
水稻小粒基因SG101的鉴定和精细定位
2017-12-02 03:19:03汪玉琼杨窑龙冷语佳黄李超陈龙代丽萍涂政军高易宏胡江朱丽张光恒任德勇高振宇董国军陈光郭龙彪叶国友钱前曾大力
中国水稻科学 2017年6期
汪玉琼 杨窑龙 冷语佳 黄李超 陈龙 代丽萍 涂政军 高易宏 胡江 朱丽张光恒 任德勇 高振宇 董国军 陈光 郭龙彪 叶国友 钱前,* 曾大力,*
(1中国水稻研究所 水稻生物学国家重点实验室,杭州 310006; 2国际水稻研究所,菲律宾 马尼拉;#共同第一作者; *通讯联系人,E-mail:dalizeng@126.com; qianqian188@hotmail.com)
水稻小粒基因SG101的鉴定和精细定位
汪玉琼1,#杨窑龙1,#冷语佳1黄李超1陈龙1代丽萍1涂政军1高易宏1胡江1朱丽1张光恒1任德勇1高振宇1董国军1陈光1郭龙彪1叶国友2钱前1,*曾大力1,*
(1中国水稻研究所 水稻生物学国家重点实验室,杭州 310006;2国际水稻研究所,菲律宾 马尼拉;#共同第一作者;*通讯联系人,E-mail:dalizeng@126.com; qianqian188@hotmail.com)
【目的】籽粒大小是决定水稻产量的重要农艺性状之一,开展水稻籽粒大小相关基因的克隆和功能研究对于阐述水稻产量形成的遗传调控机制具有重要意义。【方法】利用甲基磺酸乙酯诱变粳稻品种中花11,筛选获得一小粒突变体,命名为sg101(small grain 101)。通过形态学、细胞学手段调查了SG101的突变对籽粒大小、穗部主要性状及颖壳细胞数目和大小的影响,通过测定叶夹角和胚芽鞘长度分析其对外施油菜素内酯的差异响应,结合定量 PCR技术分析了油菜素内酯合成途径和信号途径相关基因表达情况,并利用图位克隆的手段精细定位了水稻小粒基因SG101。【结果】与野生型相比,突变体sg101粒长和粒宽均极显著减小,从而导致千粒重极显著降低。此外,sg101还表现出结实率降低、穗长变短、二次枝梗数减少、植株变矮等。细胞学观察发现 sg101的颖壳细胞大小没有改变,但细胞数目明显减少。定量PCR检测表明sg101中的细胞周期相关基因表达显著下降。另外,突变体 sg101对外施油菜素内酯响应迟钝,其油菜素内酯合成途径和信号途径相关基因表达亦显著降低。【结论】遗传分析表明sg101突变体由隐性单基因控制,通过图位克隆的方法将SG101精细定位于第1染色体上,物理距离为265 ksb的区间内。这为该基因的克隆及深入的功能研究奠定了基础。
水稻;小粒;sg101;油菜素内酯
水稻是重要的粮食作物之一,世界一半以上的人口都以稻米作为主食。近年来,随着人口快速增长、气候变化以及日益严重的土壤污染,作物的有效种植面积逐年减少。因此,提高水稻产量成为保证粮食安全的重要议题。水稻产量主要由每株有效穗数、每穗粒数和千粒重共同决定,其中,粒重的大小取决于粒长、粒宽和粒厚的变化。因此,合理优化籽粒形状,可为培育高产品种提供重要保证[1]。
已有研究表明,水稻粒型大小的遗传调控途径复杂多样,可能涉及泛素化蛋白降解、植物激素和G蛋白信号途径以及不同途径之间的相互作用等。如最先克隆的粒长基因GS3编码由4个结构域组成的跨膜蛋白;长粒品种明恢63由于OSR结构域发生碱基突变造成蛋白翻译提前终止;野生型日本晴包含完整的4个结构域而表现为中间型的籽粒;小粒品种Chuan 7由于在第5外显子发生碱基突变,造成移码而导致羧基端两个结构域缺失[2-6]。GL3.1则为编码一丝/苏蛋白磷酸酶基因,由于 4个 SNP位点的差异导致籽粒长度的改变[7-8]。美国的长粒粳稻品种的GL7位点发生了17.1 kb的DNA大片段串联重复序列,这种基因组结构变异导致基因表达量的上升而引起籽粒长度增加[9]。Hu等[10]鉴定了一个编码水稻生长调控因子OsGRF4的基因GS2,因在miR396c靶点发生一个稀有显性突变造成基因表达量的上升,促进了细胞的分裂和生长而表现粒长增加。GW2编码细胞质E3泛素连接酶,可能参与降解促进细胞分裂的蛋白而调控籽粒大小[11-12]。GW5/qSW5表现与多聚有相互作用,通过泛素蛋白酶体途径调控籽粒宽度[13]。GS5通过上调细胞周期基因的表达、促进细胞分裂而调控籽粒的大小[14],而GW8编码启动子结合蛋白,高表达的GW8促进细胞分裂、增加粒宽[15]。
