
景观格局特征与区域生物多样性的关系研究
2017-11-15 01:45:35林世滔谢弟炳刘郁林陈文波
生态环境学报 2017年10期
林世滔,谢弟炳,刘郁林,陈文波*
1. 江西环境工程职业学院,江西 赣州 341000;2. 江西农业大学景观与环境生态研究中心,江西 南昌 330045;3. 南昌市景观与环境重点实验室,江西 南昌 330045;4. 江西应用职业技术学院,江西 赣州 341000
景观格局特征与区域生物多样性的关系研究
林世滔1,2,3,谢弟炳4,刘郁林1,陈文波2,3*
1. 江西环境工程职业学院,江西 赣州 341000;2. 江西农业大学景观与环境生态研究中心,江西 南昌 330045;3. 南昌市景观与环境重点实验室,江西 南昌 330045;4. 江西应用职业技术学院,江西 赣州 341000
景观格局决定景观的功能和生态过程,影响着景观内的物种流、信息流、能源流,从而对生物多样性产生显著作用。以鄱阳湖生态经济区为研究区域,运用SPSS 17.0和Fragstats 3.3等统计分析软件,从38个县区的景观特征中选取具有生态意义的景观指数与生物多样性进行相关分析,探讨景观格局特征与区域生物多样性的关系。结果表明,(1)景观类型水平上景观指数与生物多样性之间的关系表现不一,耕地部分指标如PLAND、NP、COHESION(0.476**,P<0.01)与区域生物多样性指数呈正相关,耕地的景观特征与各物种大部分指标之间的相关关系呈不显著的负相关,表现出耕地景观与生物多样性具有复杂关系。区域内林地PLAND(0.439**,P<0.01)、NP、COHESION(0.609**,P<0.01)与区域生物多样性指数呈正相关,林地PD、LPI、AWMSI与区域生物多样性指数呈弱程度的负相关。林地景观除了跟植物的PLAND(0.655**,P<0.01)、COHESION(0.729**,P<0.01)呈极显著正相关外,与其他多数指标呈负相关。湿地景观大部分指标与区域生物多样性指数呈显著正相关。(2)景观水平上景观指数与生物多样性之间的相关性分析结果表明,区域的景观特征除MPS与区域生物多样性指数呈负相关外,其他景观指标都与区域生物多样性呈正相关,其中TA、NP、LSI、SHDI与区域生物多样性关系较为显著,说明景观的面积大小、数量、形状、多样性对生物多样性和各种生物过程都有较大影响。区域景观特征与生物多样性具有较好的相关性,关系较为复杂。区域内的湿地景观、林地景观是区域生物种的主要栖息场所,同时,作为产粮主产区,耕地景观的影响也不容忽视。因此,保护区域内的湿地、林地、耕地景观对于维持区域生物多样性具有重要作用。该研究对区域内生物多样性的保护和管理具有一定的理论指导意义。
景观格局;生物多样性;鄱阳湖生态经济区
生物多样性为人类生存和发展提供物质基础,对于全球生态平衡和人类的可持续发展都有重要意义。然而,由于人口的增长和经济的发展,人类活动的范围不断扩大,强度也不断提高,在这过程中引发的环境污染、栖息地丧失和破碎化、对动植物资源的过度开发利用等,使地球上的生物多样性不断减小,大量物种面临严重威胁(陈灵芝等,2001;刘会玉等,2005)。生物多样性的维持和保护需要一个适宜的生态环境,而景观作为生物的栖息地,同时也作为人类的生存环境,景观中斑块的类型、大小、形状、组合、动态等景观特征都会对生物多样性产生影响,因此研究人类活动影响下的景观格局特征对区域生物多样性的影响特征是生物多样性保护的迫切要求。
国内外学者对生物多样性的影响因素进行了较为深入的研究,研究发现影响生物多样性的因素有很多(Gaston et al.,2000),而且比较复杂,但环境因素被认为是影响生物多样性最为主要的因素之一(Harrison et al.,2008)。其中,环境因素根据对生物的影响可以分成生物因子、非生物因子(气候、地形、地貌)和人为因子几种。所以,国内外的研究主要集中在生物因子(Thompson et al.,2005;胡理乐等,2014)、人为因子(Wood et al.,2000;杨婧等,2014)、非生物因子(吴建国等,2009;李巧燕等,2013)或者三者相互共同作用(Kaboli et al.,2006;任学敏等,2012)对生物多样性的影响几方面。