
马尾松三代选择群体生长性状变异及选择研究
2017-11-09 13:37余启新滕国新金国庆丰忠平周志春
植物研究 2017年2期
张 振 余启新 滕国新 金国庆* 丰忠平 周志春
(1.中国林业科学研究院亚热带林业研究所,浙江省林木育种技术研究重点实验室,国家林业局马尾松工程技术研究中心,富阳 311400; 2.浙江省淳安县新安江开发总公司,淳安 311700; 3.浙江省兰溪苗圃马尾松国家良种基地,兰溪 321100)
马尾松三代选择群体生长性状变异及选择研究
张 振1余启新2滕国新3金国庆1*丰忠平2周志春1
(1.中国林业科学研究院亚热带林业研究所,浙江省林木育种技术研究重点实验室,国家林业局马尾松工程技术研究中心,富阳 311400; 2.浙江省淳安县新安江开发总公司,淳安 311700; 3.浙江省兰溪苗圃马尾松国家良种基地,兰溪 321100)
以浙江省淳安县姥山林场的6×6半双列遗传交配设计的三代测定幼林为对象,分析幼林期(2、3和5 a)的生长性状的遗传参数和育种值,及2 a生时生物量的累积与分配。结果表明,马尾松幼林期不同杂交组合间的树高、地径、冠幅和活枝数均存在显著的遗传差异,且生物量在2 a生亦存在显著的遗传差异。杂交组合2 a生的地上生物量占植株总生物量的87.17%。幼林期生长性状以树高的增长量最大,5 a生较2 a生增长了4.23倍。采用综合育种值法,依据预测的树高(H)育种值为主,结合地径(D0)和2 a生茎干生物量(Bs)指标的配合选择方式,以入选率20%优选出杂交组合分别为22×44、33×22、40×44。以5 a生时树高、地径的单株育种值为依据,2%的入选率优选出16个单株,其树高、地径、冠幅及活枝数平均增益分别为1.07 m、1.58 cm、0.32 m和4.67个。
马尾松;生物量;生长量;育种值
育种材料的评价与选择是林木遗传改良的重要途径和环节。育种值是性状表型值中加性遗传效应部分,它剔除了环境的影响,反映遗传效应的大小,是选择育种中的重要参数[1~3]。杂交育种中通过配合力的研究,可以估算亲本无性系的育种值,但配合力会随着交配群体变化的情况下跟着变化,而育种材料的基因加性效应没有变化,因此个体的育种值可以从亲本遗传方差中固定的传递给子代。基因型选择代替表型选择,通过表型值信息进行个体育种值评定可提高选择效率和精度[4]。
马尾松(Pinusmassoniana)是我国南方主要的造林树种,分布17个省(市、区),适应性强,生长快。前期项目组已针对不同遗传交配设计的测定林材料,系统开展生长、材性和干物质积累等性状的配合力分析,揭示所受的遗传控制式样[5~7]。全同胞家系中子代独特的基因型组合取决于交配亲本所含有的特定基因[3]。一般配合力(GCA)为单个亲本的特质,特殊配合力(SCA)特定于双亲交配子代的平均表现。谭小梅等[6]对本研究材料的1年生苗高生长量分析表明,SCA效应均明显地大于GCA效应;且Balocchi等[8]对火炬松(Pinustaeda)配合力的研究结论表明,未经过GCA选择和测定的材料,早期树高主要受显性基因效应控制;因此,在马尾松生长性状的早期选择过程中可注重对杂交组合子代的评价。White等[3]和孙晓梅等[4]均指出个体优良基因型选择在高世代育种中对优化种子园种子品质,提高子代遗传增益的重要性。已有研究证明,杂交子代与亲本的表现并不完全一致,且不同亲本间组配能力也不同,综合评价子代测定林的表型性状及对育种值的有效预测,可为高世代遗传改良提供材料[9~11]。
马尾松生长性状在幼林期已具有较高的遗传变异,且早、晚相关表明,材积、材性等性状的早期具有较大的选择潜力[12~15]。前期关于马尾松的遗传选择的相关研究较多,而通过预测育种值综合评价多个年度马尾松试验测定林,分析马尾松幼龄期生物量分配及其与生长性状相关的研究较少涉及。本研究选用马尾松2代育种亲本,采用6×6半双列交配设计创制的双亲控制授粉子代测定林,开展2、3和5 a生的马尾松生产力遗传参数和育种值分析,解析2 a生时干物质积累量,揭示马尾松幼龄期生物量分配模式及其与生长性状的相关,旨为马尾松高世代遗传改良提供依据。
