
MC1R与Slc7a11基因多态性与哈萨克羊毛色相关的研究
2017-10-23 06:59:20何军敏阿依达尔霍江台木合依扎热很别克黄锡霞
中国畜牧杂志 2017年10期
何军敏,孙 军,余 雄,阿依达尔.霍江台,木合依扎.热很别克,黄锡霞*
(1.新疆农业大学动物科学学院,新疆乌鲁木齐 830052;2.特克斯县畜牧兽医局,新疆特克斯 835500)
MC1R与Slc7a11基因多态性与哈萨克羊毛色相关的研究
何军敏1,孙 军2,余 雄1,阿依达尔.霍江台2,木合依扎.热很别克1,黄锡霞1*
(1.新疆农业大学动物科学学院,新疆乌鲁木齐 830052;2.特克斯县畜牧兽医局,新疆特克斯 835500)
本研究采集新疆伊犁哈萨克自治州特克斯县4种不同被毛颜色2~3周岁的220只哈萨克母羊血样用于提取DNA,其中黑色被毛羊52只,棕色被毛羊52只,青灰色被毛羊59只,白色被毛羊57只;并采集绵羊左侧肩胛皮肤样本,每种毛色各10个,用于组织形态学观察。本研究运用PCR-SSCP技术及测序技术对MC1R基因及Slc7a11基因多态性及其对哈萨克羊毛色的影响进行研究。运用皮肤样本制作切片用于不同毛色组织形态学观察。结果表明:MC1R基因突变位点在白色和棕色被毛群体中,只存在BB基因型;而在黑色和青灰色被毛群体中,存在AA、AB和BB 3种基因型;且AB基因型均为优势基因型,A等位基因为优势等位基因。由组织形态学观察可知,黑色和青灰色被毛组织毛囊分布极为相似,毛囊分布稀疏且皮脂腺发达;棕色被毛最具特色,毛囊丛分布明显,皮脂腺分布不发达;白色被毛毛囊分布最为密集,且皮脂腺分布密集并且最为发达。经χ2检验,MC1R基因的基因型频率在4种毛色羊中的分布差异极显著(P<0.01)。Slc7a11基因位点在伊犁哈萨克羊群体中发现其不存在多态性。
哈萨克绵羊;MC1R基因;Slc7a11基因;PCR-SSCP;遗传多态性
哈萨克绵羊是我国3大绵羊品种之一,同时也是新疆的优秀地方绵羊品种。目前国内外对于MC1R基因对哈萨克羊毛色影响的研究报道较少,尤其本研究中哈萨克羊群体存在4种毛色,目前仅在特克斯县发现第4种毛色的哈萨克羊。孙浩浩[1]研究报道了TYR基因、ASIP基因多态性在绵羊中的相关分析。毛色受TYR、ASIP、MC1R、TYRP1等多种基因控制和影响,其通过参与生物合成活性或者调节有关黑色素生成细胞的分布过程等来调控毛色[2-3]。MC1R与黑色素细胞刺激激素α结合激活腺苷酸环化酶(cAMP),最终增加了黑色素的合成[4]。当Slc7a11基因突变后,导致其编码的蛋白缺乏,使得氨基酸在转运载体时失去功能,导致细胞不能摄取到足够的胱氨酸,因此无法还原成半胱氨酸,从而无法满足细胞中合成伪黑色素的需要,最终导致毛色主要表现为真黑色素的黑颜色,从而缺乏由伪黑色素产生的黄色[5-7]。王乐等[8]总结了目前MC1R基因在各种动物中的研究,但关于哈萨克羊存在青灰色被毛的研究至今未见报道。本实验旨在研究特克斯县哈萨克羊形成4种毛色的原因,从而进一步了解哈萨克羊毛色形成的规律,完善哈萨克羊毛色基因的研究。
1 材料与方法
1.1 实验材料 本研究采集新疆伊犁哈萨克自治州特克斯县4种不同被毛颜色2~3周岁的220只哈萨克母羊血样及40个肩胛皮肤组织,所有绵羊毛色均为纯色,不含有任何斑点或其他杂色,头毛与蹄毛均与被毛一致,均为纯色。其中黑色被毛样本52个,棕色被毛样本52个,青灰色被毛样本59个,白色被毛样本57个,血样-20℃保存用于提取基因组DNA;采集绵羊左侧肩胛皮肤组织每种毛色各10个,用福尔马林保存用于组织形态学观察。