水稻粒型基因中除了控制籽粒大小的主效QTL之外,还有许多与生长发育和激素传导相关的基因也影响籽粒大小。如 D61、D11、SMG1、DSG1、BG1-D[16-20]也影响籽粒大小、株高及叶片形态等。本研究通过EMS诱变粳稻品种中花11,在田间发现了一粒型变小且能稳定遗传的小粒突变体,并命名为sg101 (small grain 101)。从表型、生理、遗传等方面对该突变体进行鉴定,明确了其遗传特性,并对该突变基因进行了精细定位。通过对突变体的研究,有利于进一步了解水稻籽粒发育的调控机制,为水稻传统育种提供理论基础。
1 材料与方法
1.1 材料
利用EMS诱变粳稻品种中花11,从中筛选到一个小粒突变体,经多代连续自交,获得突变表型稳定的小粒突变体,命名为sg101(small grain 101)。遗传分析是利用突变体分别与台中本地1号(TN1)、南京6号、9311杂交,观察并统计F1植株表型和F1自交形成的F2植株表型。
1.2突变体表型分析
将野生型中花11和突变体sg101在同一时期、相邻小区种植,并设置3个重复;每个小区种植6行,每行6株;待到植株成熟后分别对野生型中花11和突变体sg101的株高、各节间长度、穗长、每穗粒数、一次枝梗数、二次枝梗数、粒长、粒宽、千粒重等农艺性状进行测定。取样时排除边际效应,取中部植株并且测量的均为植株主穗,重复6次。
1.3细胞学观察
1.3.1 石蜡切片观察
取中花11和突变体sg101抽穗后且未开花的颖壳,将其迅速置于FAA固定液 (38%甲醛5 mL,冰醋酸5 mL,70%酒精90 mL) 中,并对固定的材料进行真空抽气,样品经软化、脱水、透明、浸蜡、包埋、切片、染色等,倒置在光学显微镜下(Nikon 90i)拍照。
1.4sg101突变体基因定位
本研究利用混合分离分析法(BSA)构建突变体混池,在定位群体中取带有突变表型的单株 30株左右,分别称取等量叶片混合,利用CTAB法提取DNA[21]。根据实验室已有的163对SSR标记对分离双亲、F1和混池的多态性进行分析,选取突变体与混池DNA电泳条带一致的标记,即与突变基因SG101连锁的标记,再取 93株带有突变表型单株对得到的连锁标记进行验证。将SG101定位于两标记之间,并利用籼稻 9311序列(http://www.rise.genomics.org.cn)和粳稻日本晴(NPB)序列(http://rgp.dna.affrc.go.jp),在目标区间开发多态性标记,进一步进行精细定位。应用Primer Premier 5软件进行引物设计。
1.5 油菜素内酯(BR)处理
1.5.1 叶夹角试验
将中花11和sg101种子脱壳后灭菌,在1/2 MS培养基上发苗,在30℃培养箱里培养10 d,培养条件为16 h光照/8 h黑暗,待小苗长到1叶1心后,剪掉基部,并置于不同浓度梯度(分别是0.00、0.01、0.10、1.00 μmol/L)的油菜素内酯(2,4-epiBL)水溶液中浸泡,28℃下黑暗处理 2 d。处理结束后,用量角器测量叶夹角,并选典型的小苗拍照。
1.5.2胚芽鞘伸长试验
将中花11和sg101种子脱壳后灭菌,在含有不同浓度油菜素内酯(分别是 0.00、0.01、0.10、1.00 μmol/L)的1/2 MS培养基上发芽,30℃培养箱中黑暗培养 7 d,培养结束后,量取胚芽鞘的长度并拍照。
1. 6 RT-PCR
用总RNA提取试剂盒提取突变体sg101和野生型中花11的幼苗的根和茎叶部位总RNA,利用经过DNase Ⅰ处理过的总RNA为模板,使用实时PCR用 cDNA合成试剂盒(TOYOBO)反转录合成cDNA第1链。利用实时定量PCR(RT-PCR)分别检测细胞周期相关基因和油菜素内酯合成途径及信号途径相关基因在野生型和突变体根茎叶中的表达量,以 UBQ5(LOC_Os01g22490)作为内参基因。10 μL实时荧光定量PCR体系如下:cDNA模板1 μL,2× SYBR®Select Master Mix(TOYOBO) 5 μL,正反引物(10 μmol/L)各 0.35 μL,ddH2O 补足 10 μL。实时荧光定量PCR程序如下:95℃下反应10 min,95℃下反应15 s;60℃下反应1 min,72℃下反应15 s,40个循环。用 2-△△Ct法计算各基因的相对表达量。用于检测细胞周期相关基因、油菜素内酯合成途径和信号途径相关基因表达量的引物见表1。

表1 本研究用于RT-PCR分析的引物Table 1. Primers used for real-time RT-PCR in the study.