1 材料与方法
1.1 试验材料
供试材料为6×6半双列交配设计创制的双亲控制授粉子代测定林。亲本来自浙江省淳安县姥山林场的马尾松2代育种群体。2008年底收获种子,2009年播种育苗,2010年1月营建在浙江省淳安县姥山林场苗圃,2012年1月改建于姥山林场试验林基地,交配设计见表1。其中,亲本33号(1145×1126)、20号(1103×3412)、40号(5163×5907)、22号(6627×3412)、56号(3412×5907)为一代亲本杂交后代(一代亲本选择子代林中优良单株),亲本44号为种源1123子代中优良单株,以当地马尾松优良天然林分子代作对照1(CK1),以当地一代种子园混系采种子代作为对照2(CK2)。试验林采用随机完全区组设计,10株小区,株行距2 m×2 m,重复5次。测定林地处于中亚热带北缘(119°03′E,29°37′N),海拔150~250 m,年平均气温17℃,≥10℃的年积温为5 410℃,年降水量1 430 mm,全年日照时数1 951 h,土壤为山地黄土,立地条件中等。
表1马尾松6×6半双列交配设计
Table1Schematicdiagramofthe6×6halfdiallelcrossdesign

母本Femaleparents父本Maleparents33204022564433×××××20××××40×××22××56×44
1.2 数据采集测定
分别在2011、2012和2014年的10月份进行测定林的每木调查。调查树高、地径、冠幅、活枝数及当年抽梢性状值[16]。在马尾松2 a生进行生物量的测定,即每个组合根据树高与地径的小区平均值分别在每个小区选择2单株,称量针叶、枝、茎干、主根及须根的总鲜重(g),取样称重并带回实验室80℃烘干至恒重,称量各部位的干物质重(g)。
1.3 遗传统计分析
以小区内单株测定值为单位,采用SPSS19.0软件检验各性状测定值在不同杂交组合间的遗传差异显著性。方差分析采用以下线性模型Ⅰ:
Yijkl=μ+fij+bk+(fb)ijk+eijkl
(1)
式中:μ为总平均值;fij为第i亲本与第j亲本杂交组合效应;bk为第k个区组效应;(fb)ijk为杂交组合与区组的交互效应;eijkl为随机误差[17]。
方差分析时年抽梢次数和活枝数均经平方根数据转换。采用MTDFREML软件[18]单株模型估算遗传方差组分,估算杂交组合及组合内单株育种值。综合育种值(IC)为3个测定年度平均育种值,即为
(2)
式中,Ij为第j年的育种值。
2 结果与分析
2.1 幼龄马尾松生物量变化规律
2 a生的生物量方差分析结果表明,幼龄马尾松的针叶、枝条、茎段、主根及地上部分的生物量在不同杂交组合间的遗传差异皆达到1%显著水平(40×22号杂交组合保存率仅为14%,没有进行方差分析)。马尾松2 a生的地上生物量占植株总生物量的87.17%,其中,针叶、枝条、茎干、主根和须根分别占生物量的43.44%、18.44%、25.24%、6.52%和6.28%。参试的所有家系中,33×22号杂交组合的针叶、枝条、主根及地上生物量在15个杂交组合中均最高,分别高于最低杂交组合的91.44%、92.29%、79.84%及80.89%,分别高于CK1的121.67%、145.20%、122.63%及144.30%。22×44号组合的茎干最高,高于最低杂交组合的89.78%,高于CK1的213.87%。

表2 生物量分配及方差分析结果
注:**代表1%显著水平;*代表5%显著水平 下同。
Note:*and**mean significance at the 0.05 and 0.01 probability levels respectively. The same as below.
