
黑色素细胞中过量表达lpa-miR-nov-66对TYRP2的影响
2017-12-01 02:33:03杨姗姗范瑞文焦丁兴董常生
中国兽医杂志 2017年10期
杨姗姗 , 范瑞文 , 焦丁兴 , 董常生
(1.山西农业大学动物科技学院 , 山西 太谷 030801 ; 2.河北省清河县农业局 ,河北 清河 054000)
黑色素细胞中过量表达lpa-miR-nov-66对TYRP2的影响
杨姗姗1, 范瑞文1, 焦丁兴2, 董常生1
(1.山西农业大学动物科技学院 , 山西 太谷 030801 ; 2.河北省清河县农业局 ,河北 清河 054000)
本研究旨在探讨lpa-miR-nov-66对黑色素生成相关基因TYRP2的影响。采用细胞转染技术,在羊驼黑色素细胞中过表达lpa-miR-nov-66,运用反转录聚合酶链反应(RT-PCR)、实时荧光定量聚合酶链反应(quantitative real-time polymerase chain reaction, qRT-PCR)技术和免疫印迹法(western blotting)技术分别检测转染lpa-miR-nov-66后黑色素细胞内TYRP2在mRNA和蛋白水平的表达差异。RT-PCR结果显示:TYRP2的部分片段扩增,扩增片段长度为151 bp。qRT-PCR和免疫印迹(western blotting)结果显示:与对照组相比,实验组TYRP2在mRNA表达水平显著降低,降低0.44倍;蛋白表达水平也显著降低,降低0.78倍。结果表明,lpa-miR-nov-66可以影响TYRP2的表达,可能参与了动物毛色的形成过程。
mircoRNA ; lpa-miR-nov-66 ; 黑色素细胞 ; TYRP2
数百年来,动物饲养者一直在探讨以行为学、形态学、生理学、病理学特征为目标的表型变化。其中毛用动物(羊驼和绵羊)的毛色得到了相当大的重视[1]。动物毛色主要由沉着在皮肤内黑色素的数量及其种类决定。动物黑色素主要分为真黑色素和褐黑色素两种,两者的相对数量及分布决定了动物皮肤及被毛的颜色[2]。许多研究已经明确了基因和microRNAs (miRNAs) 在人类和其他脊椎动物毛发和皮肤颜色中的重要作用[3]。microRNAs (miRNAs),一种小RNA分子,长度在22~25 nt,是调控大多数细胞生物过程的必需因子,可以调节人类大约三分之一的蛋白质编码基因[4-5]。因此对miRNAs作用的研究显得极为重要。miRNA对基因表达的调控主要有两种方式,一种是加速靶向基因mRNA的降解,一种是阻碍靶向基因mRNA的翻译,而哺乳动物主要是以后者为主[6]。许多研究表明,miRNAs与调控动物毛色基因有相互作用关系。例如Zhu等通过实验验证miR-25抑制了MITF的表达,同时黑色素含量也呈现降低的趋势[7]。Dong等人通过通过显微注射的方法将miR-137注射到C57黑色小鼠胚胎内,发现后代的转基因小鼠的毛色明显变浅[8]。实验室前期通过对白色和棕色羊驼皮肤的深度测序,获得了一系列可能参与毛色调控的mircoRNAs。lpa-miR-nov-66就是其中的一个[9]。本文通过细胞转染技术在羊驼黑色素细胞内过量表达lpa-miR-nov-66,以探究其过量表达对黑色素相关基因TYRP2的影响。
