
遮光和施氮对暗紫贝母生长和C-N平衡的影响
2017-11-10 01:47郭海霞石福孙
植物研究 2017年5期
郭海霞 徐 波 石福孙 吴 彦*
(1.成都师范学院,成都 611130; 2.中国科学院成都生物研究所生态恢复与生物多样性保育四川省重点实验室,成都 610041; 3.阿坝师范学院,汶川 623002)
遮光和施氮对暗紫贝母生长和C-N平衡的影响
郭海霞1徐 波3石福孙2吴 彦2*
(1.成都师范学院,成都 611130;2.中国科学院成都生物研究所生态恢复与生物多样性保育四川省重点实验室,成都 610041;3.阿坝师范学院,汶川 623002)
为了了解暗紫贝母对资源变化的适应策略,本研究通过田间控制试验,研究了遮光和施氮对暗紫贝母生物量、生物量分配,以及叶、根C、N状态的影响。结果表明:遮光和施氮处理后,暗紫贝母生物量均减少;遮光下叶片的生物量分配相对增加,叶根生物量比显著升高(P<0.05),但施氮处理下叶根生物量比的变化不明显;遮光和施氮后,暗紫贝母叶、根N含量均显著升高(P<0.05),叶片和根的C:N显著降低(P<0.05);相关分析显示,叶片N、C∶N与暗紫贝母总生物量、叶根生物量比有极显著的相关性(P<0.01)。本研究表明,与生物量分配相比,C-N平衡,尤其是叶片的C∶N比能更好的解释暗紫贝母对资源变化的适应。
暗紫贝母;遮光;施氮;生物量分配;C-N平衡
研究者很早便发现,植物有维持生长平衡的倾向。由于地上地下资源的不平衡,陆生植物需要调整各个器官的生物量分配,尤其是根叶生物量分配,以维持资源吸收的平衡,实现植物的最优生长,这一理论被称为“功能平衡理论”[1]。根据功能平衡理论,地上资源不足时,向叶片分配的生物量比例增加,而地下资源不足时,向根分配的生物量比例增加。例如,Grechi[2]等的研究发现,光照不足时,葡萄的叶根生物量比例增加,N供应不足时,叶根生物量比例降低。然而,研究者发现,植物并不总是依靠生物量分配的调整来适应环境资源的变化。例如,Poorter[3]等总结发现,对比生物量分配与其他形态指标,植物更多的是通过形态的调整来适应资源的变化,而不是生物量分配。此外,“功能平衡理论”也无法解释这些现象:K、Mg和Mn不足时,根叶生物量比反而降低;CO2浓度升高对植物的生物量分配无影响[4]。
“C-N平衡”理论是由“功能平衡理论”演化而来的一个理论[5]。自从Liebig揭示营养元素对作物生长的影响以来,人们就认识到了N元素对于植物的重要意义,并开展了一系列研究。而随着这些研究的进行,人们发现了另一个重要的问题,C和N元素之间的协同作用对植物生长和环境适应的重要意义。植物主要依赖叶片获得C元素,依赖根系获得N元素。但是,叶片对C元素的获取能力与光合作用的各种酶紧密相光,而这些酶的合成又依赖于来自根系的N元素。同样,根系对N元素的吸收也离不开来自叶片的C元素[6]。这就暗示,植物要维持正常生长,就需要体内的C、N保持一定的平衡。模型研究证实,植物可根据环境状况,调整叶片和根系之间的C、N流通,适应资源的变化。而叶片和根系之间的C、N流通也决定了叶片和根的生物量分配[7~8]。研究者将这一现象称作协调理论(coordination theory)或C-N平衡理论[9~10]。Ericsson发现,尽管N不足时,K、Mg和Mn不足时,以及CO2浓度升时,这些情况下,植物生物量分配的响应不相同,但是都由叶片和根系内的C-N平衡决定的。由此可见,C-N平衡理论或许能更好的解释植物对资源变化的适应。
