型研究更科学的配套栽培措施,以充分发挥其各自优势。
[1] 徐云碧,申宗坦. 早籼稻品种分蘖角度的遗传分析. 浙江农业学报,1993,5(1): 1-5.Xu Y B,Shen Z T. Genetic analysis of tiller angles for early season indica rice. Acta Agric,1993,5(1): 1-5. (in Chinese with English abstract)
[2] Dong H J,Zhao H,Xie W B,Han Z M,Li G W,Yao W,Bai X F,Hu Y,Guo Z L,Lu K,Yang L,Xing Y Z. A novel tiller angle gene,TAC3,together with TAC1 and D2 largely determine the natural variation of tiller angle in rice cultivars. PLoS Genet,2016. https://doi.org/10.1371/journal.pgen.1006412.
[3] Takahashi M. Linkage groups and gene schemes of some striking morphological characters in Japanese rice//IRRI.Rice Genetics and Cytogenetics. Amsterdam: Elsevier,1964: 215-236.
[4] Takahashi M,Kinoshita T,Takeda K. Character expression and caudal genes of some mutants in rice. Fac Agric Hokkaido Univ,1968,54: 496-512.
[5] Kinoshita T,Takahashi M,Mori K. Character expression and inheritance node of three kinds of dwarf rice. Res Bull Farm Hokkaido Univ,1974,19: 64-75.
[6] 李培金,曾大力,刘新仿,徐聃,谷岱,李家洋,钱前.水稻散生突变体的遗传和基因定位研究. 科学通报,2003,48(21): 2271-2274.Li P J,Zeng D L,Liu X F,Xu D,Gu D,Li J Y,Qian Q.Genetic analysis and gene mapping of rice tiller angle mutant. Sci Bull,2003,48(21): 2271-2274. (in Chinese with English abstract)
[7] Tan L,Li X,Liu F,Sun X,Li C,Zhu Z,Fu Y,Cai H,Wang X,Xie D,Sun C. Control of a key transition from prostrate to erect growth in rice domestication. Nat Genet,2008,40(11): 1360-1364.
[8] Yamamoto T,Sasaki T,Yano M. Genetic analysis of spreading stub using indica/japonica backcrossed progenies in rice. Breeding Sci,1997,47(2): 141-144.
[9] 杨菲,张德春. 一个水稻散生突变体tag1的遗传分析及基因定位. 农业现代化研究,2015,36(4): 684-689.Yang F,Zhang D C. Genetic analysis and fine-mapping of a tiller spreading mutant in rice. Res Agric Modern,2015,36(4): 684-689. (in Chinese with English abstract)
[10] 方立魁,桑贤春,杨正林,凌英华,王楠,何光华. 一个水稻分蘖角度突变体tac2的遗传分析和基因初步定位.中国水稻科学,2009,23(3): 315-318.Fang L K,Sang X C,Yang Z L,Ling Y H,Wang N,He G H. Genetic analysis and gene mapping of a rice tiller angle mutant tac2. Chin J Rice Sci,2009,23(3): 315-318. (in Chinese with English abstract)
[11] Li Z K,Paterson A H,Pinson S R M,Stansel J W. RFLP facilitated analysis of tiller and leaf angles in rice.Euphytica,1999,109(2): 79-84.
[12] Yan J Q,Zhu J,He C X,Benmoussa M,Wu P.Quantitative trait loci analysis for the developmental behavior of tiller number in rice (Oryza sativa L.). Theor Appl Genet,1998,97(1/2): 267-274.