2.2 生物量与生长性状相关性
植株各部位生物量与生长性状相关性分析表明(表3),树高、地径、冠幅及活枝数与地上生物量极显著正相关,与地下生物量的主根部分极显著正相关;须根仅与地径极显著正相关。总体呈现幼龄马尾松随着树高、地径及冠体积的增长,地上各部分及主根生物量随之增长,而须根的生长量受地径的影响明显。另外,抽梢性状中自由生长量及抽梢次数分别和茎干呈极显著和显著地正相关,说明幼林期抽梢次数与自由生长能显著地影响茎干的生长。
2.3 马尾松生长性状的年度变化
表4列出了马尾松杂交组合的生长性状在3个树龄的均值表现及变异趋势。分别比较树高、地径、冠幅和活枝数在4年间的生长量,树高增长3.91 m,地径增长4.45 cm,冠幅增长2.24 m,活枝数增加17.25个。树高增长幅度最大,5 a生较2 a生增长了4.23倍。
方差分析结果表明,生长性状在2、3和5 a生均表现出极显著的杂交组合效应。由组合间的变化幅度可知,2 a时树高、地径、冠幅及活枝数生长最快杂交组合分别是对照CK1的104.44%、42.88%、47.20%和123.25%;3a时树高、地径、冠幅及活枝数生长最快杂交组合分别是对照CK1的69.29%、38.01%、40.66%和78.34%;5 a时树高、地径、冠幅及活枝数生长最快杂交组合分别是对照CK1的46.90%、74.61%、27.97%和83.35%。说明创制的杂交组合间持续存在丰富的变异,具有较大的选择潜力,有依据结合性状表型值,分析遗传效应大小,选择优良组合和单株。
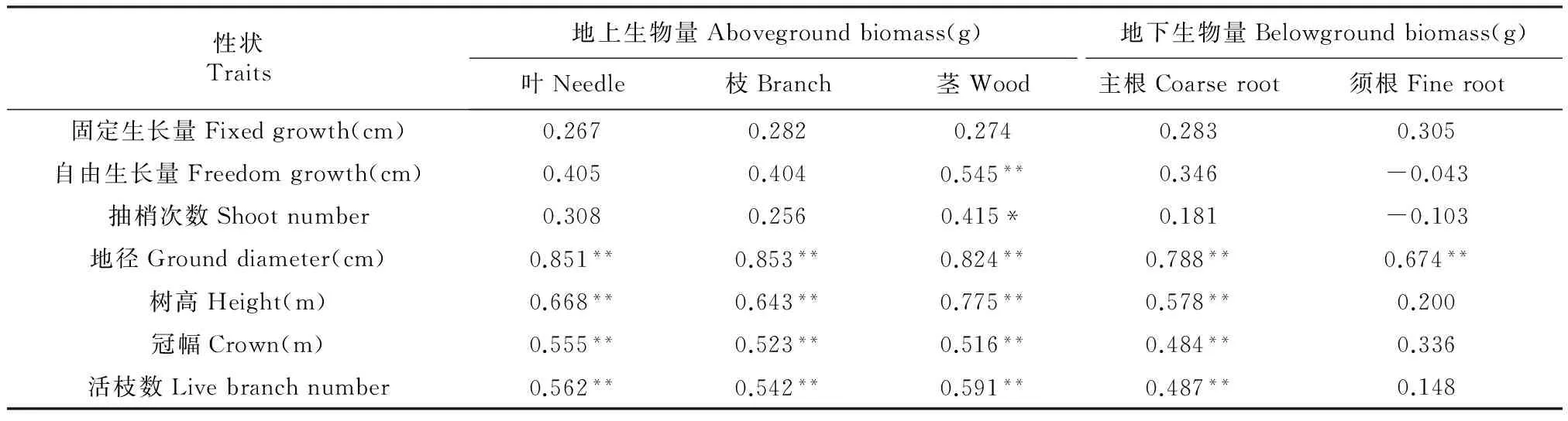
表3 各部分生物量与生长性状相关性分析
注:*表示在5%水平(双侧)上显著相关;**表示在1%水平(双侧)上显著相关。
Note:*and**indicate significant difference(bilateral) at 0.05 or 0.01 level,respectively.

表4 生长性状方差分析
注:生长性状对应的组合、区组、区组×组合的自由度分别为:16、4和64;3 a时保存率为88.59%;5 a时保存率为86.35%。
Note:DF of cross combination, block and cross combination×block is 16,4 and 64,respectively;Born three years and five years of survival rate is 88.59% and 86.35%, respectively.