暗紫贝母(FritillariaunibracteataHsiao et K.C.Hsia)是药材“川贝母”之一“松贝”的主要基原植物,为“川贝母”中的上品,由于资源的过度开发,早在1987年便被列入了《国家重点保护野生药材名录》[11]。近年来,随着人工采挖的增加,以及生境的变化,暗紫贝母的野生资源面临着更大的威胁。植物的环境适应策略是决定其能否适应环境变化的重要因素。但是,由于该物种生长于海拔3 200~4 300 m的高海拔地区,自然条件恶劣,少有研究者对它的适应策略进行探讨。光和N元素是植物生长所需最重要的地上和地下资源,也是影响暗紫贝母生长的重要因子。高海拔地区土壤温度低,土壤对植物的N供应速率慢,根系对养分的吸收能力也弱,因此植物往往受到N的限制[12~13]。此外,暗紫贝母生长于高海拔草甸和灌丛中,光照条件也可能是影响它生长的重要因子[14~15]。因此,本试验将通过研究遮光和N添加下暗紫贝母生物量分配和C-N平衡的调整,探讨它对资源变化的适应策略,为其野生资源的保护及开发提供一定的理论依据。
1 材料与方法
1.1 试验设置
试验基地位于阿坝州松潘县(32°56′18″N,103°42′45″E,海拔3 300 m)。该地年均温2.7℃,最低月均温-7.6℃,最高月均温11.7℃。年均降水717.7 mm,且80%集中在5~10月。
试验设置于2013年5月初。首先从大田里取土,过筛后混合。混合后的土壤含有机质96.04 g·kg-1,总N 3.60 g·kg-1,总P 0.83 g·kg-1,总K 21.29 g·kg-1。取混合土壤,装于塑料盆中备用(长×宽×高=0.45 m×0.35 m×0.2 m)。然后,从田间选取植株大小相似的三年生暗紫贝母,移栽到塑料盆中。每盆12株,每株间距10 cm。
试验采用完全随机设计。设置对照(CK)、施氮(NA)和遮光(SH)3组,每组各包括45盆植株。遮光处理:遮光处理利用遮阳网实现。首先利用木料和钢丝,搭建一个高0.75 m的棚,然后将遮阳网铺设固定在棚上方。根据前期的试验和文献调研[15],我们推断70%的遮光强度会对暗紫贝母的生长产生明显的抑制。因此,本研究中将遮光强度设置为70%。施氮处理:利用硝酸铵(NH4NO3)进行氮添加。郑军[16]对川贝母的研究发现,随着N添加的增加,鳞茎产量逐渐降低,在40 g·m-2时降到最低;总生物碱则逐渐增加,在40 g·m-2时最高;而可溶性淀粉和可溶性蛋白质均在40 g·m-2施氮处理下最低。因此,本研究选择40 g·m-2作为施氮量。试验期间共进行两次施氮处理,分别在移栽后一周和第二年春天土壤解冻时。
1.2 样品采集与处理
于第二年生长季节旺期(2014年6月,暗紫贝母开花盛期),进行样品采集。从每组的45个塑料盆中,随机抽取50株植物进行采样。由于抽取的株数大于盆数,因此将盆数乘以2作为(即90)样本数,1~45和46~90分别对应塑料盆1~45。利用SPSS 16.0的抽样功能从90个样本抽取50个作为被采样的塑料盆。与之相似,将每盆中的植株进行编号,利用SPSS 16.0的抽样功能确定采样盆中的采样株。植株采集后,迅速清洗干净,分成叶片、茎、鳞茎、根4部分。然后,将各部分置于烘箱内,于75℃下烘干至恒重后,进行称重。称重后将每种处理方式下的叶片和根,分别每10株做为一份,磨碎,利用TOC/TN分析仪(Milti N/C2100S,Jena,Germany),测定其总C、总N含量。
1.3 数据分析