1 材料与方法
1.1 主要材料与试剂 黑色素细胞培养基(Sciencell公司,美国),TRIZol(Invitrogen公司,美国);Plasmid 提取试剂盒(Qiagen公司,美国);根据lpa-miR-nov-66序列合成的真核表达载体(Invitrogen公司,美国);RT-PCR Kit (TaKaRa公司),TYRP2多克隆兔抗抗体(Santa Cruz Biotechnology公司),HRP标记的羊抗兔二抗 (北京康为世纪生物科技有限公司);总蛋白提取试剂盒 (北京碧云天生物技术研究所);蛋白Marker (Fermentas公司,美国);ECL发光试剂盒 (北京康为世纪生物科技有限公司)。
1.2 试验方法
1.2.1 细胞培养 羊驼黑色素细胞由本实验室保存并提供[10]。从液氮中取出保存的绵羊黑色素细胞,复苏后,于37 ℃,5%CO2培养箱中培养。待细胞生长至90%左右时进行传代。
1.2.2 细胞转染 接种106个/孔细胞于6孔细胞培养板,待细胞生长至60%~75%时,进行细胞转染。按照转染试剂盒说明书进行转染,其比例为转染试剂∶质粒=1 μl∶3μg。转染6 h后,用含有两倍血清的培养基终止转染,12 h后移去两倍血清的培养基,用完全培养基继续培养56 h后收取样品。
1.2.3 总RNA的提取及反转录合成cDNA 转染细胞后继续培养56 h,用TRIZol法提取转染细胞内的总RNA,并用Nanodrop1000测其品质与浓度。按照反转录试剂盒说明书进行反转录。
1.2.4 引物设计与合成 Real-time PCR检测 利用Premier 3.0 在线软件,根据NCBI提交的羊驼TYRP2的序列设计Real-time PCR引物,并将β-actin设为内参引物(见表1)。

表1 目的基因和内参基因引物序列
1.2.5 实时荧光定量聚合酶链反应体系的建立 按照RT-PCR Kit说明书操作,每个样品3个重复,反应体系为25 μL。反应条件:预变性95 ℃ 30 s,95 ℃ 5 s,55 ℃ 30 s,72 ℃ 15 s,循环数为40个,终延伸72 ℃ 10 min。试验数据采用△△Ct法:
目的基因的相对表达水平=

△Ct(试验组)=Ct(目的基因)-Ct(内参基因) △Ct(对照组)=Ct(目的基因)-Ct(内参基因)
△△Ct=△Ct(实验组)-△Ct(对照组)目的基因的相对表达水平 = 2-△△Ct
所有数据用SPSS 17.0软件进行统计分析,荧光实时定量 PCR 结果均用均值±标准差(Means±SD)表示, 其中各基因的表达量所示结果均由内参基因β-actin的表达量进行校正,所有数据采用SPSS 17.0统计软件进行单因素方差分析检验。