2.4 育种值预测和选择
2.4.1 杂交组合的选择
项目组前期研究马尾松树高与胸径呈极显著的正相关关系[19]。本研究材料为马尾松幼龄林,通过分析生长性状的变化规律,依据预测的树高(H)育种值进行选择为主,采用配合选择并结合地径(D0)和2 a生茎干生物量(Bs)指标选择优良杂交组合。如表5所示,首先估算各年度杂交组合的育种值,然后估算3个年度的综合育种值(IC)。结合IC值分别选择树高(H)、地径(D0)和茎干生物量(Bs)的综合育种值排名前5位的杂交组合。结果显示,树高、地径与茎干生物量指标的IC值排名前五名的杂交组合相同,仅排序有所不同,分别为22×44、33×44、33×22、40×44和20×44组合。而组合20×44和33×44的保存率较低,分别为72%和84%。最终,以入选率为20%优选的杂交组合分别为22×44、33×22、40×44。22×44组合的树高、地径和生物量的IC值均为最高,茎干生物量和5年生树高、地径分别高于CK1的213.87%、46.90%和68.01%,高于CK2的110.36%、32.71%和59.14%。
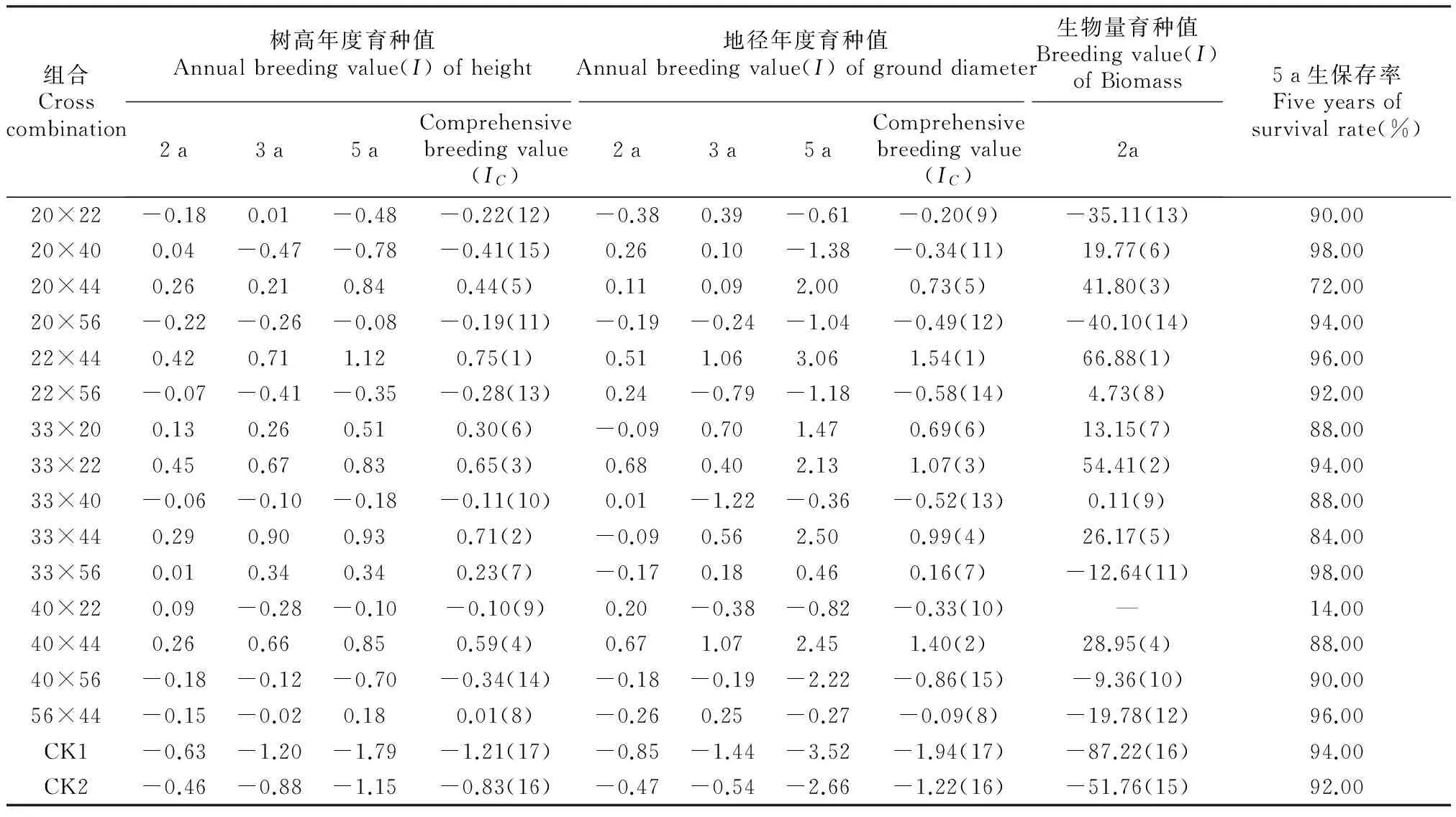
表5 杂交组合预测的育种值
注:“()”内为综合育种值排名顺序。
Note:“()” is comprehensive breeding value order.