随着尺度不同其影响生物多样性的因素也不同(Harrison et al.,2008)。生物因素只在小尺度上成为群落多样性的影响因素,而非生物因素和人为因素则在一系列尺度上都产生影响。大尺度上的非生物因素为生物多样性的形成提供了基础条件,同时在生物因素和人为因素的作用下产生各种景观格局,产生的各种形态、功能、生态过程共同影响区内生物多样性。目前对景观格局与生物多样性的研究尚少,主要集中在,(1)景观破碎化对生物多样性的影响(贺达汉等,2009)。武晶等(2014)研究表明,景观破碎化影响着遗传多样性、物种多样性、生态系统等层次,对种群的遗传多样性、适合度和种群分化有着深刻影响,生境破碎化造成的生境丧失和生境退化是生物多样性丧失的最主要原因之一,改变了生态系统中的能量平衡和能量流动的渠道等方面。(2)景观多样性和景观格局的测度(马克明等,1998)。(3)人类活动对景观多样性的影响(周华锋等,1999)。陈利顶等(1996)以我国黄河三角洲地区东营市为研究区域,选取景观多样性、优势度、景观破碎度和景观分离度作为评价指标,分析了该区人类活动和景观结构之间的关系。研究人类活动的区域差异对景观结构的影响,探讨人类活动的强弱对生物生境和资源分布格局的干扰,成为景观生态学研究的一个重要方面。这些研究基本涵盖了景观格局和生物多样性的多个层次,但有关景观格局与区域生物多样性的相关性研究鲜有报道。
鄱阳湖是中国第一大淡水湖,区内独特的地理环境和气候条件孕育了大量的野生动植物,生物资源丰富,是长江中下游地区重要的生态屏障。然而,近年来,随着鄱阳湖生态经济区工业化、城镇化、农业化的推进,人类活动的强度与频度不断加强,区内的景观发生了很大变化,区内生物多样性受到严重威胁(黄金国等,2007)。因此,探讨鄱阳湖景观特征对生物多样性的影响研究显得尤其重要。本文以鄱阳湖生态经济区为研究区域,试从 38个县区的景观特征中选取具有生态意义的景观指数与生物多样性进行相关分析,探讨景观格局特征对区域生物多样性的影响机理,包括景观格局和空间配置对生物多样性的影响,对科学地保护和管理生物多样性有一定的指导意义。
1 研究区概况
鄱阳湖生态经济区位于江西省北部,包括南昌、景德镇、鹰潭3市,以及九江、新余、抚州、宜春、上饶、吉安市的部分县(市、区),共 38个县(市、区)和鄱阳湖全部湖体在内,面积为5.12×104km2,约占全省国土面积的三分之一(图1)。研究区属于中亚热带温暖湿润气候区,光照充足,雨量充沛,年平均温度为 16.7~17.7 ℃,年均降水量为1400~1900 mm。地形复杂多样,主要以平原和湖泊为主。因其独特的地理环境和气候水文条件,区内生态环境良好,生物资源丰富,特别是鱼类和鸟类,鱼类有139种,占全省鱼类种数的82%,鸟类有310种,占全国鸟类种数的25.41%。区内的社会经济发展良好,2010年生产总值为5478.99亿元,占据全省的60%,人口2039.74万,占全省的 43.51%,非农人口比重上升,2010年区内城市化率为45.1%。
图1 鄱阳湖生态经济区位置Fig. 1 Poyang Lake ecological economic region
2 数据来源及研究方法
2.1 景观分类
本文的景观数据直接来源于 2007年二调鄱阳湖生态经济区各县(市、区)的土地利用图,土地利用类型分类采用两个层次的分类体系,分为 12个Ⅰ级地类和 56个Ⅱ级地类。根据生态系统的分布及动植物的栖息地特点,结合前人对景观分类的研究,对源数据的不同地类进行简化合并,合并后的景观有:林地(FO)、农田(FA)、园地(GA)、湿地(WE)、草地(GR)、建设用地(CO)和其他用地(OT)7大类,如表1所示。
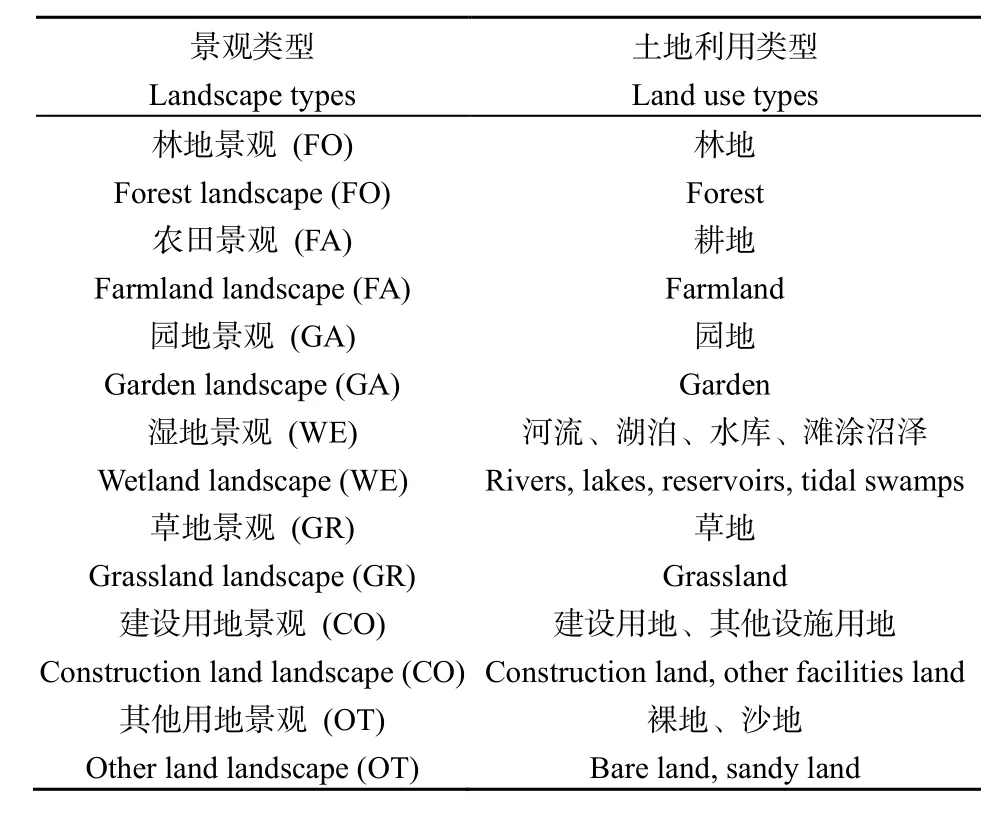
表1 景观类型划分Table 1 Types of landscape
2.2 景观指数的选取