2 结果与分析
2.1 突变体sg101表型特征和农艺性状分析
与野生型中花11相比,突变体sg101的籽粒显著变小(图1),粒长、粒宽分别只有6.67 mm和5.20 mm,仅为野生型的78%和90%。由于sg101的籽粒在粒长和粒宽上均比野生型显著减小,由此导致sg101在千粒重上也显著降低,结果显示突变体sg101的千粒重只有16.63 g,比野生型中花11的千粒重(25.5 g)下降了35%左右(图1-E)。sg101除了籽粒变小之外,还表现植株变矮,其株高只有野生型的60% (图1-F)。通过进一步比较野生型和突变体sg101各节间长度,发现突变体各节间长度较野生型均发生不同程度缩短,其中第 1节间下降最大,节间长度仅为野生型的60%(图1-G、H)。此外,突变体sg101还表现其他农艺性状的改变,如穗长缩短,一次枝梗数、二次枝梗数和每穗颖花数减少,结实率下降等(表2)。
2.2 突变体sg101籽粒的细胞数目减少
水稻籽粒大小通常由颖壳细胞大小和细胞数目的多少决定。为了明确突变体sg101籽粒变小的原因,我们对完全抽穗但尚未开花的野生型中花11和突变体 sg101的颖壳石蜡切片进行观察(图2-A~C)。通过对颖壳横切面细胞数目统计,发现野生型中花 11的细胞数目可达 1640个,而突变体sg101只有1470个,突变体较野生型减少了10%(图2-D)。另外,我们还通过扫描电镜比较了中花 11和sg101颖壳内表皮的细胞大小(图2-E~F),结果表明其细胞大小没有显著差异(图 2-G)。由此,我们推断sg101突变体籽粒变小是由细胞数目减少引起。
2.3 突变体sg101细胞周期相关基因表达下调
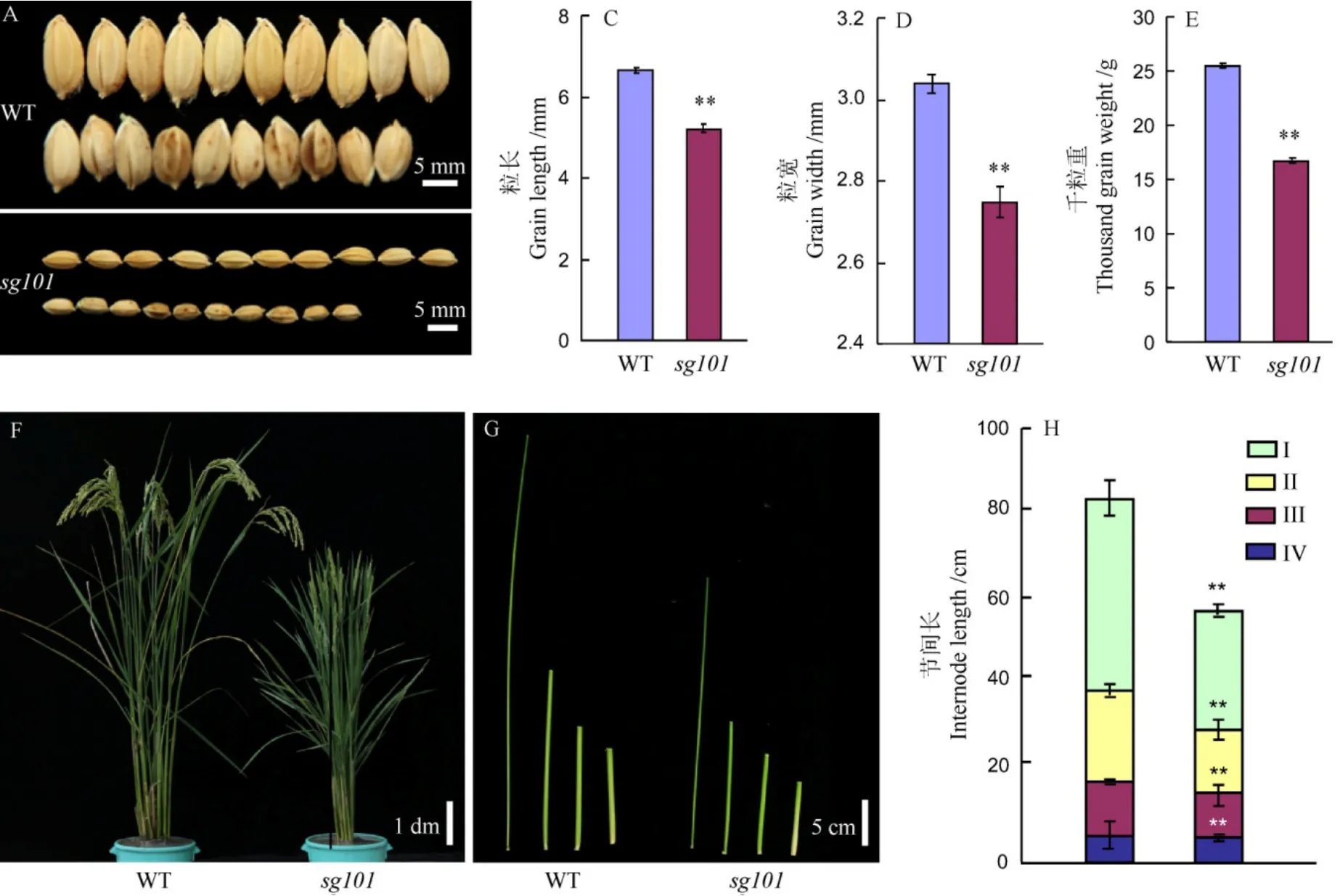
图1 野生型中花11和突变体sg101的表型比较Fig. 1. Phenotypes of Zhonghua 11(WT) and sg101.

表2 野生型中花11和突变体sg101的穗部性状比较Table 2. Comparison of panicle-related traits between Zhonghua 11 and sg101.
细胞数目的变化通常与细胞分裂的改变有关,为了确定突变体sg101细胞数目变少是否与细胞分裂有关,我们利用实时定量PCR技术分析细胞周期相关基因在野生型和突变体中的表达。结果发现,在突变体sg101中,我们所检测的5个细胞周期相关基因的表达均大幅下降(图3)。其中,E2F2的表达量不到野生型的 10%,而 CYCT1:2、CYCD4:1、CYCD7:1和CYCB2:2也只有野生型1/5左右。sg101中细胞周期相关基因表达的显著下调,表明SG101可能通过调控细胞周期的变化而影响籽粒的大小。
2.4sg101对油菜素内酯(BR)处理的响应
sg101植株矮小、叶色深绿、籽粒小而圆且叶夹角变小,呈BR相关突变体的类似表型。为检测sg101是否与油菜素内酯合成或者信号途径有关,我们利用油菜素内酯2,4-epiBL处理野生型中花11和突变体sg10幼苗来测试其对BR的敏感性。由图4-A可以看出,中花11的叶夹角随着2,4-epiBL浓度的增加而增大,当2,4-epiBL的浓度达1 µmol/L时,中花11的叶夹角已明显下垂;而突变体sg101叶夹角的变化始终不明显(图4-B)。另外,我们还进一步比较了外施BR时胚芽鞘的伸长状况。由图4-C发现,2,4-epiBL可以促进野生型中花11和sg101的胚芽鞘生长,中花11对2,4-epiBL更敏感。
另外,我们还利用实时定量PCR技术检测了油菜素内酯合成相关基因D2、OsCPOD和DWARF-4Q以及信号途径相关基因BU1、BZRF、D61、PAVL1、OSLIC和OsMDP1在野生型和sg101幼根中的表达情况(图5)。结果显示,除OsLIC外,所检测的其他与BR合成和信号途径的相关基因的表达均有不同程度的下降。D2在sg101中的表达只有野生型的34%左右,BR信号途径中的BU1、BZRF、D61分别只有野生型的27%、11%和21%。由此可以推断突变体sg101对外源BR处理表现钝感可能是其BR合成途径和信号途径受到影响。

图2 野生型中花11和突变体sg101籽粒的比较Fig. 2. The comparison of grain between Zhonghua 11(WT) and sg101.
