
西藏冬青稞胚乳淀粉粒发育特征研究
2017-10-16 07:07:23侯维海王建林
麦类作物学报 2017年9期
侯维海,王建林,旦 巴,胡 单
(西藏农牧学院植物科学学院,西藏林芝 860000)
西藏冬青稞胚乳淀粉粒发育特征研究
侯维海,王建林,旦 巴,胡 单
(西藏农牧学院植物科学学院,西藏林芝 860000)
为了解青稞胚乳淀粉及淀粉粒形成与生长特征,以西藏育成青稞品种冬青8号为材料,对籽粒发育过程中胚乳灌浆速度、淀粉积累量及淀粉粒的产生、二维面积、粒径变化进行了分析。结果表明,胚乳发育过程中,干物质和淀粉积累量呈S型曲线,两者呈正相关。花后5 d,青稞胚乳中可零星见到粒径和二维面积较小的A型小淀粉粒;花后10~15 d,A型小淀粉粒数量显著增大,但粒径和二维面积增加较慢;花后20 d,B型小淀粉粒可能已经形成,与A型小淀粉粒组成一个复合群体;花后25 d,形成了粒径>10 μm的A型淀粉粒、粒径5~10 μm的B型淀粉粒和粒径<5 μm的C型淀粉粒;花后30~55 d,不同粒径的淀粉粒数量和二维面积逐渐增加,其中,C型淀粉粒数量不断增加直至籽粒成熟。青稞成熟籽粒胚乳中,淀粉粒的粒径为1.45~44.76 μm,二维面积为3.56~1 750.8 μm2;A型淀粉粒形状为近圆形,表面光滑,个别表面附着有C型淀粉粒,B型淀粉粒形状有近圆形、椭圆形多种类型,C型淀粉粒形状比B型淀粉粒的变异更大。
冬青稞;淀粉粒;淀粉粒形成
Abstract: To study the formation and growth of hulless barley endosperm starch and starch granules,hulless barley Dongqing 8(cultivated in Tibet) was chosen as the target material to analyze endosperm filling rate,starch accumulation as well as the formation,2D area and size of starch granules in the process of granule development. The results showed that during the development of endosperm,dry matter auumulation and starch accumulation perform an S curve,and are positively related to each other. On the 5 days post anthesis(DPA),A-type starch granules with small sizes and 2D areas can be sparsely seen in hulless barley endosperm. During 10 to 15 DPA,the number of small A-type starch granules increased significantly,but their size and 2D area showed a slow increase. On 20 DPA,B-type small starch granules may come into being and they will be combined with A-type small starch granules to form a composite polymer. On 25 DPA,the size of A-type starch granules was above 10 μm,and the size of B-type starch granules ranged from 5 to 10 μm,and the size of C-type starch granules was less than 5 μm. During 30 to 55 DPA,the number and 2D areas of starch granules with different sizes increased a lot,and the number of C-type starch granules grew until the granules mature. Diameter of starch granules in mature granule endosperm of hulless barley ranged from 1.45 μm to 44.76 μm,and the 2D area ranged from 3.56 μm2to 1 750.8 μm2. A-type starch granules is roughly spherical with smooth surface. C-type starch granules adhered to surface of some A-type starch granules. B-type starch granules are shaped in sub-spherical and oval. Shape of type-C starch granules had greater variation than that of B-type starch granules.