近年来,景观指数在生态学方面的运用越来越广泛和深入,一般将景观格局指数分为斑块水平(patch level)、类型水平(class level)、景观水平(landscape level)3种类型,区域内野生动植物的栖息地主要分布在林地、农田、湿地3类生态系统中,因此,本研究主要分析景观类型水平和景观水平上的格局指数,其中,在景观类型上主要探讨林地、农田和湿地的景观指数,针对面积/密度/边长、形状、聚集/分布、连接性和多样性等5大类型展开研究,选取的指数包括类型水平斑块面积(CA)、斑块占景观面积比例(%)2个,类型及景观水平斑块数量(NP)、斑块密度(PD)、最大斑块占景观面积比例(LPI)、边缘密度(ED)、形状指数(LSI)、面积加权平均分形指数(AWMSI)、景观结合度(CONHESION)7个,景观水平景观面积(TP)、平均斑块面积(MPS)、香农多样性指数(SHDI)、香农均匀度指数(SHEI)4个。
本文在选取景观指数时尽量克服景观指数缺点——出现数据冗余和景观指数存在生态学意义不明确(陈文波等,2002),尽量选取更多的变量进行相关分析,其中部分变量所代表的含义可能有重叠。所以,在SPSS 17.0中利用相关分析剔除相关系数较高的影响因子,减少数据冗余,选取更具有代表性的景观指数,从而更加准确地分析景观格局特征对区域生物多样性的影响因素。
2.3 生物多样性指数的计算及研究方法
本文生物多样性数据来源于2007年江西省“生物多样性调查与评价研究”项目,成果收录在《江西生物多样性调查与评估(2010)》(张海星,2010)中,调查范围包括陆域、水域数据,用生物多样性指数来表示区域的生物多样性状况。其中,生物多样性指数(BI)是物种丰富度、生态系统类型多样性、植被垂直层谱的完整性、生境自然度、物种特有性、外来物种入侵度6个评价指标的加权求和。研究过程中发现野生动物与野生植物的物种数相差较大,应分开计算;植被垂直层谱的完整性指标的区域差异不明显;应增加物种受威胁程度的指标。因此,参照国家环保部发布的中华人民共和国国家环境保护行业标准(HJ623—2011)——《区域生物多样性评价标准》(中华人民共和国国家环境保护部,2011)最终确定了生物多样性指数(BI)由野生维管束植物丰富度、野生高等动物丰富度、生态系统类型多样性、物种特有性、受威胁物种的丰富度、外来物种入侵度6个评价指标进行加权求和。6个评价指标数据均来源于《江西生物多样性调查与评估》(张海星,2010)。
具体计算方法参照国家环保部 2011年发布的中华人民共和国国家环境保护行业标准(HJ623—2011)——《区域生物多样性评价标准》。
生物多样性指数(BI)=归一化的野生动物丰富度×0.2+归一化后的野生维管束植物丰富度×0.2+归一化的生态系统类型多样性×0.2+归一化后的物种特有性×0.2+归一化后的受威胁物种的丰富度×0.1+(100-归一化后的外来物种入侵度)×0.1
本文的景观指数计算在Fragstats 3.3软件中进行,景观指数之间的相关性以及景观指数对生物多样性之间的相关关系运用SPSS 17.0的相关分析方法,采用生物多样性指数表征区域生物多样性状况;同时,为了更好研究景观格局特征对于各物种的影响,对景观指数与物种丰富度进行相关分析。
3 研究结果
3.1 区域生物多样性状况
利用ArcGIS软件对鄱阳湖生态经济区的38个县(市、区)区域生物多样性数据进行空间表达(图2),结果显示,鄱阳湖生态经济区各县(市、区)的生物多样性指数(BI)整体平均值为 40.39,按多样性分级评价划分为中等,说明全区的生物多样性较为丰富。区域内生物多样性在空间分布上差异明显,其中BI值大于30的县有30个,占了全区的78.95%,即物种较丰富,特有属、种较多,生态系统类型较多,局部地区生物多样性高度丰富;最高BI值出现在星子县(64.61),都昌县、永修县、南昌县、庐山区等县BI值也较高,BI值小于30的县区有8个,主要分布在各市辖区,即物种较少,特有属、种不多,局部地区生物多样性较丰富,但是生物多样性总体水平为一般。在空间分布上表现为鄱阳湖周边县市的生物多样性较高,各市辖区以及西南部各县的生物多样性较低,其余县市生物多样性指数处于中等水平。总体上,研究区生物多样性呈现由湖体向四周逐步降低的格局。
3.2 景观指数相关分析