表6 入选单株5年生时生长表现和预测育种值
2.4.2 单株选择
以5 a生时树高、地径的单株育种值为依据,根据预测的育种值以2%的入选率选出16个优良单株(表6)。为保证尽可能宽的遗传基础,适当选择所在杂交组合育种值较高、个体育种值排在前列的单株。在单株选择过程中,在生长表现差异不大的情况下,树高育种值高的单株优先入选。优选的16个单株树高较群体均值增益1.07 m,地径增益1.58 cm,冠幅增加0.32 m,活枝数平均增加4.67个。入选的树高平均生长量为5.74 m,地径为8.72 cm,均显著高于对照。
3 讨论
目前,在马尾松的遗传改良过程中,家系和单株选择仍然是常用的育种手段。通过杂交制备杂种将生产力强的有利基因牢固,是传统遗传改良中最有成效的一种育种技术[20~25]。2 a生马尾松的针叶、枝条、茎干、主根及地上部分的生物量遗传测定结果表明,在不同杂交组合间存在极显著的遗传差异;2、3和5 a的生长性状在杂交组合间均达到极显著差异,与周志春[7]和谭小梅等[6]在马尾松全同胞交配设计的杂交组合间的遗传差异研究结果一致。2 a生茎干生物量与5 a生的树高及胸径的变化幅度分别在85.84~162.91 g、4.21~6.52 m和5.87~8.90 cm,说明杂交组合间存在丰富的变异,通过杂交制种创造变异并利用变异可开展优良杂交组合及单株的选择。谭小梅等[6]利用本研究材料苗期的苗木生长及苗高生长参数的GCA/SCA进行分析,提出马尾松苗期生长性状所受遗传控制式样和赵颖等[26]与周志春[7]等林木幼林期研究结果类似,同为SCA的选择较GCA重要,且苗高的显性基因效应方差分量占总基因效应方差分量的81.8%。在其研究的15个半双列杂交组合中,基于苗高SCA优选出的33×44、22×44、33×22和40×44组合同为本研究优选的杂交组合,说明这些组合自苗期一直保持着较强杂种优势。亲本44、33及22号无性系均出现在优选的组合中,说明其具有较高的组配能力,从而进一步印证了谭小梅等[6]的研究指出的亲本44、33和22号无性系的GCA最高。另外,亲本间具有亲缘关系的4个半同胞杂交组合的育种值的树高及地径的表型值较低等水平,同谭小梅等[6]趋于中等表现水平的研究结果不一致。推测可能的原因为,随着树龄增加马尾松树高的变异系数有增加的趋势[12],1、2、3及5 a生树高的遗传变异系数分别为15.65%[6]、18.84%、23.14%及28.61%,且与试验林保存率下降有一定关系[27~28],从而影响其遗传方差组分;从亲本来源分析,因杂交组合的亲本间具有半同胞家系的亲缘关系,且半同胞家系子代的育种值一般来自于母本,故致使该群体的遗传均值和遗传方差的改变,较亲缘关系远的亲本创制的杂交组合性状平均表现低[29]。
马尾松各部位生物量随着树龄增加而增加,树干生物量增长速度最快[15]。本研究材料为幼林,茎干生物量所占比例为25.24%,而马尾松成熟龄树干生物量比例达到70%左右[15]。树高及胸径与树干生物量呈显著地正相关[3],本研究树高、地径、冠幅及活枝数与地上生物量极显著正相关,与地下生物量的主根部分极显著正相关;须根仅与地径极显著正相关。总体呈现幼龄马尾松随着树高、地径及冠体积的增长,地上各部分及主根生物量随之增长,而须根的生长量受地径的影响明显。抽梢性状中自由生长量及抽梢次数分别和茎干呈极显著和显著地正相关,说明幼林期抽梢次数与自由生长能显著地影响茎干的生长,与Bridgwater等[30]和周志春等[16]表明的抽梢性状可反映树木生长期内的生长规律,影响树木高生长的结论相似。
育种值预测是遗传评价的核心,对于下一育种世代来说,在计算单株育种值的基础上选择优良单株可以提高期望遗传增益[31]。本研究材料为幼林,以树高的测定更为准确。同时,抽梢性状可反映树木生长期内的生长规律,是影响树木高生长的重要因子[16,32]。因此,以预测的树高育种值为主要选择依据,地径及生物量为配合选择,以入选率20%进行杂交组合选择,选择出22×44、33×22、40×44组合;为保证尽可能宽的遗传基础,结合抽梢性状的表现适当选择所在杂交组合育种值较高、个体育种值排在前列的单株,以入选率2%进行单株选择,选择出16个单株。刘天颐[29]分析14年生马尾松全同胞家系育种值表明,采用配合选择方式,入选家系的胸径遗传增益达到8.56%,这为本研究以树高选择为主,茎干生物量和地径作为辅的配合选择提供主要理论依据。
在林木遗传改良实践中,因遗传交配设计不同,亲本群体及其包含的家系数量和试验林保存率等各不相同,育种值的精确预测变得复杂多样化[33]。本研究采用多性状非求导约束最大似然法(MTDFREML)程序对杂交组合和参试个体进行育种值预测,通过校正环境因素影响,能准确估测遗传参数,预测个体的多性状育种值[6]。通过建立混合模型,确定变量,以区组作为固定效应;随机效应中,父本×母本(SCA)效应、杂交组合×区组效应可用于计算个体随机效应。软件运行的关键是将每一单株准确给予一个ID,根据父母本生成相应的PED文件和根据变量生成DAT文件。我们进行遗传参数估算常采用家系模型Ⅰ,而进行单株育种值估算时要采用以下模型(Ⅱ):
Yijkl=μ+ID+bk+(fb)ijk+eijkl
(3)
式中:μ为总平均值;ID为单株号;bk为第k个区组效应;(fb)ijk为第i亲本与第j亲本杂交组合与区组的交互效应;eijkl为随机误差。
这样,通过对应单株的数量和ID,估算出组合及每个单株的育种值。本研究测定结果为马尾松三代遗传测定林幼林期,其生长性状表现仍不稳定,具有一定的阶段性特点。虽然育种目标是提高轮伐期的材积生长量,但一般要在幼林期就对目标性状进行选择[34]。因此,后期应对试验林的不同发育阶段的遗传控制式样研究,探明影响材积等重要经济性状的基因位点所起的作用大小;注重年年遗传相关,提高早期选择效率。