Keywords: Winter hulless barley; Starch granule; Starch granule formation
大麦(HordeumvulgareL.)是世界最古老的作物之一,也是第四大谷类粮食作物,其收获面积和产量仅次于小麦、水稻和玉米[1]。在青藏高原地区,裸大麦又称青稞(Hordeumvulgaressp.vulgare),是藏族人民主要粮食作物之一和主要的饮品原料[2]。淀粉是大麦籽粒最主要的储藏碳水化合物,占籽粒干重的50%~60%。淀粉由直链淀粉和支链淀粉组成,以大小不同的淀粉粒形式贮存在于胚乳中[3]。基于淀粉粒粒径和形状的差异,一般将小麦族植物胚乳淀粉粒分为A、B两种类型,也有A、B、C三种类型之说和大、中、小的分级学说[4]。一般A型淀粉粒粒径>10 μm,形状似圆盘或扁豆状,B型淀粉粒粒径<10 μm,形状为近圆形[5],将粒径<5 μm、形状不规则的淀粉粒称为C型淀粉粒[6],C型淀粉粒通常被认为是B型淀粉粒的亚群,一般将其归为B型淀粉粒。植物不同物种及同一物种不同基因型间淀粉粒形状、体积、 数量、粒径及淀粉含量有一定差异,遗传是决定这些差异的关键因素[6-8]。青稞不同基因型间的A型淀粉粒和B型淀粉粒的数量间有显著差异[9]。外界环境因素也是影响籽粒发育、淀粉积累及淀粉粒粒径分布的重要因素。温度、水分亏缺、营养、光照强度均可显著改变淀粉粒的粒度分布和直链淀粉含量[9,10-11]。此外,穗位不同,籽粒淀粉粒径也有一定差异,如小麦中间小穗籽粒比顶部和基部小穗籽粒有更高的B型淀粉粒占比[12]。
不同类型的淀粉粒有不同的结构组成、生化特性,进而影响淀粉热特性、糊化特性、膨胀特性、烘焙特性等理化性质,最终影响大麦籽粒的加工品质[13-15]。研究表明,A型淀粉粒有更高的糊化焓、黏连参数,B型淀粉粒有更高的脂质复合直链淀粉含量和膨胀能力、更宽的糊化范围及较低的糊化焓[16]。大麦胚乳中的小淀粉粒较多则有利于造纸和化妆品工业,大淀粉粒多则有利于酿造工业[17];小麦A型淀粉粒可用于无碳复写纸的制造,而B型淀粉粒是较好的塑料薄膜填料[18]。因此,为培育具有不同类型淀粉粒的青稞品种,有必要了解青稞胚乳发育过程中不同淀粉粒的动态发育规律。