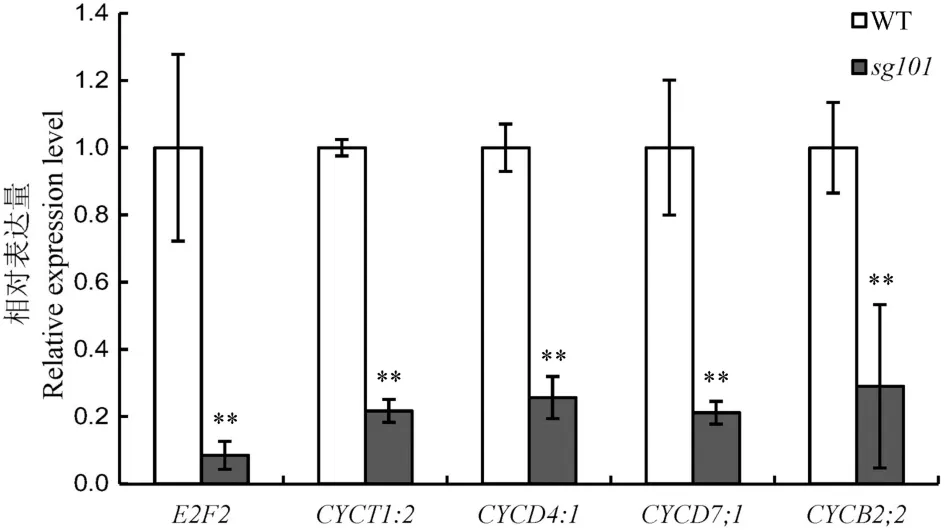
图3 细胞周期相关基因在野生型中花11和突变体sg101中的表达差异Fig. 3. Expression levels of cell cycle-related genes in the wild type(WT) and sg101.
2.5SG101的遗传分析
为了明确水稻矮秆小粒突变体sg101的遗传特性,将突变体sg101分别与籼稻品种TN1、南京06号、9311进行正、反交实验,结果表明不论是正交还是反交,F1单株均表现与野生型类似的表型,说明该表型是受隐性基因控制的(表3);通过对各组合F2代分离群体中正常表型与突变表型植株数量比值进行 χ2测验,其结果均为 χ2<χ20.05,1=3.84,显示野生型植株和突变表型的单株在各 F2群体中的分离比符合 3∶1,表明sg101的突变表型是由单基因控制的质量性状(表3)。
2.6SG101基因定位
为了分离SG101基因,我们用sg101与TN1杂交后代F2群体中选取2600株具突变表型的植株作为定位群体,从中随机挑选30株构建突变体混池,利用本实验室均匀分布于12条染色体上的163对SSR和InDel标记,挑选在sg101和TN1间呈多态性的分子标记用于初步定位。琼脂糖凝胶电泳检测sg101、TN1、F1和突变体混池的PCR产物,发现水稻第1染色体上SSR标记RM8097的PCR扩增产物明显偏向突变体sg101带型,推测RM8907可能与目标基因连锁。将用于构建混池突变体的30株分别提取DNA,并对RM8907及其附近的SSR标记RM6703和RM3738进行验证,结果显示大部分单株在这两个标记上的带型与sg101一致,表明这两个标记亦与目标基因连锁。进一步用93个具sg101表型的F2单株进行验证,发现分子标记RM8097、RM6703和RM3738的交换个体分别为15、10和16,RM3738的交换单株不包含RM6703的单株,而RM8907的交换单株与RM6703和RM3738各不相同,因此推断目标基因SG101介于SSR标记RM8907和RM6703之间,其遗传距离分别为8.1 cM和5.4 cM(图6-A)。
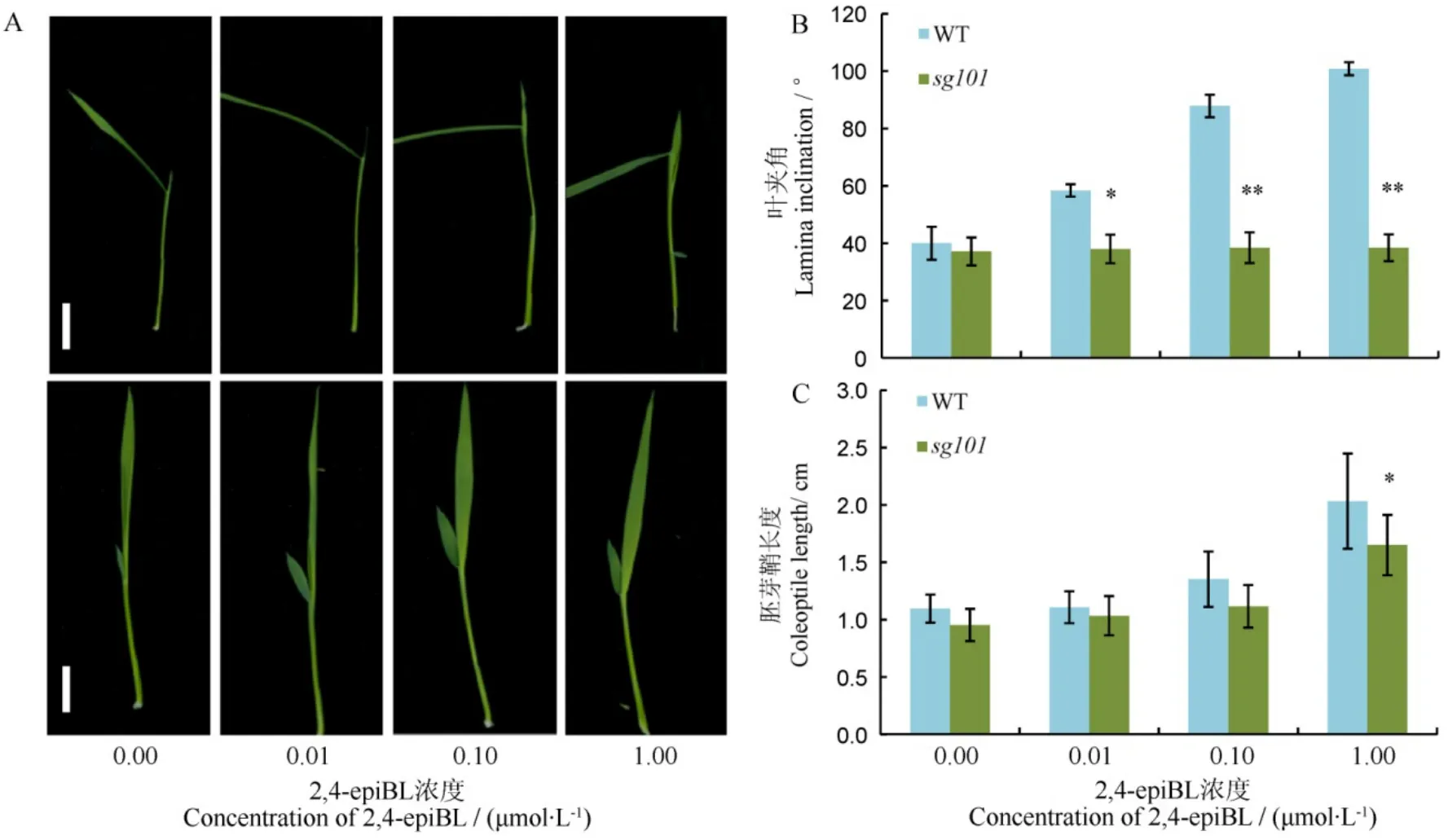
图4 中花11和突变体sg101对2,4-epiBL处理的响应Fig. 4. Lamina joint test to 2,4-epiBL in Zhonghua 11(WT) and sg101.