1.Cotterill P P,Dean C A.Successful tree breeding with index selection[M].Melbourne:National Library of Australia Cataloguing,1990.
2.马浩,张冬梅,李荣幸,等.泡桐属植物育种值预测方法的研究[J].林业科学,2003,39(1):75-80.
Ma H,Zhang D M,Li R X,et al.Study on predicting method of breeding values inPaulownia[J].Scientia Silvae Sinicae,2003,39(1):75-80.
3.White T L,Thomas A W,Neale D B.Forest genetics[M].Wallingford,London:CAB International,2007.
4.孙晓梅,杨秀艳.林木育种值预测方法的应用与分析[J].北京林业大学学报,2011,33(2):65-71.
Sun X M,Yang X Y.Applications and analysis of methods for breeding value prediction in forest trees[J].Journal of Beijing Forestry University,2011,33(2):65-71.
5.金国庆,秦国峰,刘伟宏,等.马尾松测交系杂交子代生长性状遗传分析[J].林业科学,2008,44(1):70-76.
Jin G Q,Qin G F,Liu W H,et al.Genetic analysis of growth traits on tester strain progeny ofPinusmassoniana[J].Scientia Silvae Sinicae,2008,44(1):70-76.
6.谭小梅,金国庆,周志春.马尾松3代种质苗高生长参数的配合力分析[J].林业科学研究,2011,24(5):663-667.
Tan X M,Jin G Q,Zhou Z C.Combining ability analysis on seedling shoot elongation and growth parameters for the third generation germplasm ofPinusmassoniana[J].Forest Research,2011,24(5):663-667.
7.周志春,金国庆,秦国峰,等.马尾松纸浆材重要经济性状配合力及杂种优势分析[J].林业科学,2004,40(4):52-57.
Zhou Z C,Jin G Q,Qin G F,et al.Analysis on combining ability and heterosis of main economic traits ofPinusmassonianafor pulp production[J].Scientia Silvae Sinicae,2004,40(4):52-57.
8.Balocchi C E,Bridgwater F E,Zobel B J,et al.Age trends in genetic parameters for tree height in a nonselected population of loblolly pine[J].Forest Science,1993,39(2):231-251.
9.Huang M,Chen L Y,Chen Z Q.Diallel analysis of combining ability and heterosis for yieldand yield components in rice by using positive loci[J].Euphytica,2015,205(1):37-50.
10.Worku M,Banziger M,Friesen D,et al.Relative importance of general combining ability and specific combining ability among tropical maize(ZeamaysL.) inbreds under contrasting nitrogen environments[J].Maydica,2008,53:279-288.
11.张振,张含国,张磊.红松自由授粉子代家系生产力年度变异与家系选择[J].植物研究,2016,36(2):305-309.
Zhang Z,Zhang H G,Zhang L.Age variations in productivity and family selection of open-pollinated families of Korean pine(Pinuskoraiensis)[J].Bulletin of Botanical Research,2016,36(2):305-309.
12.金国庆,秦国峰,刘伟宏.不同林龄马尾松的种源选择效果[J].林业科学,2011,47(2):39-45.
Jin G Q,Qin G F,Liu W H.Provenance selection effect at different stand age ofPinusmassoniana[J].Scientia Silvae Sinicae,2011,47(2):39-45.