[13] 钱前,何平,滕胜,曾大力,朱立煌. 水稻分蘖角度的QTLs分析. 遗传学报,2001,28(1): 29-32.Qian Q,He P,Teng S,Zeng D L,Zhu L H. QTLs analysis of tiller angle in rice (Oryza sativa L.). Acta Genet Sin,2001,28(1): 29-32. (in Chinese with English abstract)
[14] 沈圣泉,庄杰云,包劲松,郑康乐,夏英武,舒庆尧. 水稻分蘖最大角度的QTL分析. 农业生物技术学报,2005,13(1): 16-20.Sheng S Q,Zhuang J Y,Bao J S,Zheng K L,Xia Y W,Shu Q Y. QTL analysis of rice maximum tiller angle.Agric Biotechnol J,2005,13(1): 16-20. (in Chinese with English abstract)
[15] 余传元,刘裕强,江玲,王春明,翟虎渠,万建民.水稻分蘖角度的QTL定位和主效基因的遗传分析. 遗传学报,2005,32 (9): 948-954.Yu C Y,Liu Y Q,Jiang L,Wang C M,Zhai H Q,Wan J M. QTLs mapping and genetic analysis of tiller angle in rice (Oryza sativa L.). Acta Genet Sin,2005,32(9):948-954. (in Chinese with English abstract)
[16] 赵春芳,周丽慧,于新,赵庆勇,陈涛,姚姝,张亚东,朱镇,王才林. 基于CSSL的高密度物理图谱定位水稻分蘖角度QTL. 植物学报,2012,47(6): 594-601.Zhao C F,Zhou L H,Yu X,Zhao Q Y,Chen T,Yao S,Zhang Y D,Zhu Z,Wang C L. Identification of tiller angle quantitative trait loci based on chromosome segment substituted lines and high-density physical map in rice.Chin Bull Bot,2012,47(6): 594-601. (in Chinese with English abstract)
[17] Yu B S,Lin Z W,Li H X,Li X J,Li J Y,Wang YH,Zhang X,Zhu Z F,Zhai W X,Wang X K,Xie D X,Sun C Q. TAC1,a major quantitative trait locus controlling tiller angle in rice. Plant J,2007,52(5): 891-898.
[18] 李家洋. 水稻分蘖数目与分蘖角度的分子机理. 中国基础科学,2008,10(3): 14-15.Li J Y. Molecular regulation mechanism of rice tillering number and tillering angle. China Basic Sci,2008,10(3):14-15. (in Chinese with English abstract)
[19] 陈宗祥,左示敏,张亚芳,陈红旗,冯明辉,姜伟,冯凡,马玉银,华鹤良,李国生,潘学彪. 抗纹枯病基因qSB-9TQ和分蘖角基因TAC1TQ在抗病育种中的互作效应及育种价值. 中国水稻科学,2014,28(5): 479-486.Chen Z X,Zuo S M,Zhang Y F,Chen H Q,Feng M H,Jiang W,Feng F,Ma Y Y,Hua H L,Li G S,Pan X B.Breeding potential and interaction effects of qSB-9TQand TAC1TQin rice breeding against sheath blight disease.Chin J Rice Sci,2014,28(5): 479-486. (in Chinese with English abstract)
[20] Han Y P,Xing Y Z,Gu S L,Chen Z X,Pan X B,Chen X L. Effect of morphological traits on sheath blight resistance in rice. Acta Bot Sin,2003,45(7): 825-831.