前人对小麦不同类型淀粉粒的形成规律已有深入研究[19]。有关大麦胚乳淀粉粒发育、淀粉积累量的动态变化研究较少,且尚无青稞胚乳淀粉粒变化特征的相关报道。因此,本研究以西藏育成青稞品种冬青8号为材料,研究籽粒胚乳发育过程中淀粉粒表型和淀粉积累量的变化及其相互关系,以期为西藏青稞育种奠定理论基础。
1 材料与方法
1.1 试验材料与设计
供试材料为冬青8号,于2015年3月中旬种植于西藏农牧学院农场(E94°25′,N29°59′)。随机区组排列,3个重复,小区面积2 m×3 m,露地条播,行距20 cm,基本苗为330 万·hm-2。管理措施同当地常规大田生产。从开花到成熟期,设定12个发育阶段,分别为花后5、10、15、20、25、30、35、40、45、50、55、60 d。在开花期选取生长发育一致、健壮植株的主茎进行挂牌标记。在设定的各发育阶段,剪取3个主茎穗,密封放入-80 ℃冰箱保存,用于胚乳淀粉粒生长发育动态的显微观察;剪取10个主茎穗用于灌浆速度测定;剪取10个主茎穗,密封放入冰箱保存,用于胚乳发育过程中淀粉含量测定。
1.2 方 法
1.2.1 总淀粉含量的测定
按照GB/T5514-2008方法检测青稞种子总淀粉含量。
1.2.2 总淀粉粒的提取
参照Peng等[20]方法并做了部分改进。取2粒青稞籽粒放入1.5 mL的离心管中,加入1 mL 0.5 mol·L-1的NaCl溶液中浸泡12 h,置研钵中研磨至匀浆,6 000 r·min-1离心5 min,弃上清,沉淀依次用2 mol·L-1的NaCl、2% SDS 和0.2%的NaOH溶液洗涤,以去除蛋白质,至沉淀变为白色;用丙酮清洗3 次,风干,于-20 ℃贮存备用。每个样品重复2 次。
1.2.3 干物质积累量的测定