数据分析和图片处理主要利用SPSS 16.0(Statistical Package for Social Sciences program 16.0)和Origin 9.0进行,测定结果用“平均值±标准误”表示。使用one-way ANOVA分析和LSD比较不同处理间的差异,利用Pearson相关性分析分析暗紫贝母C、N状态与总生物量、生物量分配之间的相关性。
2 试验结果
2.1 遮光和施氮对生物量和生物量分配的影响
试验结果显示,在不施加处理的情况下,暗紫贝母的生物量随着生长发育的进行逐渐增加,而长时间的遮光和施氮对生物量的累积过程,产生了明显的抑制。从图1可以看出,随着施氮的进行,到最后一次采样时,施氮处理下的总生物量(TB)和鳞茎生物量(BB)都显著低于对照(P<0.05);与之相似,遮光处理下,最后一次采样下的TB、叶片生物量(LB)、根生物量(RB)和BB也显著的低于对照(P<0.05)。
总体来看,随着生长发育的进行,暗紫贝母的生物量分配格局发生了一定程度的变化。叶根生物量比(LB/RB)在整个试验期间无明显变化;地上地下生物量比(AB/BeB)和根生物量/总生物量(RB/TB)呈现出降低趋势;叶片生物量/总生物量(LB/TB)则仅在第一个生长季内表现出明显的降低,第二个生长季节末与第一个生长季节末之间则无显著差异(图2)。施氮和遮光对暗紫贝母生物量分配格局产生了不同的影响。施氮处理并没有改变暗紫贝母生物量格局的变化趋势,各个采样时间下,其生物量分配与对照均无显著差异(P>0.05)。而遮光处理对生物量分配格局的影响则很明显,在最后一次采样时,LB/RB、AB/BeB和LB/TB均显著高于对照(P<0.05)。

表1 遮光和施氮对暗紫贝母C、N的影响(平均值±标准误)
注:小写字母表示同一处理下不同采样时间之间的差异(P<0.05);大写字母表示同一采样时间下不同处理方式之间的差异(P<0.05)。
Note:Lower case letters indicate the difference between different sampling time under the same treatment(P<0.05);Upper case letters indicate the difference between different treatments on the same sampling day(P<0.05).
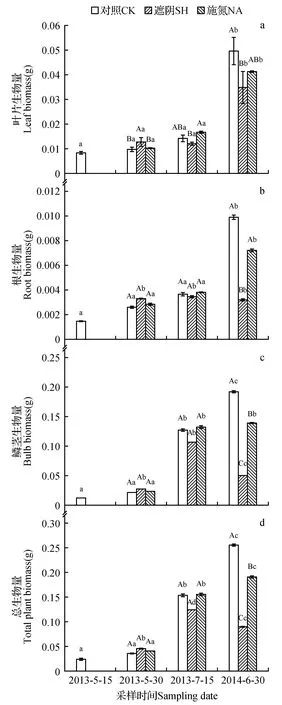
图1 遮光和施氮对暗紫贝母叶片生物量(a)、根生物量(b)、鳞茎生物量(c)和总生物量(d)的影响 小写字母表示同一处理下,不同采样时间的差异(P<0.05);大写字母表示同一采样时间下不同处理方式的差异(P<0.05) 下同。Fig.1 Effects of shading and nitrogen addition on leaf biomass(a),root biomass(b),bulb biomass(c) and total plant biomass(d) of F.unibracteata Lower case letters indicate the difference between different sampling time under the same treatment(P<0.05);Upper case letters indicate the difference between different treatments on the same sampling day(P<0.05) The same as below.