[21] 潘学彪,陈宗祥,徐敬友,童蕴慧,王子斌,潘兴元.不同接种调查方法对抗水稻纹枯病遗传研究的影响. 江苏农学院学报,1997,18(3): 27-32.Pan X B,Chen Z X,Xu J Y,Tong Y H,Wang Z B,Pan X Y. The effects of different methods of inoculation and investigation on genetic research of resistance to rice sheath blight. J Jiangsu Agric Coll,1997,18(3): 27-32. (in Chinese with English abstract)
[22] 左示敏,张亚芳,殷跃军,陈宗祥,潘学彪. 田间水稻纹枯病抗性鉴定体系的确立与完善. 扬州大学学报: 农业与生命科学版,2006,27(4): 57-61.Zuo S M,Zhang Y F,Yin Y J,Chen Z X,Pan X B.Establishment and improvement of inoculation technique and rating system in researching rice sheath blight resistance in field. J Yangzhou Univ: Agric amp; Life Sci,2006,27(4): 57-61. (in Chinese with English abstract)
[23] 梁彦,王永红. 水稻株型功能基因及其在育种上的应用.生命科学,2016,28(10): 1156-1167.Liang Y,Wang Y H. The genes controlling rice architecture and its application in breeding. Chin Bull Life Sci,2016,28(10): 1156-1167. (in Chinese with English abstract)
[24] 谢元璋,夏仲炎. 粳稻品种分蘖性状的遗传研究. 安徽农业科学,1994,22(4): 319-322.Xie Y Z,Xia Z Y. Genetic analysis of japonica rice tillering. Anhui Agric Sci,1994,22(4): 319-322. (in Chinese with English abstract)
Breeding Potential of Rice TAC1 Gene for Tiller Angle
CHEN Zongxiang1,FENG Zhiming1,WANG Longping1,FENG Fan1,ZHANG Yafang1,MA Yuyin2,PAN Xuebiao1,ZUO Shimin1,*
(1Key Laboratory of Plant Functional Genomics,Ministry of Education / Co-Innovation Center for Modern Production Technology of Grain Crops,Yangzhou University,Yangzhou 225009,China; 2 Yangzhou Polytechnic College,Yangzhou 225000,China; *Corresponding author,E-mail: smzuo @yzu.edu.cn)
【Objective】Rice tiller angle is an important plant-type trait. The appropriate tiller angle is generally a critical factor in developing ideal-type high-yielding rice variety. 【Methods】Effects of tiller angle controlling gene TAC1 on agronomic traits,sheath blight resistance,grain yield and quality were investigated under conditions of one-plant one-point and multi-plants one-point transplantation by using four pairs of near isogenic lines(NILs) at TAC1 gene.【Results】TAC1 increased rice tiller angle in all the genetic backgrounds tested,which is beneficial for reducing adverse impacts of sheath blight,without affecting other agronomical traits. Under high-yielding cultivation condition of multi-plant one-point transplantation,numbers of effective panicles in all the NILs with TAC1 were apparently higher than those of corresponding NILs with tac1. In the background of indica rice Teqing,the seed-setting rate,1000-grain weight and plant yield of Teqing-TAC1 line were all higher than those of Teqing-tac1 line; however,the reverse results were obtained in the background of Lemont. In backgrounds of Wulingjing 1 and Zhendao 88,NILs with TAC1 gene displayed slightly grain yield than those with tac1 gene. The grain processing and appearance qualities of Teqing-TAC1 line were slightly improved compared to the corresponding control,while other TAC1-harboring NILs showed just the reverse with statistically insignificant difference. 【Conclusions】The indica rice with long leaf has a narrow range of appropriate tiller angles but for japonica rice with short leaf,the range is wider. Increasing rice tiller angle to an extent is beneficial for reducing sheath blight influence. TAC1 improves the plant type of japonica rice as well as the compact type indica rice.
tiller angle; ideal-type plant; sheath blight; grain yield
Q754; S511.032
A
1001-7216(2017)06-0590-09
10.16819/j.1001-7216.2017.7100
2017-08-20; 修改稿收到日期:2017-10-11。
国家自然科学基金资助项目(31571748); 扬州市重点研发计划(现代农业)资助项目(YZ2015028); 江苏省自然科学基金资助项目(BK20171293); 江苏省农业重点研发计划资助项目(BE2015341)。
猜你喜欢
区域治理(2022年40期)2022-11-27
科学与财富(2021年33期)2021-05-10
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
农民致富之友(2019年21期)2019-08-08
安徽农业科学(2018年30期)2018-11-02
农家科技中旬版(2018年2期)2018-05-05
新农业(2017年6期)2017-07-15