1.2 方法 绵羊血样基因组DNA运用酚-氯仿法进行抽提。皮肤组织由兰州军区总医院组织切片制作室进行切片制作。运用PCR-SSCP技术分析MC1R基因在不同颜色被毛哈萨克羊的多态性,以提取的绵羊基因组DNA为模板进行PCR,反应体系参照Taq DNA聚合酶说明书。运用显微镜进行组织形态学观察并进行图像采集。
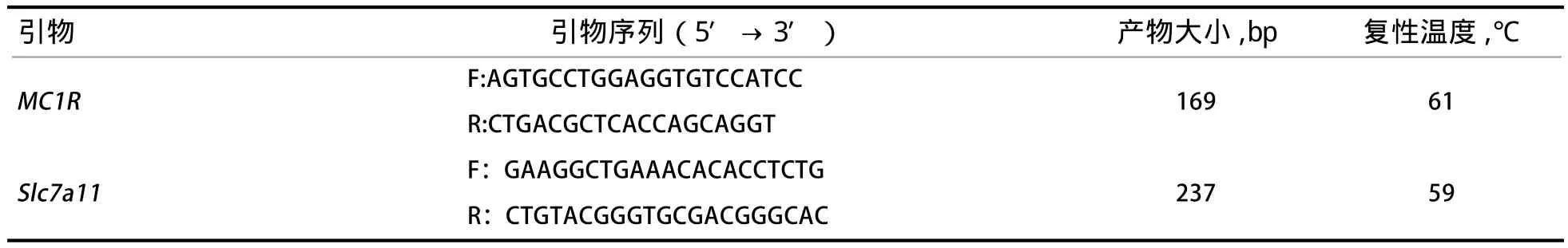
表1 PCR扩增的引物序列
1.2.1 引物设计 本研究MC1R基因(GenBank登录号为FN600553)引用李洪涛[9]的引物序列,Slc7a11基因(GenBank登录号为JQ085938)采用Primer Premier 5.0软件设计引物,由上海生工合成,引物序列如表1所示。
1.2.2 PCR扩增程序的建立 PCR扩增条件:经94℃预变性3 min;94℃变性30 s,MC1R基因61℃、Slc7a11基因59℃退火45 s,72℃延伸45 s,共35个循环;72℃延伸5 min。于低温保存待后续实验。
1.2.3 PCR反应体系的建立 PCR扩增反应体系20 μL,其中 10 μL 2× PCR Master Mix,1 μL DNA 模板,上、下游引物(10 μmol/L)各0.5 μL,8 μL ddH2O 。
1.2.4 PCR产物测序 经过普通PCR扩增及2%琼脂糖凝胶电泳检测合格后,将PCR产物送到上海生工进行单向测序。
1.2.5 PCR-SSCP过程 扩增产物经2%琼脂糖凝胶电泳检测,取3 μL的PCR产物,加入7 μL 上样缓冲液(9.8 mL甲酰胺,10 mg 溴酚蓝,10 mg二甲苯青, 0.5 mol/LEDTA 0.2 mL)混合,98℃变性10 min,立即冰浴15 min,并用12%聚丙烯酰胺凝胶(高板)进行电泳检测。300 V 预电泳5 min 使样品快速入胶,然后室温恒压180 V 电泳16~18 h。银染液显色后拍照,统计各基因型的个体数。
1.2.6 组织切片制作 皮肤组织送往兰州军区总医院组织切片制作室进行切片制作,对皮肤组织进行横切后制成切片,运用显微镜进行观察并采集图片。
1.3 统计分析 运用SPSS 18.0软件对数据进行χ2独立性检验,并进行多态信息含量(PIC)、遗传纯合度(Ho)、杂合度(He)和有效等位基因数(Ne)的计算。
2 结果与分析
2.1 组织形态学观察 由图1可知,在白色被毛组织中,毛囊分布密集但并未形成明显的毛囊丛,毛囊分布均匀。皮脂腺分布密集较为发达。由图2可知,在黑色被毛皮肤组织中,毛囊分布较白色稀疏,同样未有明显的毛囊丛存在,皮脂腺发达并且分布均匀。由图3可知,在青灰色皮肤组织中,与黑色皮肤组织极为相似,毛囊分布稀疏,皮脂腺发达,分布均匀。由图4可知,在棕色皮肤组织中,形成了明显的毛囊丛,单一的毛囊丛中毛囊分布密集,毛囊丛之间分布较为离散,皮脂腺不发达。

图1 白色被毛皮肤组织(100×)
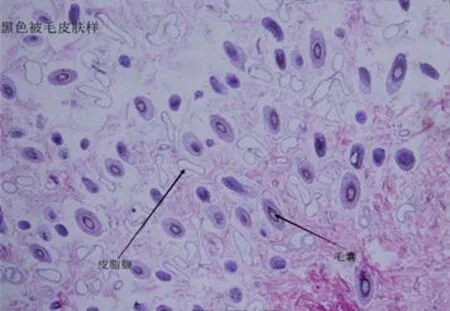
图2 黑色被毛皮肤组织(100×)
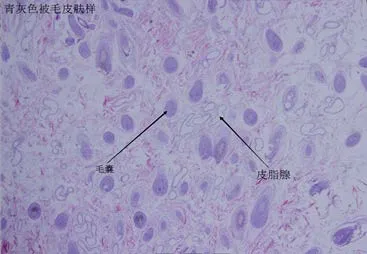
图3 青灰色被毛皮肤组织(100×)

图4 棕色被毛皮肤组织(100×)
2.2 DNA完整性检测结果 将提取的基因组DNA用1%的琼脂糖凝胶进行电泳检测。结果表明,实验提取的基因组DNA模板质量良好、纯度高、不需要纯化,可直接用作模板DNA。