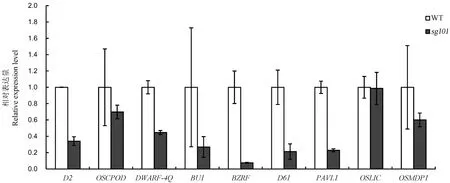
图5 BR合成和信号途径相关基因在野生型中花11和突变体sg101中的表达Fig. 5. Expression levels of BR synthesis and signal transduction pathway genes in Zhonghua 11(WT) and sg101.
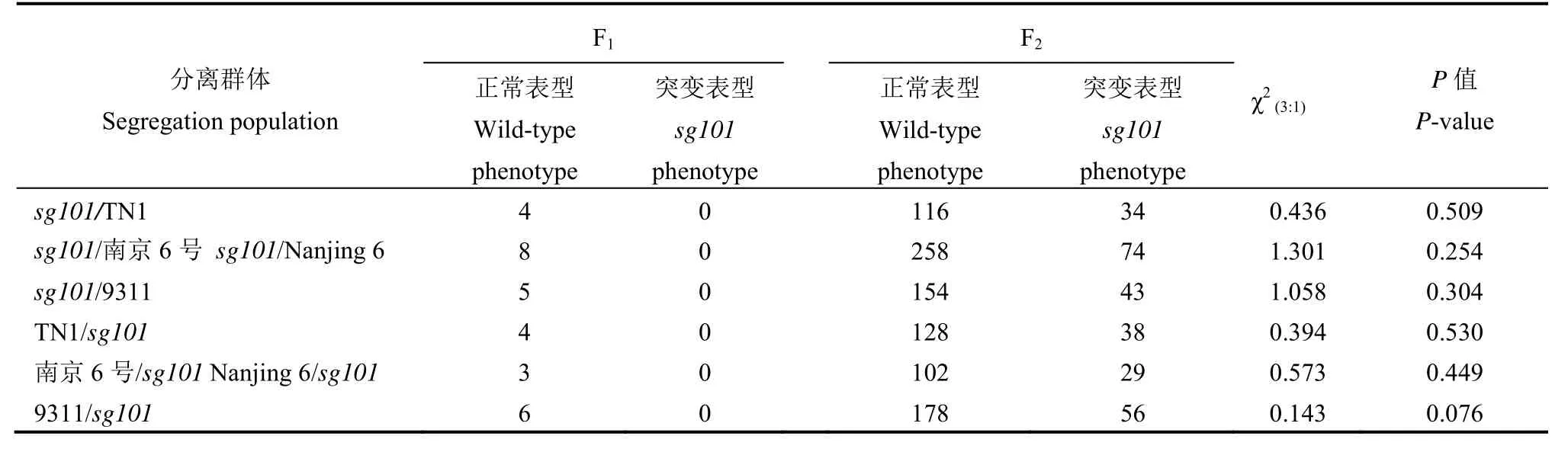
表3 SG101的遗传分析Table 3. Genetic analysis ofSG101.
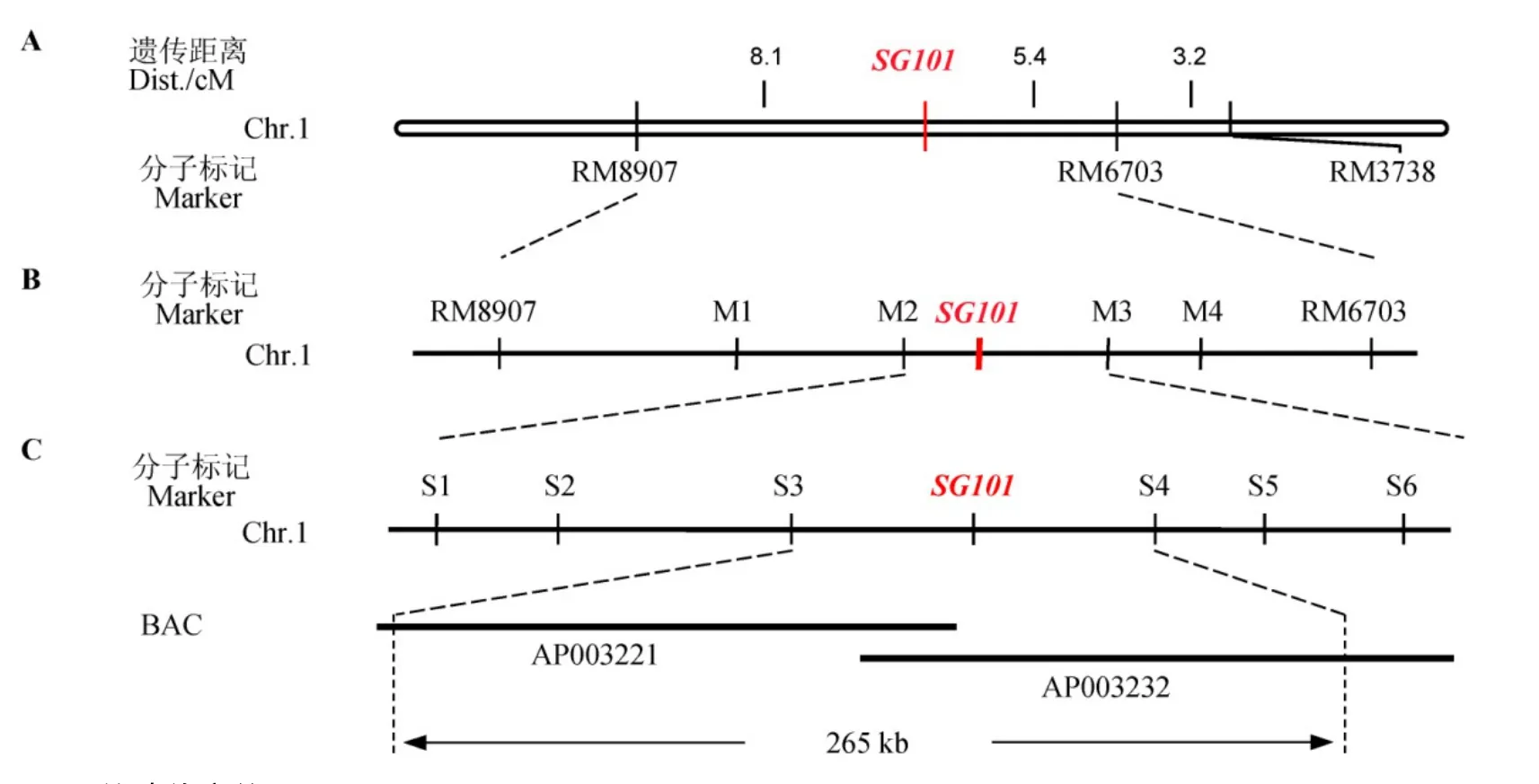
图6 SG101的遗传定位Fig. 6. Molecular mapping of SG101.