2.2 遮光和施氮对叶片和根C、N的影响
施氮和遮光处理下,与对照相比,暗紫贝母的C、N状态均发生了一定改变,并且随着生长发育的进行,遮光和施氮处理的影响逐渐明显。更重要的是,数据显示,遮光和施氮对暗紫贝母C、N表现出相似的影响。从表1可以看出,在最后一次采样中,遮光和施氮处理下,叶片N(LN)、根N(RN)均显著升高(P<0.05),根C(RC)、叶片CN比(LC/LN)、根CN比(RC/RN)和叶根N比(LN/RN)均显著降低(P<0.05)。但是,叶片C(LC)在遮光处理下显著降低(P<0.05),在施氮处理下变化不显著;而叶根C比(LC/RC)在施氮处理下显著升高(P<0.0.05),在遮光处理下变化不显著。
2.3 生物量及其分配与C、N状态的相关性
从表2可知,暗紫贝母叶、根的C、N含量及比例与暗紫贝母的生物量和生物量分配存在不同的联系。总生物量(TB)与LN、RN和LC/RC有极显著的负相关(P<0.01),而与RC、LC/LN和RC/RN呈极显著正相关(P<0.01)。LB/RB与LC、LC/LN和LC/RC极显著负相关(P<0.01),而与LN呈极显著正相关(P<0.01)。
表2暗紫贝母C、N状态与其生物量的相关性(n=36)
Table2CorrelationsamongstatusofC,Nandtotalplantbiomassandbiomassallocation(n=36)

指标IndexLCLNRCRNLC/LNRC/RNLC/RCLN/RNTB0.007-0.539∗∗0.530∗∗-0.476∗∗0.529∗∗0.609∗∗-0.430∗∗0.17LB/RB-0.414∗0.391∗0.0690.292-0.444∗∗-0.214-0.426∗∗0.081
*P<0.05;**P<0.01
3 讨论
3.1 遮光和施氮对暗紫贝母C-N平衡的影响
研究指出,高山植物的N含量一般高于全球植物的平均N含量。全球植物叶片N均值约为20.1 mg·g-1,根系N均值约为9.8 mg·g-1[17]。本研究中,暗紫贝母的叶片N和根N含量分别为53.4和36.2 mg·g-1,远高于全球平均值。遮光和施氮处理后,暗紫贝母根、叶的N含量进一步升高,根、叶的C∶N均明显降低。Dickman[18]等认为,植物的C∶N决定于光照与可利用N之间的比率。本研究中,栽培使用的混合土,总N含量3.60 g·kg-1,含N量较高。施氮后,N得到进一步补充,此时,光照成为相对受限的资源,其可利用比率较低。同样,遮光处理下,光照的可利用比率也较低。这可能正是遮光和施氮下C∶N均表现为降低的原因。
3.2 遮光和施氮对暗紫贝母生物量的影响
光照和氮元素是影响植物生长的两个关键因子。光照代表植物生长所需的地上资源,而氮元素代表地下资源。本研究中,长期的遮光和施氮下,暗紫贝母的生长受到了抑制。暗紫贝母是一种多年生草本植物,生长在草甸和灌丛中。适当的遮光有利于暗紫贝母的生长[14]。然而,本研究中,70%的遮光明显不利于三年生暗紫贝母。这与Li[14]等对川贝母的研究结果不同。他发现50%的遮光处理下,川贝母净同化率增加。这可能是因为,遮光既能降低植物的光合产量,也能减少植物的呼吸消耗。当遮光下光合作用的降低程度小于呼吸消耗降低的程度时,遮光有利于植物的生长,而当光合作用的降低程度大于呼吸消耗降低的程度时,遮光便会抑制植物的生长[19~20]。此外,植物实现最优生长,需要一定的C∶N。高山植物的C∶N含量一般较低,遮光和施氮后,这一比值进一步降低,加剧了C、N之间的不平衡,由此影响了植物的生长[21~22]。这也可能是遮光和施氮后,暗紫贝母生长均受到抑制的原因。
3.3 遮光和施氮对暗紫贝母生物量分配的影响
本研究中,遮光下,暗紫贝母的叶根生物量比极显著升高,而施氮下的变化则不明显。其原因可能是,首先,生物量分配不是植物调整资源吸收的唯一方式,叶面积,比根长等的调整均可改变植物对资源的吸收[3,23]。此外,生物量分配的影响因素也不只是环境因子。研究指出,随着植物生长的进行,生物量分配也会发生改变[24~26]。尽管生物量分配的变化在遮光和施氮处理后表现出一定的差异,但相关分析发现,暗紫贝母的C、N状态,尤其是叶片的N和C∶N与暗紫贝母总生物量和生物量分配均有极显著的相关性,这暗示了暗紫贝母C-N平衡与生物量分配、资源吸收策略之间的联系。此外,有研究指出,C∶N与植物生物碱的合成与积累相关,而生物碱是“川贝母”主要的药效成分[11,27]。由此可见,C∶N是研究暗紫贝母的一个关键指标。
4 结论
本研究中,遮光和施氮后,暗紫贝母叶、根N含量均显著升高(P<0.05),叶片和根C∶N显著降低(P<0.05)。而叶根生物量比在遮光下极显著升高,在施氮处理下变化不明显。相关分析显示,暗紫贝母的C、N状态,尤其是叶片的N和C∶N,与其生长及生物量分配有显著的相关性。这表明,与生物量分配相比,暗紫贝母的C-N平衡,尤其是叶片的C-N平衡,或许能更好的解释暗紫贝母对资源变化的适应策略。
1.Brouwer R.Nutritive influences on the distribution of dry matter in the plant[J].Journal of Agricultural Science,1962,10:399-408.