1.2.6 蛋白免疫印迹试验(Western Blotting) 按照总蛋白提取试剂盒说明书提取转染后黑色素细胞的总蛋白,并测其浓度。上样后,进行SDS-PAGE电泳,转膜至PVDF膜。PVDF膜用5%脱脂奶粉进行封闭,时间为1 h。加入首抗,4 ℃,过夜孵育。用TBST洗膜,5 min×6次,放入HRP标记的二抗中,37 ℃,孵育1 h。用TBST洗膜,5 min×6次。将膜取出,用ECL发光试剂盒显色后曝光。对获得的目的条带进行扫描分析。利用Image-ProPlus 6.0软件测定目的蛋白的灰度值,并进行半定量分析。数据用Means±SE表示,全部数据采用SPSS 统计软件进行单因素方差分析检验。
2 结果
2.1 羊驼黑色素细胞的生长观察 在合成质粒时,所用载体为可在哺乳动物细胞表达的pcDNA6.2-GW/ EmGFPmiR并连有1个启动基因绿色荧光蛋白(green fluorescent protein, GFP)和增强性启动子(promoter)CMV ,在由attBI-和attBII-切开的载体间插入了lpa-miR-nov-66 序列 (图1)。

图1 pcDNATM6.2-GW/EmGFP-miR 表达质粒结构
正常羊驼黑色素细胞在复苏后,贴壁生长,伸展性能较好,生长状态良好。24 h后细胞呈现典型的树突状。随着培养时间的延长,细胞密度增大,细胞形态呈长梭形(图2A)。图2B和2C显示,转染效率较高,并且细胞形态单一,生长良好。
2.2 转染lpa-miR-nov-66对TYRP2 mRNA影响的测定 将提取的总RNA进行凝胶电泳检测,图3A中可见3条清晰的条带,并且28 s的亮度高于18 s,说明RNA质量较好,可用于后续试验。RT-PCR结果(图3B)显示,扩增产物片段大小为151 bp,与预期目的基因的片段大小相符合。qRT-PCR结果显示,与NC组中TYRP2基因mRNA表达水平相比,lpa-miR-nov-66组TYRP2的mRNA表达量呈下降趋势,其mRNA的表达量是NC组0.44倍,差异极其显著(Plt;0.001) (图3C)。
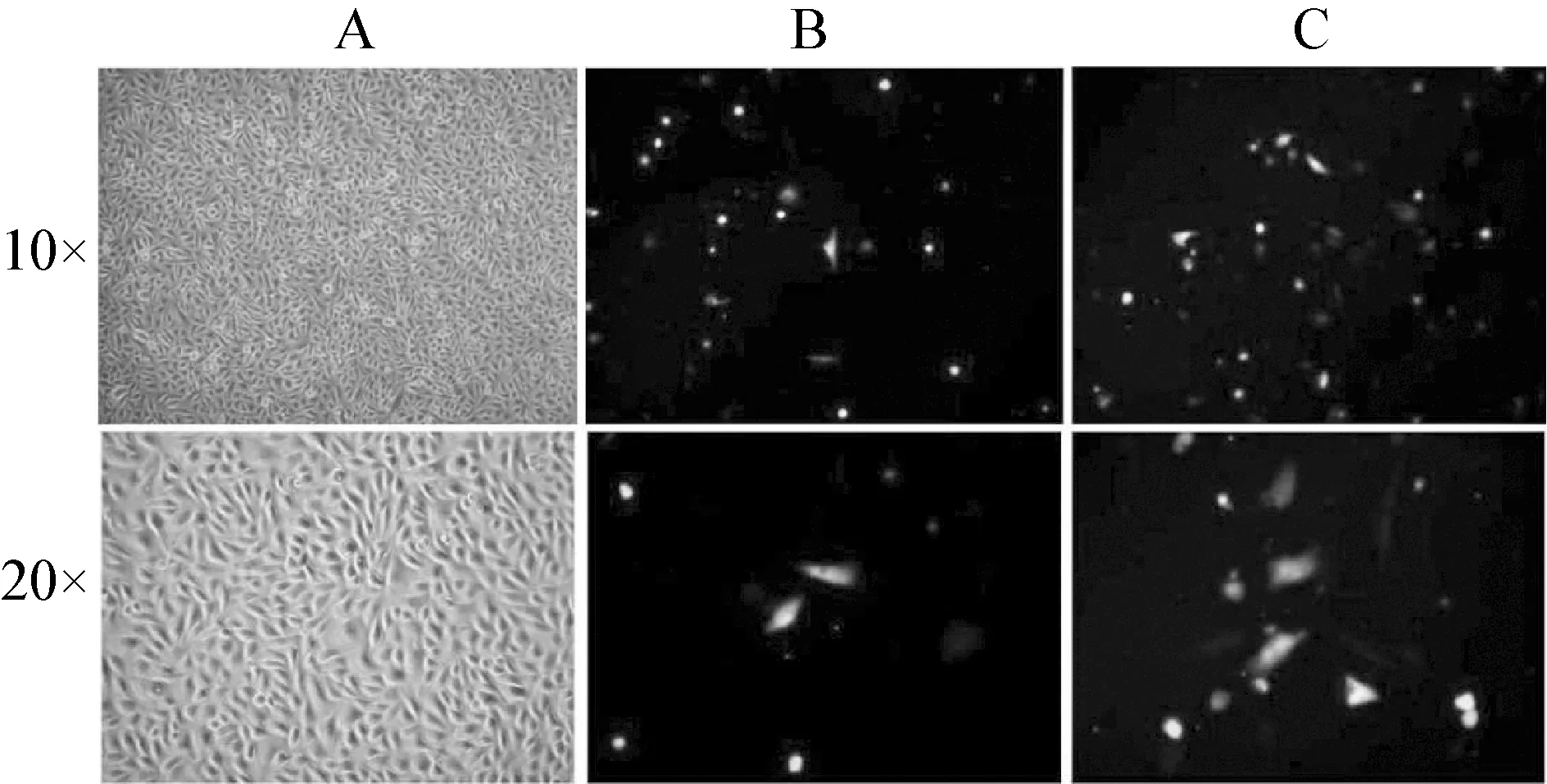
图2 羊驼黑色素细胞
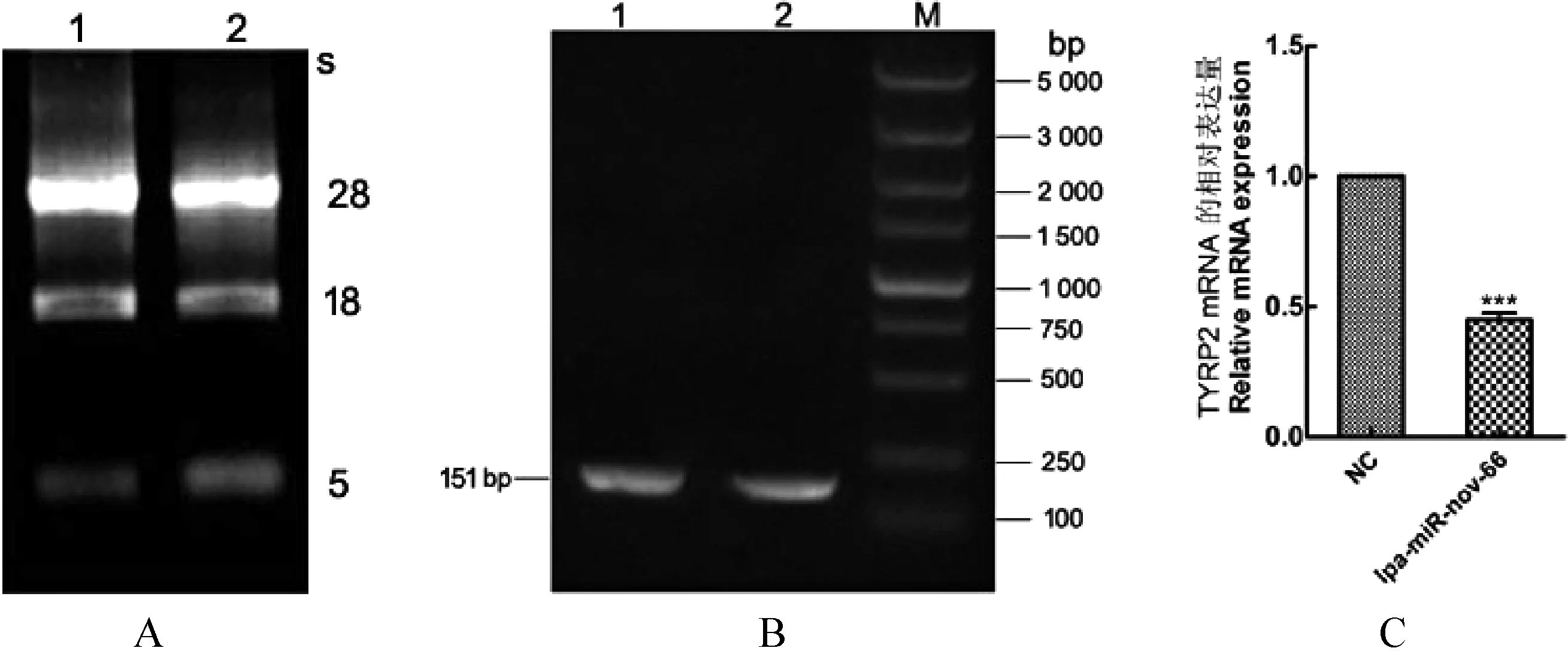
图3 TYRP2 mRNA在转染NC和lpa-miR-nov-66质粒中的表达检测和分析
2.3 转染lpa-miR-nov-66对TYRP2蛋白表达量影响的测定 蛋白质免疫印迹结果表明,转然后黑色素细胞内的TYRP2蛋白质可以与TYRP2多克隆抗体结合 (图4A)。结果显示,与NC组相比,lpa-miR-nov-66组TYRP2的蛋白表达水有显著降低,NC组的0.78倍(Plt;0.01) (图4B)。
3 讨论
TYRP2/Dct作为小鼠的 slaty/Dct (Slt)基因座编码,可以将多巴色素异构化为5,6-二羟吲哚-2羧酸(5,6-dihydroxyindole-2-carboxylic acid,DHICA)。因此slaty/Dct(Slt)野生型小鼠毛发的真黑素富含DHICA。研究者证实2,3,5-吡咯三羧酸(pyrrole-2,3,5-tricarboxylic acid, PTCA)可作为含有DHICA色素的特有标记物[11]。