景观类型水平9个指数之间的相关性分析结果见表 2。从表中可以看出,大部分指数之间存在较为显著的相关关系,其中CA、ED、LSI这3个指数与其他指数之间相关关系极为显著,显著相关的比例达到75%,说明大部分指标之间存在较为复杂的关系。数据之间存在冗余,因此将CA、ED、LSI这3个相关系数较高的景观指数剔除,选取剩下的景观类型水平上的6个景观指数进行分析。
同样,景观水平上的 11个景观指数之间的相关性分析结果表明(表 3):景观水平上的景观指数之间存在比景观类型上更高的相关性,大部分指标之间的相关关系显著,只有TA、SHEI与其他指标之间相关性较弱,保持较好的独立性,因此参考景观指数选取的方法,剔除LPI、ED、AWMSI这几个景观指数,在景观水平上最终确定了8个景观指数进行数据分析。
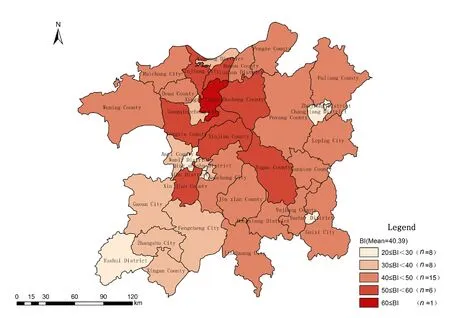
图2 鄱阳湖生态经济区生物多样性空间分布图Fig. 2 The distribution map of biodiversity spatial ecological economic zone of Poyang Lake
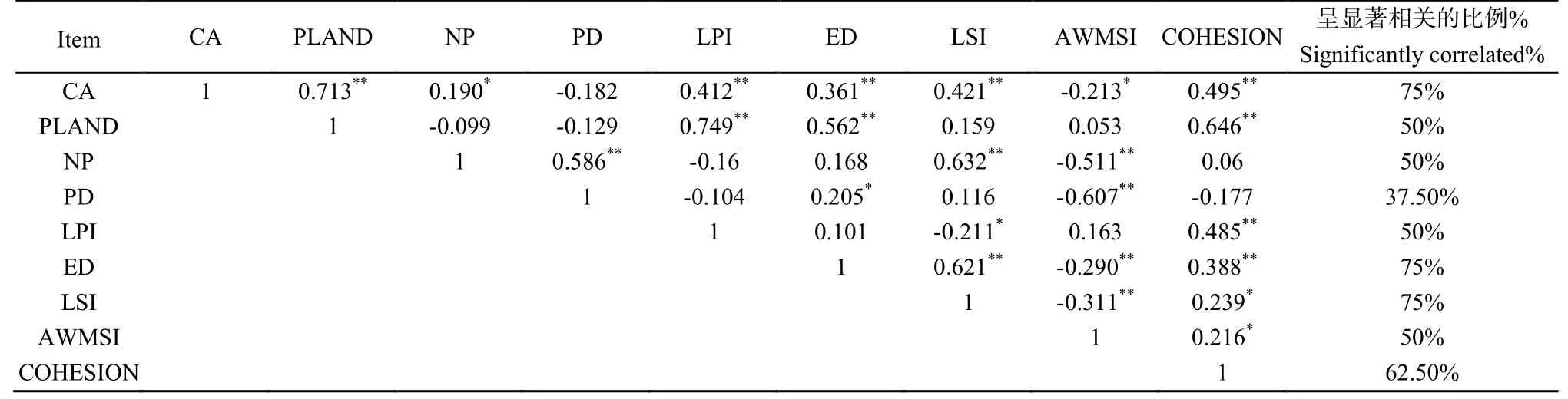
表2 景观类型水平上的9个景观指数相关性分析Table 2 9 landscape landscape types index level of correlation analysis
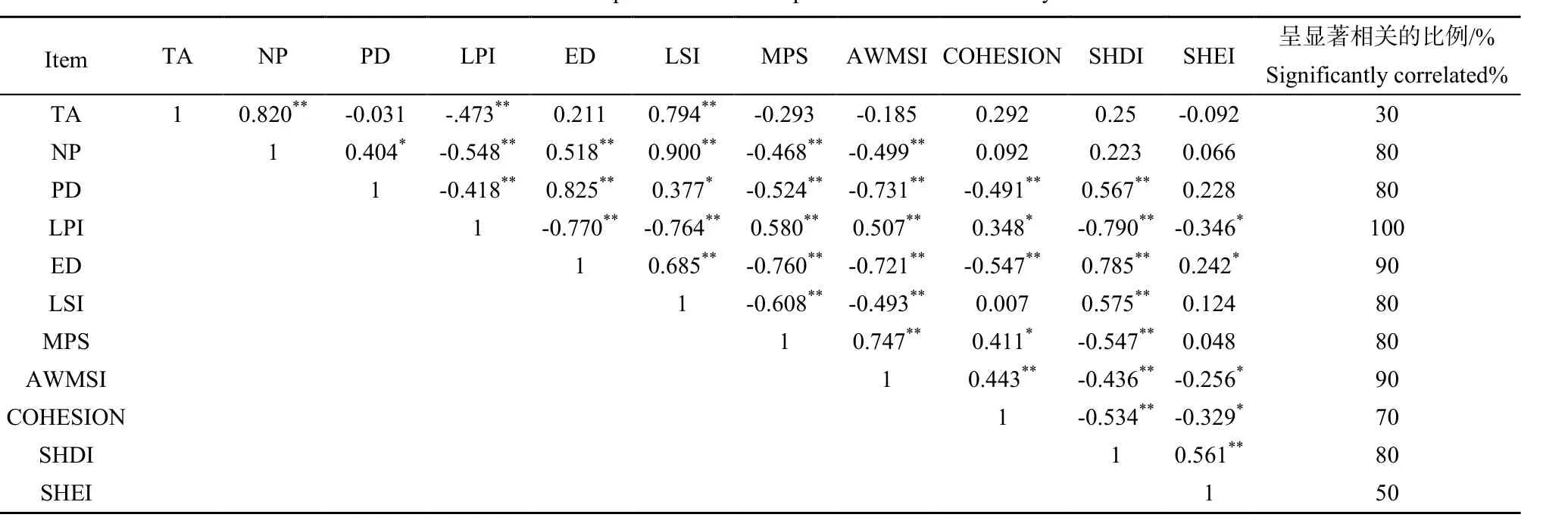
表3 11个景观水平上的景观指数相关性分析Table 3 Landscape level 11 landscape indices correlation analysis
3.3 区域景观格局特征对区域生物多样性的影响
3.3.1 景观类型水平上景观格局特征与生物多样性之间的相关关系
景观类型指数与生物多样性指数之间的相关性分析结果显示(表 4),耕地的 PLAND、NP、COHESION(0.476**,P<0.01)与区域生物多样性指数呈正相关,而与PD、LPI、AWMSI和区域生物多样性指数呈弱程度的负相关。耕地的景观特征与各物种大部分指标之间的相关关系呈不显著的负相关关系,少部分如耕地的斑块数跟各物种之间呈正相关。
区域内林地景观对区域生物多样性的影响也具有一定作用,林地的PLAND(0.439**,P<0.01)、NP、COHESION(0.609**,P<0.01)与区域生物多样性指数呈一般正相关,林地的PD、LPI、AWMSI与区域生物多样性指数呈弱程度的负相关。林地景观除跟植物的 PLAND(0.655**,P<0.01)、COHESION(0.729**,P<0.01)呈极显著的正相关外,林地PLAND、LPI、AWMSI、COHESION与高等动物各物种之间呈现负相关,其中 PLAND、LPI这两个指标与各动物物种之间的关系呈显著负相关,而林地NP、PD与各动物物种之间的关系为弱程度的正相关。
区域内湿地景观对生物多样性的影响具有显著作用,除了与AWMSI呈负相关外,湿地景观大部分指标与区域生物多样性指数呈显著正相关。湿地景观除了与植物物种丰富度呈负相关外,与动物各物种大部分指标的相关关系为显著正相关关系。
3.3.2 景观水平上景观指数与生物多样性之间的相关关系
对整个区域景观水平上的景观指数与生物多样性进行相关性分析,结果如表5所示,区域景观特征除了MPS与区域生物多样性指数呈负相关外,其他景观指标都与区域生物多样性呈正相关,其中TA、NP、LSI、SHDI与区域生物多样性关系较为显著,但是区域景观水平上的景观指数总体上跟鸟类、两栖类、鱼类的相关性较弱。
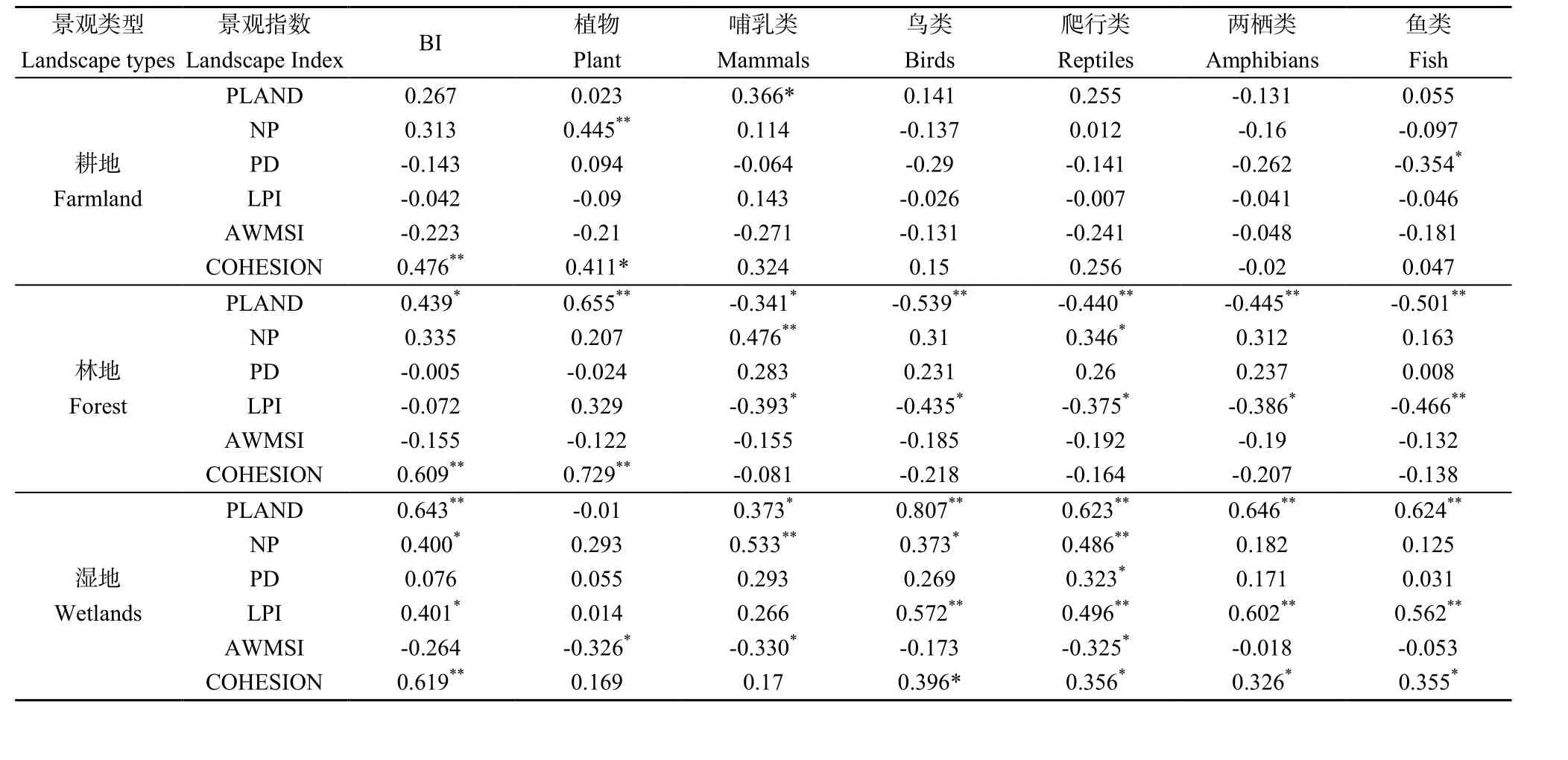
表4 景观类型指数与生物多样性之间的相关性分析结果Table 4 Correlation analysis landscapes and biodiversity index
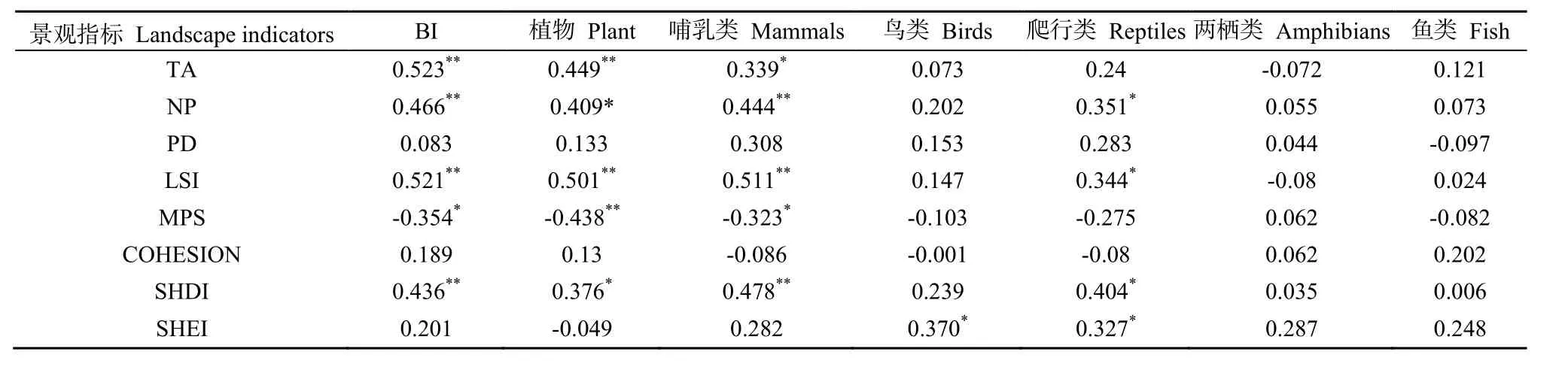
表5 景观水平指数与生物多样性之间的相关分析结果Table 5 Correlation analysis and landscape level biodiversity index