13.刘青华,周志春,范辉华,等.马尾松产脂力与生长性状的家系变异及优良家系早期选择[J].林业科学研究,2013,26(6):686-691.
Liu Q H,Zhou Z C,Fan H H,et al.Variation for oleoresin yielding capability and growth among families and early selection for superior families inPinusmassoniana[J].Forest Research,2013,26(6):686-691.
14.周全连,杨章旗,覃开展.马尾松自由授粉子代测定及优良家系选择[J].广西科学,2001,8(1):63-65.
Zhou Q L,Yang Z Q,Qin K Z.Open pollination progeny test and selection of good families inPinusmassoniana[J].Guangxi Sciences,2001,8(1):63-65.
15.丁贵杰,王鹏程.马尾松人工林生物量及生产力变化规律研究Ⅱ.不同林龄生物量及生产力[J].林业科学研究,2001,15(1):54-60.
Ding G J,Wang P C.Study on change laws of biomass and productivity of masson pine forest plantation Ⅱ.Biomass and productivity of stand at different ages[J].Forest Research,2001,15(1):54-60.
16.周志春,戴德升,吴吉富,等.不同磷营养环境下马尾松优良种源的抽梢行为[J].林业科学研究,2001,14(1):54-59.
Zhou Z C,Dai D S,Wu J F,et al.Shoot growth behavior of superior Masson pine provenances under different phosphorus environment[J].Forest Research,2001,14(1):54-59.
17.续九如.林木数量遗传学[M].北京:高等教育出版社,2006.
Xu J R.Quantitative genetics in forestry[M].Beijing:Higher Education Press,2006.
18.Boldman K G,Kriese L A,Van Vleck L D,et al.A Manual for Use of MTDFREML[M].U.S.Department of Agriculture:Agricultural Research Service,1995.
19.周志春,金国庆,周世水.马尾松自由授粉家系生长和材质的遗传分析及联合选择[J].林业科学研究,1994,7(3):263-268.
Zhou Z C,Jin G Q,Zhou S S.Genetic analysis and combined selection for growth and wood quality of open-pollinated families of Masson pine[J].Forest Research,1994,7(3):263-268.
20.Park Y S,Gerhold H D.Population hybridization in Scotch pine(PinussylvestrisL.) I.Genetic variance components and heterosis[J].Silvae Genetica,1986,35(4):159-165.
21.Baltunis B S,Wu H X,Dungey H S,et al.Comparisons of genetic parameters and clonal value predictions from clonal trials and seedling base population trials of radiata pine[J].Tree Genetics & Genomes,2009,5(1):269-278.
22.Sluder E R.Results at age 15 years from a half-diallel cross among 10 loblolly pines selected for resistance to fusiform rust(Cronartiumquercuumf.sp.Fusiforme)[J].Silvae Genetica,1993,42(4-5):223-230.
23.齐明.杉木育种中GCA与SCA的相对重要性[J].林业科学研究,1996,9(5):498-503.
Qi M.Relative lmportance of GCA and SCA in Genstic breeding of Chinese fir[J].Forest Research,1996,9(5):498-503.
24.王赵民,陈益泰.杉木主要生长性状配合力分析及杂种优势的利用[J].林业科学研究,1988,1(6):614-624.
Wang Z M,Chen Y T.An analysis on the combining ability of main growth character in Chinese fir and the application of its Heterosis[J].Forest Research,1988,1(6):614-624.
25.李力,施季森,陈孝丑,等.杉木两水平双列杂交亲本配合力分析[J].南京林业大学学报,2000,24(5):9-13.
Li L,Shi J S,Chen X C,et al.Combining ability analyses of parents in two-level diallel cross experiment of Chinese Fir[J].Journal of Nanjing Forestry University,2000,24(5):9-13.
26.赵颖,周志春,金国庆.马尾松苗木生长和根系性状的GCA/SCA及磷素环境影响[J].林业科学,2009,46(6):27-33.
Zhao Y,Zhou Z C,Jin G Q.GCA SCA of seedling growth and root parameters inPinusmassonianaand the phosphorus environment influence[J].Scientia Silvae Sinicae,2009,46(6):27-33.
27.Wu H X,Powell M B,Yang J L,et al.Efficiency of early selection for rotation-aged wood quality traits in radiata pine[J].Annals of Forest Science,2007,64(1):1-9.
28.Kumar S,Lee J.Age-age correlations and early selection for end-of-rotation wood density in radiata pine[J].Forest Genetics,2002,9(4):323-330.
29.刘天颐,杨会肖,刘纯鑫,等.火炬松基因资源的育种值预测与选择[J].林业科学,2014,50(8):60-67.