Lamoreux 等对具有Slaty基因座的黑色、棕色、黑chinchilla和棕chinchilla野生型小鼠的毛发进行分析,发现PTCA含量含量较高。这都证明了小鼠的TYRP2可决定色素中DHICA的含量[12]。Slaty等位点变异所形成的纯合子小鼠,TYRP2功能较正常低3倍到10倍,甚至失去功能。分析发现Slaty变异的基因型小鼠毛发总色素量没有变化,只有PTCA值大量降低,导致PTCA和总色素量的比值只有原来的1/4到1/6,形成的真黑素中DHICA 含量非常少。人类TYRP2/Dct基因存在多态性并影响人类毛发的色素沉积[13]。但是,TYRP2/Dct多态性在人类皮肤和毛发浅色发生机制中所具有的重要地位仍然不能确定。令人感兴趣的是,人类棕色或黑色毛发毛囊色素细胞不能表达TYRP2/Dct蛋白。因此,研究者推测人类棕色和黑色毛发的形成不需要TYRP2的参与[14]。

图4 TYRP2 蛋白在转染NC和lpa-miR-nov-66质粒中的表达量
本试验利用细胞转染技术,将lpa-miR-nov-66转染羊驼黑色素细胞,进而研究了lpa-miR-nov-66对黑色素细胞内黑色素生成的影响。试验通过qRT-PCR 以及Western Boltting方法对黑色素细胞中的与黑色素生成有关基因TYRP2进行检测,结果表明,lpa-miR-nov-66的过表达可以使黑色素细胞中TYRP2 mRNA和蛋白表达水平均有下降,这暗示了lpa-miR-nov-66可能会通过调控TYRP2影响黑色素颗粒的合成。研究表明,TYRP2对黑色素细胞的存活具有一定的支持作用[15]。TYRP2作为黑色素合成过程中的关键酶,可以稳定酪氨酸酶的活性,维持黑色素小体结构的完整性[16],其表达水平的降低会引起黑色素含量的减少。lpa-miR-nov-66过表达下调了TYRP2在翻译及转录水平的表达,说明在黑色素细胞中过表达lpa-miR-nov-66会抑制TYRP2的表达,从而对黑色素细胞内黑色素颗粒的生成产生一定的抑制作用。然而,lpa-miR-nov-66的过量表达对黑色素细胞中TYRP2的影响是否还存在其他的作用途径,相关途径之间是否存在交叉等一些问题目前尚未见报道,还有待于进一步的研究。
4 结论
通过在羊驼黑色素细胞中过表达lpa-miR-nov-66,发现过表达的lpa-miR-nov-66可以降低TYRP2 mRNA和蛋白的表达。结果表明,lpa-miR-nov-66与绵羊毛色形成过程具有一定的相关性。
[1] McGregor B. Production, attributes and relative value of alpaca fleeces in southern Australia and implications for industry development [J]. Small Ruminant Research, 2006, 61(2):93-111.
[2] Simon J D, Peles D N. The red and the black [J]. Accounts of Chemical Research, 2010, 43(11): 1 452-1 460.