2.3 MC1R基因的PCR扩增结果 PCR产物经2.0%的琼脂糖凝胶进行电泳检测,可知MC1R与Slc7a11基因引物PCR产物长度为169 bp和237 bp,与生物公司提供片段符合。结果表明,PCR扩增特异性良好,片段长度一致,引物的片段长度符合预期大小,可直接用于PCR-SSCP的分析。
2.4 MC1R和Slc7a11基因的PCR-SSCP多态性检测结果 如图5、6所示,MC1R基因存在3种不同的带型,将其命名为AA、AB、BB型;Slc7a11基因均为AA基因型,不存在多态性。

图5 MC1R的SSCP

图6 Slc7a11基因的SSCP产物
2.5 MC1R序列分析 由测序结果分析可知,在哈萨克羊群体中发现此序列在91 bp处发生突变(图7)。由AA基因型突变为BB基因型,AB为杂合子。通过对氨基酸序列的分析发现,其A91T突变点为非同义突变,由甲硫氨酸突变为赖氨酸。
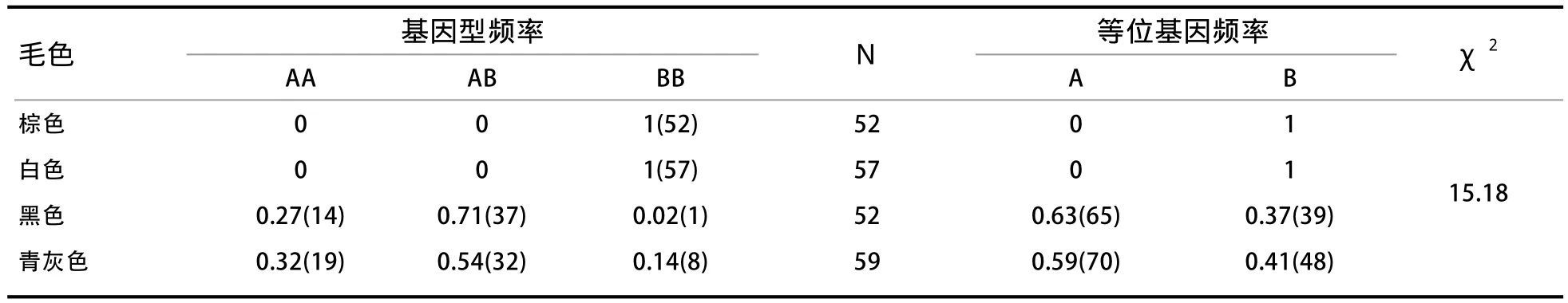
2.6 MC1R的遗传多态性分析 由表2可知,MC1R基因在不同毛色中均有差异。在白色和棕色被毛群体中,只存在BB基因型;在黑色被毛群体中,存在AA、AB和BB 3种基因型,其基因型频率分别为0.27、0.71和0.02,且BB基因型频率所占比例极小。由此可知,AB基因型为优势基因型;其A和B等位基因频率分别为0.63和0.37,可知A等位基因为优势等位基因;而在青灰色被毛群体中,存在AA、AB和BB 3种基因型,其基因型频率分别为0.32、0.54和0.14,由此可知AB基因型为优势基因型;其A和B等位基因频率分别为0.59和0.41,可知A等位基因为优势等位基因。经χ2检验,MC1R基因的基因型频率在4种毛色羊中的分布差异极显著(P<0.01)。

图7 MC1R突变点不同基因型测序峰图

表3 哈萨克羊MC1R基因遗传参数
2.7 MC1R基因遗传参数分析 由表3可知,MC1R基因在该哈萨克羊群体中黑色与青灰色均处于中度多态(0.25 大量研究表明,多数脊椎动物MC1R基因的突变能够引起毛色的变化,如小鼠、牛、马、鸡、狐狸、猪、绵羊、山羊和狗,均发现毛色由浅变深;对人的研究亦是如此[10]。对小鼠进行遗传学研究表明 ,MCIR是一种独特的具有双功能控制的受体。对猪的 MC1R 基因已经定位 ,并且已建立了PCR-RFLP技术来检测不同猪种的等位基因的多肽。研究发现绵羊中的extension 等位基因为ED, 从而导致显性的黑色毛色。Vage等[11]分析了黑色被毛绵羊的MC1R基因,发现其存在2个突变即Asp121Asn 与Met73Lys。而对引进鼠的相对应的MC1R 基因进行药理学分析发现,Met73Lys突变点是组成型活性的MC1R,其可能是导致绵羊的黑色毛色的主要原因[8]。姜俊兵等[12]实验证明,羊驼MC1R基因中存在着单链构象多态性,存在AA和AB 2种基因型。 表2 MC1R基因频率和基因型频率的分布以及Hardy-Weinberg平衡的卡方检验 本研究结果发现,MC1R基因突变的AB基因型与黑色被毛极显著相关,说明该位点对黑色被毛有显性作用,这与Vage等[11]报道的显性黑等位基因(ED)控制黑色被毛的结果相一致。本研究发现,哈萨克羊被毛的颜色与MC1R基因有密切联系,它的突变决定着黑色和青灰色被毛的性状,且在黑色与青灰色被毛中AB基因型所占频率大。MC1R基因的基因型频率在4种毛色羊中的分布差异极显著。黑色与青灰色被毛中存在AA、AB和BB 3种基因型,与李洪涛等[9]的结果有一定差异;而在白色和棕色群体中仅存在BB基因型,与李洪涛等[9]的结果有一定的相似性。李洪涛等[9]将MC1R和ASIP进行基因型联合分析时没有发现对毛色有互补效应,说明控制哈萨克绵羊黑色被毛的主要基因是MC1R。