4 讨论与结论
4.1 景观类型水平上景观指数与生物多样性的关系分析
耕地景观与生物多样性相关性分析结果显示,区域内耕地景观对区域生物多样性具有一定的影响。耕地的部分指标与生物多样性指数呈正相关,这是因为耕地作为半自然状态下的人工生态系统,是部分动植物生活的栖息地,对动植物具有一定的维持作用,这在以往研究已得到证实(Tattersall et al.,2002;Benton et al.,2003)。耕地的占比、景观破碎化、景观的结合度都与生物多样性有一定的正相关,即耕地比例越高,破碎化程度越高,结合度越好,区域生物多样性也越丰富。然而,与此同时,耕地作为动植物栖息地的破碎化因子,受人类农业活动的干扰影响大,因此耕地并不是作为动植物栖息的理想之地,这在耕地的景观特征与各物种大部分指标之间的相关关系呈不显著的负相关关系中得到体现。因此,耕地作为一种半人工生态系统,其对动植物的影响是多方面的。
区域林地景观对生物多样性的相关性分析结果表明,林地作为动植物的主要栖息地,对保护生物多样性具有重要作用。林地的比例、景观破碎度、结合度对于区域生物多样性特别是陆域山地生态系统具有重要影响,其中林地景观特征跟植物的相关关系表明,林地是植物的主要栖息地,植物又为动物提供了良好的生长环境。然而,本研究发现区域内林地的景观特征与各物种之间的相关关系并未达到显著水平,且很多呈现负相关,这并不是说明林地对于高等动物不重要,主要是区域内的林地主要分布在海拔较高的丘陵、岗地等地方,而区域内的鸟类、鱼类、两栖类等动物种数丰富的地区主要分布在海拔较低的平原、沼泽和水域等生境。一方面,需保护林地生境中的动植物,另一方面,由于区域内较多的动物分布在林地之外的平原、湿地等生境中,失去了森林等高大植被的覆盖,更容易遭受人类的干扰,所以,在保护湿地的同时,在有条件的地区增加林地、保护灌丛等措施来保护区域的生物多样性。
区域内的湿地景观对于维持区域生物多样性具有重要的作用,其中除了与AWMSI呈负相关外,湿地景观大部分指标与区域生物多样性指数呈显著正相关。说明湿地的面积、优势度、破碎化、连接度与区域生物多样性都呈显著正相关,即湿地的面积越大,优势度越明显、破碎化程度越高、连接度越大,区域生物多样性也越丰富。在探讨湿地景观与物种丰富度之间的关系时,湿地景观与植物物种呈负相关,与动物各物种大部分指标呈显著正相关,特别是湿地的面积与鸟类、爬行类、两栖类、鱼类丰富度呈显著的正相关,说明湿地是动物的主要栖息地,区域内湿地独特的水文、土壤、植被状况,为维持区域生物多样性以及维护区域生态平衡和可持续发展等方面发挥巨大的作用。然而,现实情况是区域内湿地面积由于筑堤围垦不断减少及破碎化,局部湿地受污染比较严重,以及气候变化导致水体面积变化起伏大,这些生态环境问题降低了湿地生态功能的发挥。因此,保护湿地的生态环境,减少人类活动的干扰,这对区域生物多样性的保护有重要作用。
4.2 景观水平上景观指数与生物多样性的关系分析
对整个区域景观水平上的景观指数与生物多样性进行相关性分析,结果表明,景观面积越大,区域景观斑块数目越多,景观形状指数越大,景观的平均面积越小,景观多样性越丰富,区域内生物多样性也越丰富,反映出景观的面积大小、数目、形状、多样性对生物多样性和各种生物过程都有影响。景观面积与生物多样性密切相关,种-面积关系(物种数量和生境面积)是生态学与生物地理学中研究的热点。多数研究表明(诸葛阳等,1986;胡军华等,2007),景观面积是景观内生物多样性的重要决定因素,景观面积越大,生境类型越丰富,生物多样性越丰富。但是,不同的种群(植物、鸟类、两栖类、鱼类)对景观面积的大小有不同的反应(Forman et al.,1981),如景观特征与鸟类、两栖类、鱼类的相关性不显著。这可能因为区域内的鸟类、两栖类、鱼类主要生活在鄱阳湖周边的湿地范围内,水域的生境特征跟陆域的生境有很大不同,导致其在区域尺度上的相关关系不显著,可以缩小研究尺度进行区内湿地的景观特征对生物多样性的影响研究(施建敏等,2010)。因为研究区域是面积大小不一样的县(市、区),而斑块数量跟面积具有很大的相关性,景观斑块数量越多,景观和区域生物多样性就越高。单位面积的景观数量越多,景观的破碎化越严重。景观面积的减少和斑块片段化,不利于物种的散布和迁移。