将青稞胚乳发育12个阶段剪取的10个主茎穗放入105 ℃烘箱,杀青30 min,80 ℃烘至恒重,测定百粒重。
1.2.4 花后气象数据采集
通过农田小气候自动检测站记录青稞花后田间气候变化。
1.2.5 扫描电镜观察、拍照
用棉棒粘取适量1.2.2中提取的淀粉粒干粉,均匀涂抹于载玻片上,滴60 μL浓度为20%的甘油,封片,并在酒精灯下烤至微干,用Leica DM5000 生物显微镜放大400倍,每个样品随机选取5个视野进行观察并拍照。
1.3 数据处理
利用EXCEL对淀粉含量、灌浆速度、气温变化进行数据统计分析,利用Imagepro plus对淀粉粒电子显微图片进行处理,每个视野随机选取至少200个淀粉粒,根据比例尺设定标尺,测量其面积、最大粒径、最小粒径、平均粒径及圆度值的实际值,进行统计分析。
2 结果与分析
2.1 花后籽粒表型的变化
青稞花后籽粒发育的12 个阶段,其籽粒颜色、粒长、粒宽的变化如图1所示(花后5、10、15 d,籽粒小,无拍照),花后5 d,已有极小的籽粒形成;花后5~25 d,籽粒以纵向伸长生长为主,粒宽增加缓慢;在花后25~55 d,粒长变化不明显,粒宽、粒厚逐渐增加;花后55 d后,籽粒生长几乎停滞。籽粒颜色从浅褐色到灰色转变发生在花后45 d,粒长最大值发生在花后35 d,粒宽的增加基本贯穿于整个籽粒发育的全过程。

图1 青稞花后20 d至成熟籽粒表型的变化
2.2 籽粒总淀粉含量的变化
淀粉是青稞种子最主要的贮藏物质,在胚乳发育12 个阶段,其淀粉含量呈S型变化趋势(图2)。冬青8号在花后5 d,籽粒淀粉含量约占干重的3.21%,随着籽粒干物质积累量的增加,淀粉含量逐渐增加,其中,在花后20~50 d,淀粉含量增幅最大,基本呈一次函数递增,50 d后,淀粉含量增幅减小。
2.3 干物质积累量与温度的相关分析