对于绵羊的白色和棕色可能存在着其他位点的调控。有报道称,Spotting位点决定着绵羊的白色被毛[13],Slc7a11 位点与绵羊的棕色被毛相关。而本研究Slc7a11基因位点在本实验中运用PCR-SSCP技术并未检测到其多态性,有待进一步研究。 经皮肤组织形态学观察发现在白色被毛组织中,毛囊分布密集且均匀;皮脂腺分布密集较为发达[14]。在黑色被毛皮肤组织中,毛囊分布较白色分布稀疏,皮脂腺发达并且分布均匀。在青灰色皮肤组织中,与黑色皮肤组织极为相似,毛囊分布稀疏,皮脂腺发达,分布均匀。在棕色皮肤组织中,形成了明显的毛囊丛,单一的毛囊丛中毛囊分布密集,毛囊丛之间分布较为离散,皮脂腺不发达[15]。黑色与青灰色皮肤组织观察极为相近,而白色与棕色各具特色,白色毛囊密集或许因白色毛发吸收光的能力较弱因而保暖更为密集,而其他颜色相对白色对光的吸收能力强,棕色形成了明显的毛囊丛,此原因有待进一步研究。棕色皮脂腺较其他颜色不发达,也许与毛囊丛的形成在组织学中有一定的关系,有待专业人士进一步分析。 本研究分析了MC1R和 Slc7a11基因多态性与哈萨克绵羊被毛颜色之间的关系。结果表明,哈萨克羊被毛颜色与MC1R基因有密切联系,且在黑色与青灰色被毛中AB基因型所占频率大。MC1R基因的基因型频率在4种毛色羊中的分布差异极显著。Slc7a11基因位点在此群体中运用PCR-SSCP技术未检测到多态性。由组织形态学可知,黑色与青灰色被毛组织毛囊分布极为相似;棕色被毛最具特色,毛囊丛分布明显,皮脂腺分布不发达;白色被毛毛囊分布最为密集,且皮脂腺分布密集并且最为发达。 [1] 孟浩浩. TYR、ASIP基因多态性与绵羊毛色相关的研究[D]. 石河子: 石河子大学, 2014. [2] Fontanesi L, Dall'olio S, Beretti F, et al. Coat colours in the Massese sheep breed are associated withmutations in the agouti signalling protein (ASIP) and melanocortin 1 receptor (MC1R) genes[J]. Animal, 2010, 5(1): 8-17. [3] Royo L J, Ailvarez I, Arranz J J, et al. Differences in the expression of the ASIP gene are involved in the recessive black coat colour pattern in sheep: evidence from the rare Xalda sheep breed[J]. Anim Genet, 2008, 39(3):290-293. [4] Slominshi A, Tobin D J, Shibahara S, et al. Melanin pigmentation in mammalian skin and its hormonal regulation [J]. Physiol Rev, 2004, 84:1155-1228. [5] 张俊珍, 董常生, 范瑞文, 等. 哺乳动物毛色形成研究进展[J]. 动物医学进展, 2006, (S1): 65-68. [6] 孙笑尉, 张雪莲, 李振, 等. 染色质重塑因子BPTF、BRG1在不同毛色绵羊皮肤的表达与定位[J].畜牧兽医学报, 2017, 48(3):446-453. [7] 王准. Gas在绵羊耳部不同毛色皮肤中的表达与定位[D].