景观形状同样对各种生物的扩散和动物的觅食等生态过程和各种功能流产生了重要影响,其对区域生物多样性的影响也是极其复杂的。如圆形斑块内部面积大,物种丰富,种群数量也大,但是与相邻和远距离基质间的交流较少(Diamond,1975),而狭长状的景观类型与周边相邻斑块交流较多,同时也容易受到外界的干扰(Forman et al.,1992)。整体上,景观形状的复杂使得边缘效应增强。许多研究表明,斑块边界部分常常具有较高的物种丰富度和初级生产力(Laurance et al.,1991;Didham et al.,1998)。
景观多样性与区域生物多样呈显著的正相关,跟植物、哺乳类、爬行类之间的相关关系也较密切,呈正相关关系,说明景观多样性对区域生物多样性的影响较为显著。相关研究表明,景观多样性与生态学中的物种多样性有紧密的联系,但并不是简单的正比关系,在同一景观中二者的关系一般呈正态分布(傅伯杰等,1996)。
4.3 结论
本文通过对区域不同水平上的景观指数与生物多样性进行相关分析,探讨研究区域景观特征对生物多样性的影响机理,得到以下结论:
(1)景观类型水平上景观指数与生物多样性之间的相关性分析结果表明,耕地部分指标与生物多样性指数呈正相关,耕地的景观特征与各物种大部分指标之间呈不显著的负相关,表现出耕地景观与生物多样性关系复杂。区域内林地的景观特征与各物种之间的相关关系并不是很显著,很多呈现负相关。而湿地景观大部分指标与区域生物多样性指数呈现显著的正相关,与动物各物种特别是与鸟类、爬行类、两栖类、鱼类呈显著的正相关。说明区域内的湿地景观、林地景观是区域生物多样性的主要栖息场所,同时作为产粮主产区,耕地景观的影响也不容忽视。因此保护区域内的湿地、林地、耕地景观对于维持区域生物多样性有重要的影响。
(2)景观水平上景观指数与生物多样性之间的相关性分析结果表明:景观面积大小、数量、形状、多样性对生物多样性和各种生物过程都有较大影响。区域景观特征与生物多样性具有较好的相关性,关系较为复杂。然而,也有景观特征与物种种群如鸟类、两栖类、鱼类关系之间呈现弱相关,相关关系较为复杂。
AUNDERS D A, HOBBS R J, MARGULES C R. 1991. Biological consequences of ecosystem fragmentation: A review [J]. Conservation Biology, 5(1): 18-32.
BENTON T G, VICKERY J A, WILSON J D. 2003. Farmland biodiversity:is habitat heterogeneity the key? [J]. Trends in Ecology & Evolution,18(4): 182-188.
DIAMOND J M. 1975. The island dilemma: lessons of modern biogeographic studies in space and time [J]. Landscape Ecology, 2(1):23-44.
DIDHAM R K, HAMMOND P M. 1998. Beetle species responses to tropical forest fragmentation [J]. Ecological Monographs, 68(3):295-323.
FORMAN R T T, GODRON M. 1981. Patches and structural components for a landscape ecology [J]. Bioscience, 31(10): 733-740.
FORMAN R T T, MOORE P N. 1992. Theoretical foundations for understanding boundaries in landscape mosaics [M]//In: Hansen A J, di Castri F.Landscape Boundaries: Consequence for Biotic diversity and Ecoloical Flows. New York: Springer-Verlag: 236-258.