为了对 SG101进行精细定位,在 RM8907和RM6703之间开发多个有多态性的InDel标记(表4),利用这些标记对F2群体的2600个突变单株进行连锁分析,将目标基因 SG101最终界定于分子标记S3和 S4之间大约 265 kb的物理区间内。通过Gramene和 Ricedata等网站预测,该区间共有 43个开放阅读框,其中5个编码逆转座子蛋白,1个编码转座子蛋白,10个编码表达的蛋白,2个编码假定的蛋白,7个编码推断的细胞色素P450,3个编码生长素代谢和转运蛋白,3个编码转录因子,1个编码细胞分裂素相关蛋白,还有编码水解酶蛋白、脱氢酶蛋白、微管相关蛋白、类囊体相关蛋白,50S核糖体蛋白、细胞周期相关蛋白以及推断的5个功能蛋白,但该区间内未见已克隆的与籽粒大小相关基因的报道。因此,推测SG101可能是与籽粒大小有关的新基因。
3 讨论
水稻不仅是重要的粮食作物,而且还是植物生物学研究的模式作物。高产一直是水稻育种改良的重要目标,而粒型的大小直接决定粒重的变化,是水稻产量的重要影响因素之一。因此,深入研究水稻粒型变化的分子机理不仅有助于进一步了解水稻产量形成机制,还可为水稻育种提供新的基因资源,具有重要的意义。
近些年,对调控籽粒大小做了很多研究,有研究指出器官大小由细胞数目和细胞大小共同决定,与之对应的是两个基本过程,即细胞增殖和细胞伸长[22,23]。而细胞增殖和细胞伸长与植物体内激素含量相关。调控水稻粒重和灌浆的主效基因TGW6编码一个吲哚乙酸-葡萄糖水解酶,在水稻籽粒发育阶段通过控制生长素(IAA)的供应影响细胞数目和籽粒长度[24]。Bg1-D是生长素的原初响应基因,参与生长素的转运,通过控制细胞数目来影响籽粒大小,从而增加粒重[20]。控制水稻每穗实粒数的主效基因Gn1a,编码一种降解细胞分裂素(CTK)的酶,抑制该基因的表达会使细胞分裂素在分生组织中积累,从而导致株高和每穗粒数的增加[25]。D1编码赤霉素(GA)合成过程中的赤霉素3β羟化酶,突变体d1通过抑制细胞分裂表现出矮秆和小圆粒的表型[26]。
油菜素内酯(BR)是植物体内广泛存在的一类甾醇类激素,在自然界中已发现超过70种BR[27],目前已经筛选很多与BR相关的突变体[28,29]。BR通过促进细胞分裂和细胞扩张正向调控细胞大小,所以水稻BR不敏感突变体和合成受阻突变体均表现出矮秆、叶夹角减小、籽粒变小、叶色变深和育性下降等表型,如d1、d11和d61等。本研究中突变体sg101表现出典型的BR缺失表型,如植株矮小、籽粒变小、结实率下降,生理实验也证明了该突变体对BR反应迟钝,说明SG101可能参与BR调控途径来影响籽粒大小,SG101的克隆和功能研究为BR调控水稻籽粒大小提供新的线索。如D11编码一个细胞色素P450,与几个油菜素内酯合成酶有很高的同源性,该基因的隐性突变导致BR生物合成受阻,突变体d11与野生型相比,植株变矮、籽粒变小[30]。D61/OsBRI1编码BR的受体蛋白BRI1,该基因的突变会影响细胞分裂和伸长,调控器官的发育,导致植株矮小、叶片直立和籽粒变小[31]。BR信号传递途径上,BU1是水稻BR信号传导途径中的正调控因子,过量表达该基因导致籽粒增大[16]。因此,植物激素深刻地影响种子发育并且直接或间接影响种子大小,尤其是细胞分裂素、油菜素内酯和生长素在控制籽粒大小方面起极其重要的作用。
本研究从中花11为背景的突变体库中筛选到一个籽粒变小的突变体sg101,并进行了遗传分析和精细定位。遗传分析表明该基因受一对隐性核基因控制,并将SG101精细定位于第1染色体长臂上,介于分子标记S3和S4之间的265 kb区间内。而第1染色体上已报道的控制水稻株高的sd1、控制水稻每穗粒数的主效基因Gn1a、影响细胞分裂和伸长的D61/OsBRI1均不在该区间内。因此,推断SG101为一新的与籽粒大小相关的基因,在推断的43个开放阅读框中,除6个编码逆转座子/转座子蛋白,12个假定/表达蛋白外,其中的细胞色素P450、生长素代谢和转运蛋白、转录因子、细胞分裂素相关蛋白、微管相关蛋白和细胞周期相关蛋白均可能参与籽粒大小的调控,需要作进一步的精细定位以及候选基因的功能互补。

表4 本研究中SG101定位所用引物Table 4. Primers used for fine mapping of SG101 in this study.