[3] Ambros V. The functions of animal microRNAs [J]. Nature, 2004, 431(7006):350-355.
[4] Friedman R C, Farh K K-H, Burge C B,etal. Most mammalian mRNAs are conserved targets of microRNAs [J]. Genome research,2009, 19(1):92-105.
[5] Lewis B P, Burge C B, Bartel D P. Conserved seed pairing, often flanked by adenosines, indicates that thouands of human genes are microRNA targets [J]. Cell,2005, 120(1):15-20.
[6] Bartel D P. MicroRNAs: genomics, biogenesis, mechanism, and function [J]. cell, 2004, 116(2):281-297.
[7] Zhu Z, He J, Jia X,etal. MicroRNA-25 functions in regulation of pigmentation by targeting the transcription factor MITF in alpaca (lt; igt; Lama pacoslt;/igt;) skin melanocytes [J]. Domestic animal endocrinology, 2010, 38(3):200-209.
[8] Dong C, Wang H, Xue L,etal. Coat color determination by miR-137 mediated down-regulation of microphthalmia-associated transcription factor in a mouse model [J]. RNA, 2012, 18(9):1 679-1 686.
[9] Tian X, Jiang J, Fan R,etal. Identification and characterization of microRNAs in white and brown alpaca skin [J]. BMC Genomics, 2012, 13:555.
[10] Bai R, Sen A, Yu Z,etal. Validation of methods for isolation and culture of alpaca melanocytes: a novel tool for in vitro studies of mechanisms controlling coat color [J]. Asian Aust J Anim Sci. 2010, 23: 430-436.