2.Grechi I,Vivin P,Hilbert G,et al.Effect of light and nitrogen supply on internal C∶N balance and control of root-to-shoot biomass allocation in grapevine[J].Environmental and Experimental Botany,2007,59(2):139-149.
3.Poorter H,Niklas K J,Reich P B,et al.Biomass allocation to leaves,stems and roots:meta-analyses of interspecific variation and environmental control[J].New Phytologist,2012,193(1):30-50.
4.Ericsson T.Growth and shoot:root ratio of seedlings in relation to nutrient availability[J].Plant and Soil,1995,168-169(1):205-214.
5.平晓燕,周广胜,孙敬松.植物光合产物分配及其影响因子研究进展[J].植物生态学报,2010,34(1):100-111.
Ping X Y,Zhou G S,Sun J S.Advances in the study of photosynthate allocation and its controls[J].Chinese Journal of Plant Ecology,2010,34(1):100-111.
6.Baptist F,Aranjuelo I.Interaction of carbon and nitrogen metabolisms in alpine plants[M].//Lütz C.Plants in Alpine Regions.Vienna:Springer,2012:121-134.
7.Thornley J H M.A balanced quantitative model for root:shoot ratios in vegetative plants[J].Annals of Botany,1972,36(2):431-441.
8.Reynolds J F,Thornley J H M.A shoot:root partitioning model[J].Annals of Botany,1982,49(5):585-597.
9.Osaki M.Carbon-nitrogen interaction model in field crop production[J].Plant and Soil,1993,155-156(1):203-206.
10.Reynolds H L,D’antonio C.The ecological significance of plasticity in root weight ratio in response to nitrogen:opinion[J].Plant and Soil,1996,185(1):75-97.
11.韩鸿萍,陈志.暗紫贝母研究现状[J].青海师范大学学报:自然科学版,2016(1):29-33,89.
Han H P,Chen Z.Research progress ofFritillariaunibracteataHsiao et K.C.Hisa[J].Journal of Qinghai Normal University:Natural Science,2016(1):29-33,89.
12.Körner C.高山植物功能生态学[M].吴宁,罗鹏,译.北京:科学出版社,2009:127.
Körner C.Alpine plant life:functional plant ecology of high mountain ecosystems[M].Wu N,Luo P,trans.Peking:Science Press,2009:127.
13.Sebastian N,Erika H,Christian K.Critically low soil temperatures for root growth and root morphology in three alpine plant species[J].Alpine Botany,2016,126(1):11-21.
14.Li X W,Dai Y,Chen S L.Growth and physiological characteristics ofFritillariacirrhosain response to high irradiance and shade in age-related growth phases[J].Environmental and Experimental Botany,2009,67(1):77-83.
15.徐波,王金牛,石福孙,等.青藏高原东缘野生暗紫贝母生物量分配格局对高山生态环境的适应[J].植物生态学报,2013,37(3):187-196.
Xu B,Wang J N,Shi F S,et al.Adaptation of biomass allocation patterns of wildFritillariaunibracteatato alpine environment in the eastern Qinghai-Xizang Plateau[J].Chinese Journal of Plant Ecology,2013,37(3):187-196.
16.郑军.人工栽培濒危药用植物川贝母鳞茎质量研究[D].雅安:四川农业大学,2009.
Zheng J.Study on the quality of the bulb of cultivationFritillariacirrhosa[D].Ya’an:Sichuan Agricultural University,2009.
17.Hong J T,Wang X D,Wu J B.Stoichiometry of root and leaf nitrogen and phosphorus in a dry alpine steppe on the northern Tibetan plateau[J].PLoS One,2014,9(10):e109052.