[1] Xing Y,Zhang Q. Genetic and molecular bases of rice yield. Ann Rev Plant Biol,2010,61: 421-442.
[2] Li W,Wu J,Weng S,Zhang Y,Zhang D,Shi C.Identification and characterization of dwarf 62,a loss-of-function mutation in DLT/OsGRAS-32 affecting gibberellin metabolism in rice. Planta,2010,232(6):1383-1396.
[3] Mao H,Sun S,Yao J,Wang C,Yu S,Xu C,Li X,Zhang Q. Linking differential domain functions of the GS3 protein to natural variation of grain size in rice. Proc Natl Acad Sci,2010,107(45): 19579-19584.
[4] 茆海亮. 水稻粒形基因GS3的功能研究. 武汉: 华中农业大学,2010.Mao H L. Functional study of rice grain shape gene GS3.Wuhan: Huazhong Agricultural University,2010. (in Chinese with English abstract)
[5] 石珍源. 水稻粒型QTL qGW12的精细定位和粒长调控基因SG4的克隆与功能验证. 北京:中国农业科学院,2015.Shi Z Y. Fine mapping of rice width locus GW12 and functional verification of a grain length gene SG4 in rice.Beijing: Chinese Academic of Agricultural Sciences,2015. (in Chinese with English abstract)
[6] Zuo J,Li J. Molecular genetic dissection of quantitative trait loci regulating rice grain size. Ann Rev Genet,2014,48: 99-118.
[7] Zhang X,Wang J,Huang J,Lan H,Wang C,Yin C,Wu Y,Tang H,Qian Q,Li J,Zhang H. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice. Proc Natl Acad Sci,2012,109(52): 21534-21539.
[8] Qi P,Lin Y,Song X,Shen J,Huang W,Shan J,Zhu M,Jiang L,Gao J,Lin H. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3. Cell Res,2012,22(12): 1666-1680.
[9] Wang Y,Xiong G,Hu J,Jiang L,Yu H,Xu J,Fang Y,Zeng L,Xu E,Xu J,Ye W,Meng X,Liu R,Chen H,Jing Y,Wang Y,Zhu X,Li J,Qian Q. Copy number variation at the GL7 locus contributes to grain size diversity in rice.Nat Genet,2015,47(8): 944-948.
[10] Hu,Wang Y,Fang Y,Zeng L,Xu J,Yu H,Shi Z,Pan J,Zhang D,Kang S,Zhu L,Dong G,Guo L,Zeng D,Zhang G,Xie L,Xiong G,Li J,Qian Q. A rare allele of GS2 Enhances grain size and grain yield in rice. Mol Plant,2015,8(10): 1455-1465.
[11] Song X,Huang W,Shi M,Zhu M,Lin H,. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet,2007,39: 623-630.
[12] Yan S,Zou G,Li S,Wang H,Liu H,Zhai G,Guo P,Song H,Yan C,Tao Y. Seed size is determined by the combinations of the genes controlling different seed characteristics in rice. Theor Appl Genet,2011,123(7):1173-1181.
[13] Weng J F,Gu S H,Wan X Y,Gao H,Guo T,Su N,Lei C L,Zhang X,Cheng Z J,Guo X P,Wang J L,Jiang L,Zhai H Q,Wan J M. Isolation and initial characterization of GW5,a major QTL associated with rice grain width and weight. Cell Res,2008,18(12): 1199-1209.
[14] Li Y,Fan Ch,Xing Y,Jiang Y,Luo L,Sun L,Shao D,Xu C,Li X,Xiao J,He Y,Zhang Q. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet,2011,43(12): 1266-1269.
[15] Wang S,Wu K,Yuan Q,Liu X,Liu Z,Lin X,Zeng R,Zhu H,Dong G,Qian Q,Zhang G,Fu X. Control of grain size,shape and quality by OsSPL16 in rice. Nat Genet,2012,44: 950-954.
[16] Yamamuro C,Ihara Y,Wu X,Noguchi T,Fujioka S,Takatsuto S,Ashikari M,Kitano H,Matsuoka M. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. Plant Cell,2000,12(9): 1591-1605.
[17] Tanaka A,Nakagawa H,Tomita C,Shimatani Z,Ohtake M,Nomura T,Jiang ,Dubouzet J,Kikuchi S,Sekimoto H,Yokota T,Asami T,Kamakura T,Mori M.BRASSINOSTEROID UPREGULATED1,encoding a helix-loop-helix protein,is a novel gene involved in brassinosteroid signaling and controls bending of the lamina joint in rice. Plant Physiol,2009,151(2):669-680.
[18] Duan P,Rao Y,Zeng D,Yang Y,Xu R,Zhang B,Dong G,Qian Q,Li Y. SMALL GRAIN 1,which encodes a mitogen-activated protein kinase kinase 4,influences grain size in rice. Plant J,2014,77(4): 547-557.
[19] Liu S,Hua L,Dong S,Chen H,Zhu X,Jiang J,Zhang F,Li Y,Fang X,Chen F. OsMAPK6,a mitogen-activated protein kinase,influences rice grain size and biomass production. Plant J,2015,84(4): 672-681.
[20] Liu L,Tong H,Xiao Y,Che R,Xu F,Hu B,Liang C,Chu J,Li J,Chu C. Activation of Big Grain1 significantly improves grain size by regulating auxin transport in rice.Proc Natl Acad Sci,2015,112(35): 11102-11107.
[21] Murray M G,Thompson W F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res,1980,8(19): 4321-4326.
[22] Horiguchi G,Ferjani A,Fujikura U,Tsukaya H.Coordination of cell proliferation and cell expansion in the control of leaf size in Arabidopsis thaliana. J Plant Res,2006,119: 37-42.