青稞干物质积累量表现为“慢-快-慢”的S型增长曲线(图3-A)。根据百粒重增长速度可以将青稞籽粒灌浆过程分为三段,第一段为灌浆渐增期,在花后5~20 d,百粒重为0.364 g;第二段为灌浆快增期,在花后20~55 d,百粒重达到3.991 g,是青稞灌浆的关键期;第三段为灌浆缓增期,在花后55 d以后,干物质积累量几乎无增加,百粒重达到最大值。
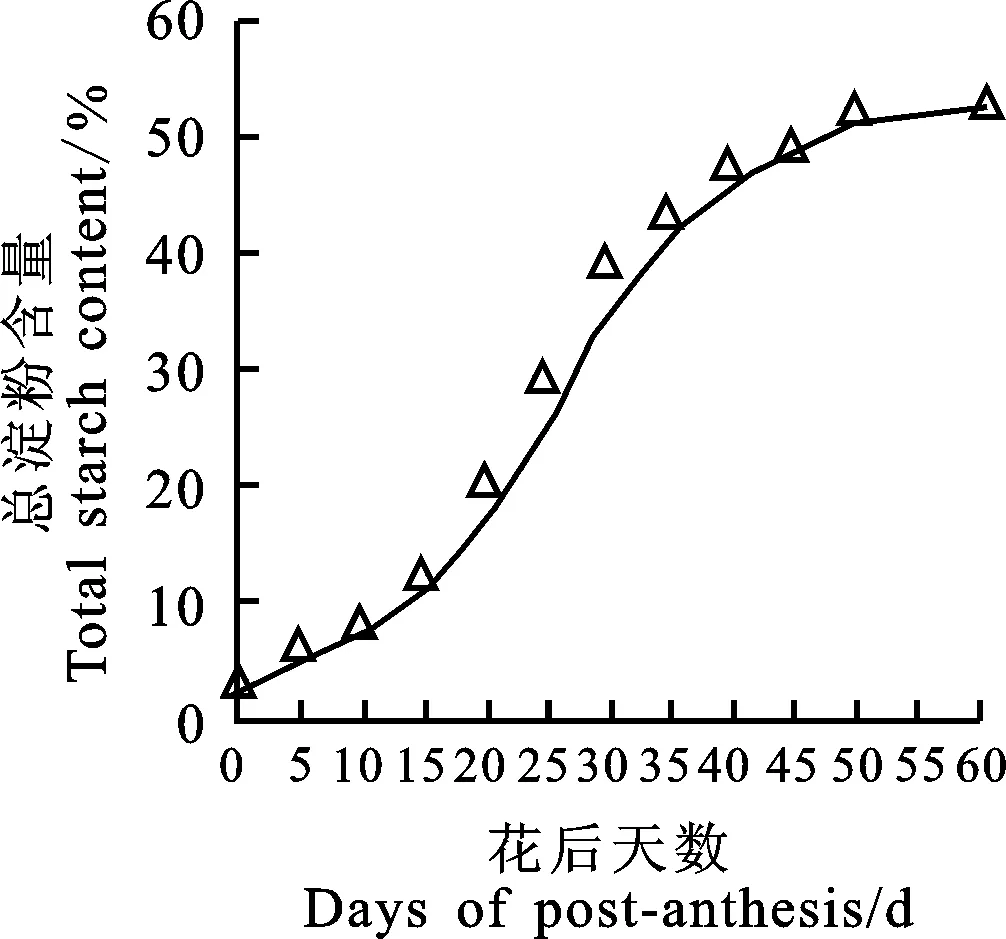
图2 青稞胚乳中总淀粉含量的变化
由图3-B可见,在青稞灌浆渐增期,即花后5~20 d,日最高温为16.4~18.4 ℃,平均高温为17.5 ℃,日最低温在3.2~5.2 ℃,平均低温为4.5 ℃;进入青稞灌浆快增期,即花后20~55 d,日最高温有较大幅度波动,但日最低温表现出持续升高趋势,平均高温为19.1 ℃,平均低温为6.8 ℃,日最高温和最低温比灌浆渐增期提高2~3 ℃。在青稞灌浆缓增期,日最高和日最低温达到年极限温度,并保持相对稳定状态。
综合分析青稞花后干物质积累量与气温变化发现,日最低温度和日平均温度显著影响青稞籽粒干物质积累速度(P<0.05)。在青稞花后1~30 d,高温相对较低,低温处于5 ℃左右,干物质积累较缓;花后30 d以后,低温持续上升,高温缓慢上升,干物质积累快速。推测低温是影响青稞干物质积累和灌浆速度的限制因素。
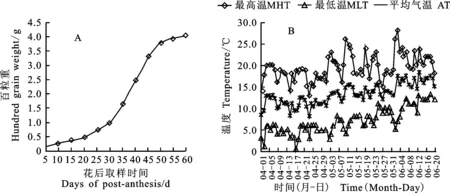
MHT:Highest temperature;MLT:Lowest temperature; AT:Average temperatur
2.4 青稞成熟籽粒淀粉粒的形态
观察青稞成熟籽粒胚乳淀粉粒,根据粒径可将淀粉粒分为A型淀粉粒(>10 μm)、B型淀粉粒(5~10 μm)和C型淀粉粒(<5 μm)。在二维形状上,A型淀粉粒大多为近圆形,部分呈椭圆形,表面光滑,少部分表面零星分散有较小的C型淀粉粒(图4-A);B型淀粉粒形状有近圆形、椭圆形、扁圆形等(图4-B),而C型淀粉粒形态变化较大,有近圆形、椭圆形、多边形及不规则形(图4-C)。在淀粉粒数量上,C型淀粉粒占主导地位,B型淀粉粒数量略多于A型淀粉粒;就淀粉粒表面积占比而言,A型、B型淀粉粒远大于C型淀粉粒。