GASTON K J, BLACKBURN T M. 2000. Pattern and process in macroecology [M]. London: Blackwell Press.
HARRISON S, CORNELL H. 2008. Toward a better understanding of the regional causes of local community richness [J]. Ecology Letters,11(9): 969-979.
KABOLI M, GUILLAUMET A, PRODON R. 2006. Avifaunal gradients in two arid zones of central Iran in relation to vegetation, climate, and topography [J]. Journal of Biogeography, 33(1): 133-144.
LAURANCE W F, YENSEN E. 1991. Predicting the impacts of edge effects in fraagmented habitats [J]. Biological Conservation, 55(1): 77-92.
TATTERSALL F H, MACDONALD D W, HART B J, et al. 2002. Is habitat linearity important for small mammal communities on farmland? [J].Journal of Applied Ecology, 39(4): 643-652.
THOMPSON W A, ELDRIDGE D J. 2005. Plant cover and composition in relation to density of Callitris glaucophylla ( white cypress pine) along a rainfall gradient in eastern Australia [J]. Australian Journal of Botany,53(6): 545-554.
WOOD A, STEDMAN-EDWARDS P, MANG J. 2000. The Root Cause of Biodiversity Loss [M]. London: UK. Earthscan Publications Ltd.
陈利顶, 傅伯杰. 1996. 黄河三角洲地区人类活动对景观结构的影响分析——以山东省东营市为例[J]. 生态学报, 16(4): 337-344.
陈灵芝, 马克平. 2001. 生物多样性科学: 原理与实践[M]. 上海: 上海科学技术出版社.
陈文波, 肖笃宁, 李秀珍. 2002. 景观指数分类、应用及构建研究[J]. 应用生态学报, 13(1): 121-125.
傅伯杰, 陈利顶. 1996. 景观多样性的类型及其生态意义[J]. 地理学报,51(5): 454-462.
贺达汉, 赵紫华, 张大治. 2009. 草原景观下昆虫群落及种群对生境破碎化的反应[J]. 草业学报, 18(6): 235-241.
胡军华, 胡慧建, 蒋志刚. 2007. 大空间尺度上物种多样性的分布规律[J]. 应用与环境生物学报, 13(5): 731-735.
胡理乐, 李俊生, 罗建武, 等. 2014. 生物质能源植物种植对生物多样性的影响[J]. 生物多样性, 22(2): 231-241.
黄金国, 郭志永. 2007. 鄱阳湖湿地生物多样性及其保护对策[J]. 水土保持研究, 14(1): 305-306, 309.
李巧燕, 王襄平. 2013. 长江三峡库区物种多样性的垂直分布格局: 气候、几何限制、面积及地形异质性的影响[J]. 生物多样性, 21(2):141-152.