[11] Ito S,Wakamatsu K. Chemical Degradation of Melanins: Application to Identification of Dopamine-melanin [J].Pigment Cell Research, 2006, 11(2):120-126.
[12] Lamoreux M, Wakamatsu K,Ito S. Interaction of major coat color gene functions in mice as studied by chemical analysis of eumelanin and pheomelanin [J].Pigment Cell Research, 2002, 14(1):23-31.
[13] McEvoy B, Beleza S,Shriver M D. The genetic architecture of normal variation in human pigmentation: an evolutionary perspective and model [J]. Hum Mol Genet, 2006, 15(suppl2):R176-R181.
[14] Commo S, Gaillard O, Thibaut S,etal. Absence of TRP-2 in Melanogenic Melanocytes of Human Hair [J].Pigment Cell Research, 2004, 17(5):488-497.
[15] Slominski A, Wortsman J, Plonka P M,etal. Hair follicle pigmentation [J]. Journal of Investigative Dermatology, 2005, 124(1):13-21.
[16] Hearing V J. Biochemical control of melanogenesis and melanosomal organization [J]. Journal of Investigative Dermatology Symposium Proceedings, 1999, Nature Publishing Group: 24-28.
TheInfluencesofover-expressinglpa-miR-nov-66onTYRP2inmelanocytesofalpaca
YANG Shan-shan1, FAN Rui-wen1, JIAO Ding-xing2, DONG Chang-sheng1
(1.College of Animal Science and Veterinary Medicine, Shanxi Agricultural University, Taigu 030801, China; 2.Agricultural Bureau of Hebei Province, Qinghe 054000,China )
The molecular mechanisms underlying the formation of coat colors in animals are poorly understood. Recent studies have demonstrated that microRNAs play important roles in the control of melanogenesis and coat color in mammals. In order to investigate the influences of lpa-miR-nov-66 on TYRP2 in melanocytes. The melanocytes were cultured and transfected with the lpa-miR-nov-66 or NC plasmid. Reverse transcription-polymerase chain reaction (RT-PCR), quantitative real-time polymerse chain reaction (qRT-PCR) and western blotting were used to detect the different expression of TYRP2 in melanocytes transfected with the lpa-miR-nov-66 or NC plasmid. A partial sequence of TYRP2 gene with the length of 151 bp was successfully amplified by RT-PCR. qRT-PCR results showed that the expression level of TYRP2 mRNA was 0.44 fold changes in the melanocytes transfected with lpa-miR-nov-66 plasmid compared with the melanocytes transfected with NC plasmid (Plt;0.001). Western blotting showed that the protein level of TYRP2 was 0.78 fold changes in in the melanocytes transfected with lpa-miR-nov-66 plasmid compared with the melanocytes transfected with NC plasmid (Plt;0.01). All those results suggest that lpa-miR-nov-66 can affect the expression of TYRP2 both at mRNA and protein levels and has a potential role in coat color regulation.
mircoRNA ; lpa-miR-nov-66 ; melanocytes ; TYRP2
DONG Chang-sheng
S852.1
A
0529-6005(2017)10-0024-04
2017-03-29
国家“863”计划 (2013AA102506); 公益性行业 (农业) 科研专项 (201303119); 山西省研究生教育创新项目 (2016BY071)
杨姗姗(1987-), 女,博士生, 研究方向为羊驼毛色, E-mail:shanshan0321@163.com
董常生,E-mail: cs-dong@sxau.edu.cn
猜你喜欢
小学时代·科学小问号(2024年5期)2024-06-26 00:00:00
学苑创造·A版(2024年5期)2024-06-10 21:55:57
阅读与作文(小学低年级版)(2019年12期)2019-12-26 01:55:34
奥秘(创新大赛)(2019年8期)2019-11-30 04:38:46
小天使·四年级语数英综合(2019年9期)2019-11-09 05:48:52
中国生殖健康(2019年11期)2019-01-07 01:27:32
青少年科技博览(中学版)(2017年5期)2018-02-28 21:23:59
作文评点报·低幼版(2018年39期)2018-01-08 07:51:46
天然产物研究与开发(2016年11期)2016-06-15 20:29:15
中国医疗美容(2015年2期)2015-07-19 10:11:59