Liu T Y,Yang H X,Liu C X,et al.Prediction of breeding values and selection to the gene resources of loblolly pine[J].Scientia Silvae Sinicae,2014,50(8):60-67.
30.Bridgwater F E,Willums C G,Canpbell R G.Patterns of leader elongation in loblolly pine families[J].Forest Sciences,1985,31(4):933-944.
31.Xiang B,Li B L.Best linear unbiased prediction of clonal breeding values and genetic values from full-sib mating designs[J].Canadian Journal of Forest Research,2003,33(10):2036-2043.
32.黄永权,钟伟华,廖文莉,等.火炬松优良家系抽梢生长的调查研究[J].福建林业科技,2006,33(4):142-151.
Huang Y Q,Zhong W H,Liao W L,et al.Study on sprouting growth of superior families ofPinustaeda[J].Journal of Fujian Forestry Science and Technology,2006,33(4):142-151.
33.White T L,Hodge G R.Predicting breeding values with applications in forest tree improvement[M].Netherlands:Springer,1989.
34.Politov D V,Belokon M M,Maluchenko O P,et al.Genetic evidence of natural hybridization between siberian stone pine,Pinussibiricadu tour,and dwarfSiberianpine,P.pumila(Pall.) Regel[J].Forest Genetics,1999,6(1):41-48.
This work was supported by “Twelfth five-year” national science and technology support project(2012BA01B02);National natural science foundation of China(31600533);Major science and technology special project of bamboo new agricultural varieties breeding in Zhejiang province(2016C02056-4)
introduction:ZHANG Zhen(1986—),male,assistant researcher,engaged in the genetic breeding and improvement of pinus massoniana.
date:2016-11-07
VariationandSelectionofGrowthCharacterontheYoungThirdGenerationofPinusmassoniana
ZHANG Zhen1YU Qi-Xin2TENG Guo-Xin3JIN Guo-Qing1*FENG Zhong-Ping2ZHOU Zhi-Chun1
(1.Research Institute of Subtropical Forestry,Chinese Academy of Forestry,Key Laboratory of Tree Breeding of Zhejiang Province,Engineering Research Center of Masson Pine of State Forestry Administration,Hangzhou 311400;2.Development Corporation of Xin’an river of Chun’an Country,Zhejiang Province,Chun’an 311700;3.National fine species base of pinus massoniana of Lanxi nursery,Lanxi 321100)
We analyzed the genetic parameters, breeding value, accumulation and distribution of 2-year raw biomass with a half diallel cross among 6×6 on three generation young growth period of forest on Laoshan farm in Zhejiang Province. There were significant genetic differences among the tree height, ground diameter, crown breadth and number of living branch in the young period ofPinusmassoniana. There were also significant genetic differences in the period of 2-year biomass between different hybrid combinations. The ground biomass accounted for 87.17% of the total plant biomass in 2-year hybrid combination. The height growth was the largest in the young period of tree growth trait. The height growth was increased by 4.23 times from 5 to 2 years. According to method of comprehensive breeding value, the tree height(H) breeding value combined with conjointly diameter(D0), and 2 a raw stem biomass index(Bs) to choose higher survival rate of hybrid combinations. The higher survival rate combinations were 22×44, 33×22 and 40×44 based on 20% year old open pollination progenies. Based on the value of tree height and ground diameter of plant breeding, 16 number of plant were chosen accounting for 2%. The average gain were 1.07 m(tree height), 1.58 cm(ground diameter), 0.32 m(crown breadth) and 4.67(living branch number), respectively.
Pinusmassoniana;biomass;growth;breeding value
“十二五”国家科技支撑课题“南方针叶树高世代育种技术研究与示范”(2012BA01B02);国家自然科学基金资助项目(31600533);浙江省农业(林木)新品种选育重大科技专项重点课题(2016C02056-4)
张振(1986—),男,助理研究员,主要从事马尾松遗传育种与改良。
* 通信作者:E-mail:jgqin@163.com
2016-11-07
* Corresponding author:E-mail:jgqin@163.com
S718.46
A
10.7525/j.issn.1673-5102.2017.02.015
猜你喜欢
农业灾害研究(2022年6期)2022-12-02
南京林业大学学报(自然科学版)(2022年1期)2022-11-29
西南农业学报(2022年5期)2022-06-06
林业科学(2021年5期)2021-07-13
中国糖料(2021年3期)2021-07-13
农业机械学报(2020年3期)2020-04-01
绿色科技(2019年5期)2019-11-29
种子(2019年5期)2019-07-02
热带农业科技(2019年1期)2019-01-14
中国果业信息(2019年11期)2019-01-05