[23] Horvath BM,Magyar Z,Zhang Y,Hamburger AW,Bako L,Visser RG,Bachem CW,Bogre L. EBP1 regulates organ size through cell growth and proliferation in plants.EMBO J. 2006,25: 4909-4920.
[24] Ishimaru K,Hirotsu N,Madoka Y,Murakami N,Hara N,Onodera H,Kashiwagi T,Ujiie K, Shimizu B,Onishi A,Miyagawa H,Katoh E. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat Genet,2013,45(6):707-711.
[25] Ashikari M,Wu J,Yano M,Sasaki T,Yoshimura A. Rice gibberellin-insensitive dwarf mutant gene Dwarf 1 encodes the α-subunit of GTP-binding protein. Proc Natl Acad Sci,1999,96(18): 10284-10289.
[26] Fujisawa Y,Kato T,Ohki S,Ishikawa A,Kitano H,Sasaki T,Asahi T,Iwasaki Y. Suppression of the heterotrimeric G protein causes abnormal morphology,including dwarfism,in rice. Proc Natl Acad Sci,1999,96(13): 7575-7580.
[27] Andrzej B. Metabolism of brassinosteroids in plants.Plant Physiol Biochem,2007,45(2): 95-107.
[28] Hong Z,Ueguchi-Tanaka M,Shimizu-Sato S,Inukai Y,Fujioka S,Shimada Y,Takatsuto S,Agetsuma M,Yoshida S,Watanabe Y,Uozu S,Kitano H,Ashikari M,Matsuoka M. Loss-of-function of a rice brassinosteroid biosynthetic enzyme,C-6 oxidase,prevents the organized arrangement and polar elongation of cells in the leaves and stem. Plant J,2002,32(4): 495-508.
[29] Hong Z,Ueguchi-Tanaka M,Umemura K,Uozu S,Fujiok S,Takatsuto S,Yoshida S,Ashikari M,Kitano H,Matsuoka M. A rice brassinosteroid-deficient mutant,ebisu dwarf (d2),is caused by a loss of function of a new member of cytochrome P450. Plant Cell,2003,15(12):2900-2910.
[30] Tanabe S,Ashikari M,Fujioka S,Takatsuto S,Yoshida S,Yano M,Yoshimura A,Kitano H,Matsuoka M,Fujisawa Y,Kato H,Iwasaki Y. A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant,dwarf11,with reduced seed length. Plant Cell,2005,17(3): 776-790.
[31] Hong Z,Ueguchi-Tanaka M,Fujioka S,Takatsuto S,Yoshida S,Hasegawa Y,Ashikari M,Kitano H,Matsuoka M. The rice brassinosteroid deficient dwarf2 mutant,defective in the rice homolog of Arabidopsis DWARF1,is rescued by the endogenously accumulated alternative bioactive brassinosteroid,dolichosterone. Plant Cell,2005,17(8): 2243-2254.
Identification and Fine Mapping of Small Grain Gene SG101 in Rice (Oryza sativa L.)
WANG Yuqiong1,#,YANG Yaolong1,#,LENG Yujia1,HUANG Lichao1,CHEN Long1,DAI Liping1,TU Zhengjun1,GAO Yihong1,HU Jiang1,ZHU Li1,ZHANG Guangheng1,REN Deyong1,GAO Zhenyu1,DONG Guojun1,CHEN Guang1,GUO Longbiao1,YE Guoyou2,QIAN Qian1,*,ZENG Dali1,*
(1 State Key Laboratory of Rice Biology,China National Rice Research Institute,Hangzhou 310006,China; 2International Rice Research Institute,Makati City,Philippines; #These authors contributed equally to this work; *Corresponding author,E-mail: dalizeng@126.com; qianqian188@hotmail.com)
【Objective】Grain size is an important agronomic trait in determining grain yield. The characterization and identification of grain size related genes will be beneficial to expound the genetic regulatory mechanisms behind yield formation in rice. 【Method】Here,we report the characterization of a rice (Oryza sativa L.) mutant,small grain 101(sg101),gained by EMS mutagenesis for Zhonghua 11. The effects of sg101 on grain size,panicle-related traits,cell number and size of lemma were assessed by morphological and cytological methods. The responses to brassinolide(BR)were detected based on the variation in leaf angle and coleoptile length. The expression level of BR signal and synthesis related genes were tested by RT-PCR. Map-based cloning was executed for fine mapping of SG101.【Result】The grain length,grain width and grain weight were significantly reduced in sg101. Moreover,sg101 showed decreased panicle length,secondary rachis branch number,plant height and seed setting rate. The paraffin section observation under a scanning electron microscope indicated that the cell number in sg101 was significantly decreased,while the cell size was similar to wild type,suggesting that SG101 affected the grain size by regulating the cell division. Quantitative RT-PCR analysis displayed that the expression of cell cycle related genes was reduced in sg101. After treatment with BR,sg101 showed smaller leaf angle and shorter coleoptile than those of the wild type and the expression of BR biosynthetic and signal pathway related genes was down-regulated in sg101. Genetic analysis revealed that sg101 phenotype was controlled by a single recessive nuclear gene.【Conclusion】SG101 was mapped on the long arm of chromosome 1 between two STS markers S5 and S6 with a 265 kb physical distance. It laid a base for the further cloning and functional analysis of SG101.
rice; small grain; sg101; brassinolide
Q343.5; S511.032
A
1001-7216(2017)06-0580-10
10.16819/j.1001-7216.2017.6152
2016-11-15; 修改稿收到日期:2017-04-03。
国家自然科学基金资助项目(91435105,91735303,31661143006); 中国农业科学院科技创新工程资助项目。
猜你喜欢
作文成功之路·小学版(2020年7期)2020-08-24 08:19:28
天津中医药(2020年5期)2020-06-01 12:16:14
中成药(2017年4期)2017-05-17 06:09:27
安徽医科大学学报(2016年12期)2017-01-15 14:21:44
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
中国民族医药杂志(2016年6期)2016-05-09 08:52:57
山东医药(2015年40期)2015-02-28 14:28:45
应用化工(2014年5期)2014-08-08 13:10:58
湖南农业科学(2014年4期)2014-02-27 14:27:07
祝你幸福·午后版(2013年7期)2013-07-18 07:20:02