图4 青稞成熟籽粒淀粉粒的分布
2.5 青稞籽粒发育过程中淀粉粒的形态变化
在青稞胚乳发育过程中,淀粉粒发育如图5和表1所示。花后5 d,胚乳中有零星的淀粉粒产生,呈单独的圆粒型且体积较小,直径1.11~3.34 μm,平均粒径为2.03 μm,平均表面积3.89 μm2;花后10 d时,胚乳中的淀粉粒数量逐渐增多,体积略有增加,粒径为1.56~4.09 μm,平均粒径为2.95 μm,平均表面积8 μm2;15 d~20 d时淀粉粒数量和体积继续增加,体积增加较明显且淀粉粒轮廓变得清晰,淀粉粒数量显著增多;25 d 时,进入淀粉粒发育的高峰期,其形态发生了巨大变化,表现为淀粉粒间相互黏连和聚合,粒径、表面积显著增大,部分淀粉粒由圆形变成不规则的多边形,扁圆形等,形成了可显著辨识的三类淀粉粒,其中,A型淀粉粒平均粒径14.24 μm,平均表面积173.07 μm2,B型淀粉粒平均粒径7.09 μm,平均表面积42.37 μm2,C型淀粉粒平均粒径3.01 μm,平均表面积9.96 μm2;30~55 d,A型淀粉粒粒径、表面积逐渐增大,B和C型淀粉粒不断形成;55 d后,A型淀粉粒粒径生长逐渐减慢,数量几乎无增加。综上所述,在青稞胚乳发育过程中,A、B、C型淀粉粒粒径、面积、数量都表现出一定的时空表达特性,其中以A型淀粉粒粒径、面积和数量变化最为明显。

图5 青稞籽粒中淀粉粒发育动态
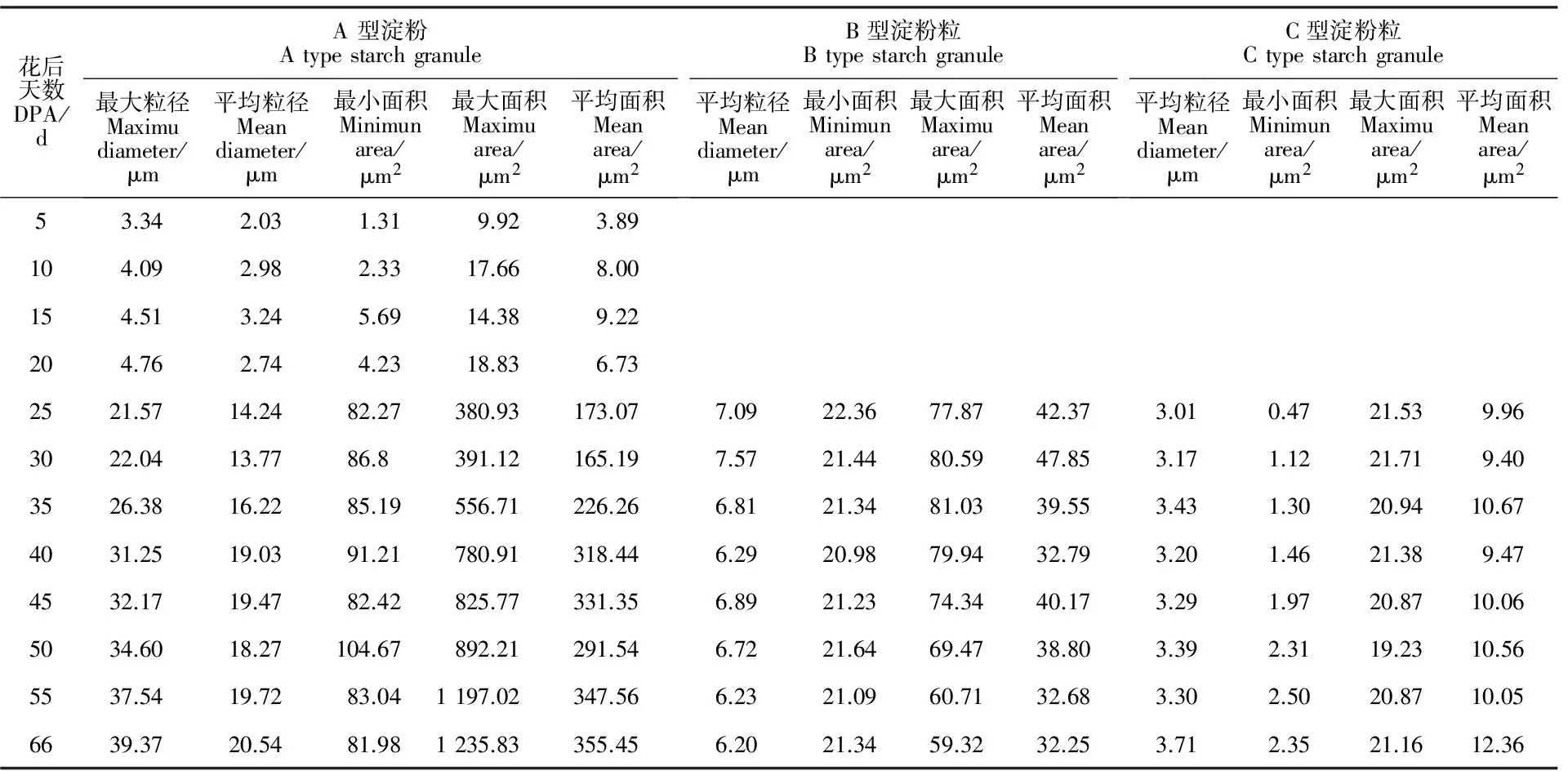
表1 青稞胚乳不同发育时期淀粉粒粒径和表面积Table 1 Starch granule size and area at different developmental stages of hulless barley endosperm
DPA:Days post-anthesis.