刘会玉, 林振山, 张明阳. 2005. 人类周期性活动对物种多样性的影响及其预测[J]. 生态学报, 35(7): 1635-1641.
马克明, 傅伯杰, 周华峰. 1998. 景观多样性测度: 格局多样性的亲和度分析[J]. 生态学报, 18(1): 78-83.
任学敏, 杨改河, 王得祥, 等. 2012. 环境因子对巴山冷杉-糙皮桦混交林物种分布及多样性的影响[J]. 生态学报, 32(2): 605-613.
施建敏, 马克明, 赵景柱, 等. 2010. 三江平原残存湿地斑块特征及其对物种多样性的影响[J]. 生态学报, 30(24): 6683-6690.
吴建国, 吕佳佳, 艾丽. 2009. 气候变化对生物多样性的影响: 脆弱性和适应[J]. 生态环境学报, 18(2): 693-703.
武晶, 刘志民. 2014. 生境破碎化对生物多样性的影响研究综述[J]. 生态学杂志, 33(7): 1946-1952.
杨婧, 褚鹏飞, 陈迪马, 等. 2014. 放牧对内蒙古典型草原α、β和γ多样性的影响机制[J]. 植物生态学报, 38(2): 188-200.
张海星. 2010. 江西生物多样性调查与评估[M]. 南昌: 江西科学技术出版社: 6.
中华人民共和国国家环境保护部. 2011. 区域生物多样性评价标准[S].北京: 中国环境科学出版社.
周华锋, 马克明, 傅伯杰. 1999. 人类活动对北京东灵山地区景观格局影响分析[J]. 自然资源学报, 14(2): 22-27.
诸葛阳, 姜仕仁, 郑忠伟, 等. 1986. 浙江海岛鸟兽地理生态学的初步研究[J]. 动物学报, 32(1): 74-86.
Study on the Relationship between Landscape Pattern and Regional Biodiversity
LIN Shitao1,2,3, XIE Dibing4, LIU Yulin3, CHEN Wenbo2,3*
1. Jiangxi Environmental Engineering Vocational College, Ganzhou 341000, China;
2. Research Center of Environment and Landscape Ecology, Jiangxi Agriculture University, Nanchang 330045, China;
3. Key Laboratory of Environment and Landscape, Nanchang 330045, China; 4. Jiangxi College of Applied Technology, Ganzhou 341000, China
Landscape pattern determines the function of landscape and ecological process, affecting the landscape within the species flow, information flow, energy flow, which has a significant effect on biodiversity. In this paper, choosing the ecological economic zone of Poyang Lake as the study area, SPSS 17.0 and Fragstats 3.3 techniques were selected with the ecological significance of landscape and biodiversity index from 38 counties in the landscape characteristics analysis, in order to explore the relationship between landscape pattern and regional biodiversity. The results showed that: (1) The relationship between the level of landscape types and landscape indices of biodiversity varies, some indicators, such as PLAND, COHESION land NP (0.476**, P<0.01), was positively correlated with the regional biodiversity index, the relationship between landscape characteristics of cultivated land and the species most negative, showing the farmland landscape for the complex relationship between biodiversity. The PLAND (0.439**,P<0.01), NP, COHESION (0.609**, P<0.01) in the region were generally positively correlated with regional biodiversity index, and the PD, LPI and AWMSI of forest land were negatively correlated with regional biodiversity index. While most of the indicators of wetland landscape and regional biodiversity index showed significant positive correlation. (2) The landscape level between the landscape index and biodiversity related results showed that the index of regional landscape features in addition to MPS and regional biodiversity was negatively correlated, other landscape indicators were associated with regional biodiversity positively, and among them, that of TA, NP, LSI, and SHDI with regional species diversity was significant, indicating the size of landscape, number, shape,and diversity had a great influence on the biodiversity and various biological processes. The wetland landscape area and forest landscape were the main habitat of regional biodiversity, and at the same time as the grain producing areas, farmland landscape effects could not be ignored. Therefore, to protect the area of wetland, woodland and farmland landscape has important implications for maintaining regional biodiversity. The research has certain theoretical guiding significance for regional biodiversity protection and management.
landscape pattern; biodiversity; Poyang Lake Eco-economic Region
10.16258/j.cnki.1674-5906.2017.10.006
X176
A
1674-5906(2017)10-1681-08
林世滔, 谢弟炳, 刘郁林, 陈文波. 2017. 景观格局特征与区域生物多样性的关系研究[J]. 生态环境学报, 26(10):1681-1688.
LIN Shitao, XIE Dibing, LIU Yulin, CHEN Wenbo. 2017. Study on the relationship between landscape pattern and regional biodiversity [J]. Ecology and Environmental Sciences, 26(10): 1681-1688.
国家自然科学基金项目(41161031;41561043);江西省教育厅科学技术研究项目(GJJ161371)
林世滔(1989年生),男,助教,硕士,研究方向为自然资源管理、林业3S技术。E-mail: linshitaofighting@163.com
*通信作者:陈文波(1974年生),男,教授,博士生导师,从事自然资源管理、景观生态学研究。E-mail: cwb1974@126.com
2015-11-17
猜你喜欢
英语世界(2023年10期)2023-11-17 09:18:18
现代装饰(2021年6期)2021-12-31 05:27:54
小学科学(学生版)(2020年12期)2021-01-08 09:28:10
少年漫画(艺术创想)(2020年12期)2020-06-09 05:50:08
科学大众(中学)(2019年3期)2019-05-17 10:04:30
汽车观察(2018年10期)2018-11-06 07:05:26
辽宁林业科技(2017年4期)2017-06-22 10:23:36
河北林业科技(2016年5期)2016-11-08 03:13:26
少儿科学周刊·少年版(2015年1期)2015-07-07 17:15:12
中国摄影家(2014年6期)2014-04-29 14:54:47