18.Dickman E M,Vanni M J,Horgan M J.Interactive effects of light and nutrients on phytoplankton stoichiometry[J].Oecologia,2006,149(4):676-689.
19.Lehto T,Grace J.Carbon balance of tropical tree seedlings:a comparison of two species[J].New Phytologist,1994,127(3):455-463.
20.Sims L,Pastor J,Lee T,et al.Nitrogen,phosphorus and light effects on growth and allocation of biomass and nutrients in wild rice[J].Oecologia,2012,170(1):65-76.
21.Körner C.The nutritional status of plants from high altitudes[J].Oecologia,1989,81(3):379-391.
22.Weih M,Karlsson P S.Growth response of Mountain birch to air and soil temperature:is increasing leaf-nitrogen content an acclimation to lower air temperature?[J].New Phytologist,2001,150(1):147-155.
23.Hodge A.The plastic plant:root responses to heterogeneous supplies of nutrients[J].New Phytologist,2004,162(1):9-24.
24.Wilson J B.A review of evidence on the control of shoot:root ratio,in relation to models[J].Annals of Botany,1988,61(4):433-449.
25.Gedroc J J,Mcconnaughay K D M,Coleman J S.Plasticity in root/shoot partitioning:optimal,ontogenetic,or both?[J].Functional Ecology,1996,10(1):44-50.
26.Mcconnaughay K D M,Coleman J S.Biomass allocation in plants:ontogeny or optimality? A test along three resource gradients[J].Ecology,1999,80(8):2581-2593.
27.佟璐,张宝友,唐中华,等.弱光胁迫对大田长春花生物量分配及次生代谢的影响[J].植物研究,2011,31(2):227-230.
Tong L,Zhang B Y,Tang Z H,et al.Effects of low light stress on the biomass distribution and secondary metabolism ofCatharanthusroseus[J].Bulletin of Botanical Research,2011,31(2):227-230.
introduction:GUO Hai-Xia(1986—),female,doctor,lecturer,research on alpine plant ecology.
date:2016-12-21
EffectsofShadingandNitrogenAdditionontheGrowthandC-NBalanceofFritillariaunibracteata
GUO Hai-Xia1XU Bo3SHI Fu-Sun2WU Yan2*
(1.Chengdu Normal University,Chengdu 611130;2.Ecological Restoration and Biodiversity Conservation Key Laboratory of Sichuan Province,Chengdu Institute of Biology,Chinese Academy of Sciences,Chengdu 610041;3.Aba Teachers University,Wenchuan 623002)
In order to check howFritillariaunibracteataHsiao et K.C.Hsia adjusts to resources variation, a controlled field experiment was set, andF.unibracteata’s responses of growth and C-N balance to shading and nitrogen addition were examined.F.unibracteatawas highly responsive to shading and nitrogen addition. Nitrogen content in leaf and root showed significant increase after shading and nitrogen addition, which induced the decrease of C∶N in leaf and root. The ratio of leaf biomass to root biomass(R1) was significantly increased(P<0.05) under shading treatment, while there was no obvious change of R1under nitrogen-addition treatment. Finally, there were significant correlations(P<0.01) between nitrogen content, C∶N and total plant biomass, leaf-root biomass ratio. Therefore, C-N balance inF.unibracteatawould be a better index of resource strategy.
Fritillariaunibracteata;shading;nitrogen addition;biomass allocation;C-N balance
郭海霞(1986—),女,博士,讲师,主要从事高山植物生态研究。
* 通信作者:E-mail:260763666@qq.com
2016-12-21
* Corresponding author:E-mail:260763666@qq.com
S567.23+1
A
10.7525/j.issn.1673-5102.2017.05.014
猜你喜欢
中国饲料(2022年17期)2022-09-08
中国土壤与肥料(2021年5期)2021-12-02
中国核电(2021年4期)2021-08-25
艺术品鉴(2021年12期)2021-05-25
科学技术创新(2020年6期)2020-04-18
热力透平(2019年4期)2019-12-18
中成药(2017年5期)2017-06-13
上海农业学报(2017年4期)2017-04-10
农家顾问(2014年9期)2014-10-20
中国野生植物资源(2014年4期)2014-03-29