太原: 山西农业大学, 2016. [8] 王乐, 张斌, 郑文新, 等. 动物毛色与黑色素皮质素受体1(MC1R)基因[J]. 草食家畜, 2009, (2):10-12. [9] 李洪涛, 曾献存, 张文祥, 等. 哈萨克绵羊MC1R和ASIP基因多态性及表达量与被毛颜色表型相关性的研究[J].畜牧兽医学报, 2013, (3):366-375. [10] Valvered P, Healy E, Jackson I, et al. Variants of the melanocyte-stimulating hormone receptor gene are associated with red hair and fair skin in humans[J]. Nat Genet, 1995, 11:328-330. [11] Vage D I, Lu D D, Dlungland D, et al. A non-epistatic interaction of Agouti and extension in the fox,Vulpes vulpes[J]. Nat Genet, 1997, 15:311-315. [12] 姜俊兵, 任杰, 于菊秀, 等. 中国羊驼种群MC1R基因的PCR-SSCP分析[J]. 家畜生态学报, 2010, 31(3): 15-18. [13] Kerns J A, Cargill E J, Clark L, et al. Linkage and segregation analysis of black and brindle coat color in domestic dogs[J]. Genetics, 2007, 176:1679-1689. [14] 姜晓龙, 陈建伟, 姚晓磊, 等. 绵羊卵巢组织石蜡切片制作条件优化研究[J]. 上海畜牧兽医通讯, 2013, (6):2-3. [15] 牟艳军, 廖和荣, 赵宗胜, 等. 生长激素受体(GHR)mRNA在绵羊皮肤组织切片定位[J]. 安徽农业科学,2006, (7):1295-1296. S826.2 A 10.19556/j.0258-7033.2017-10-039 2017-03-31; 2017-04-05 农业部体系项目(CARS-40);疆农业大学产学研联合培养研究生示范基地项目(xjaucxy-yjs-20152011) 何军敏(1991-),女,新疆乌苏人,硕士研究生,主要从事动物遗传育种的研究,E-mail: 1415230131@qq.com *通讯作者:黄锡霞(1963-),教授,博士生导师,主要从事动物遗传育种的研究,E- mail:au- huangxixia@163.com3 讨 论
4 结 论
猜你喜欢
丝绸之路(2023年2期)2023-07-10 21:34:47
军事文摘(2023年2期)2023-02-17 09:20:24
养猪(2021年4期)2021-08-26 10:57:46
感染、炎症、修复(2021年1期)2021-07-28 06:18:08
心声歌刊(2021年2期)2021-07-16 07:05:56
卫拉特研究(2018年0期)2018-07-22 05:47:28
畜牧兽医科学(2018年14期)2018-02-14 01:44:16
中国民族医药杂志(2016年9期)2016-05-09 07:51:52
保健与生活(2016年1期)2016-04-12 18:29:44
畜牧兽医学报(2015年3期)2015-07-05 08:22:51