3 讨 论
目前,有关小麦、大麦等小麦族植物淀粉粒的分型存在一些争议。一些研究者认为,小麦籽粒胚乳淀粉粒可分为三类,粒径>10 μm的淀粉粒为A型淀粉粒、5~10 μm的淀粉粒为B型淀粉粒,<5 μm的淀粉粒为C型淀粉粒[21]。另一些学者认,粒径<5 μm的淀粉粒为C型淀粉粒,5~16 μm之间的淀粉粒为B型淀粉粒,>16 μm的淀粉粒为A型淀粉粒[22];Raeker等[6]认为,3种淀粉粒之间的界限应该是粒径为2.8和9.9 μm;Francesca等[23]将淀粉粒分为两类,粒径<8 μm为B型淀粉粒,粒径8~30 μm的为A型淀粉粒。Sarita等[24]对139份不同基因型大麦淀粉粒径研究认为,>15 μm淀粉粒为A型淀粉粒,5~15 μm淀粉粒为B型淀粉粒,<5 μm的淀粉粒为C型淀粉粒,其中A型淀粉粒平均粒径为19.3(16.51~22.23) μm。本研究通过对青稞淀粉粒光学显微拍照和图片数据处理,发现在青稞花后25 d,其淀粉粒粒径、表面积、数量均可清晰的分为大、中、小三种类型,以粒径10 μm作为划分大、中淀粉粒的界限,以粒径5 μm作为划分中、小淀粉粒的界限,小型淀粉粒数量几乎占绝大多数,中型淀粉粒数量显著多于大型淀粉粒数量,而大、中型淀粉粒表面积显著大于小淀粉。因此,建议将青稞淀粉粒按照粒径大小分为>10 μm的A型淀粉粒、5~10 μm的B型淀粉粒和<5 μm的C型淀粉粒。青稞成熟胚乳中,淀粉粒径最大粒径为39.37 μm,较Sarita等[24]报道的大麦淀粉粒粒径大。这可能是因为品种不同或测量技术以及软件计算方法不同所导致的。
大量研究表明,淀粉粒粒径与淀粉含量关系紧密,A型淀粉粒比B型淀粉粒的直链淀粉含量高4~10%[23,25],Raeker 等[6]认为,淀粉含量与淀粉粒径5~9.9 μm的体积占比呈负相关,淀粉直支比与粒径2.8~9.9 μm的体积占比呈负相关,与粒径在22.8~42.8 μm体积占比呈正相关。Li等[24]认为,直链淀粉含量与粒径>22 μm的淀粉粒体积比显著正相关,而与<10 μm的淀粉粒体积占比显著负相关,表明>22 μm的淀粉粒有更高的直链淀粉含量。这些研究表明,淀粉粒数量、 粒径和面积的变化与籽粒总淀粉含量有显著关系。本研究测定了青稞胚乳发育的12个阶段的淀粉含量及淀粉粒径、表面积,结果表明,总淀粉含量、淀粉表型(数量、粒径、面积)间存在正向同步性,与前人结果一致。
小麦族植物胚乳发育过程中,A型、B型淀粉粒动态积累具有独立的时空特性。前人利用激光粒度分析仪技术对普通小麦淀粉粒发育过程的研究结果表明,在普通小麦、大麦、小黑麦等小麦族植物中,A型淀粉粒一般在花后4 d开始形成,花后7 d就出现不同粒径的淀粉粒,最大粒径达到20 μm,在花后17~19 d,A型淀粉粒不再产生,主要以体积增大为主;B型淀粉粒在花后14~16 d产生,随着胚乳发育推移,数量和大小不断增长;而C型淀粉粒首次出现在花后21 d,在胚乳成熟前,大量C型淀粉粒形成[12]。本研究利用光学显微镜对青稞胚乳发育的12 个阶段淀粉粒进行观察,结果表明,青稞花后5 d已有平均粒径达到1.78 μm、最大直径达到3.12 μm的小淀粉粒出现,推测A型淀粉粒在花后3 d左右产生;在花后10、15、20 d,淀粉粒平均粒径分别为2.45、3.24、3.74 μm,淀粉粒粒径变化并不大,数量显著增多,表明已形成的A型淀粉粒处于持续发育阶段,新的小A型淀粉粒不断形成,在花后20 d,可能有B型小淀粉粒产生;花后25 d,已形成粒径>10 μm的A型淀粉粒、5~10 μm的B型淀粉粒和<5 μm的小淀粉粒,此阶段,A型淀粉粒体积显著增大,数量变化不大,B型淀粉粒粒径快速增大,体积逐渐增大,新的小B型淀粉粒逐渐形成,C型淀粉粒产生。花后30 d至成熟,A型淀粉粒粒径、体积逐渐增加,而B、C型淀粉粒不断形成,并逐渐长大。青稞淀粉粒发育较为迟缓。此外,青稞花后1~25 d之间,淀粉和干物质积累速度均相对较低,这可能与西藏林芝地区特殊温度变化有关。在青稞花后25 d,即4月1日-4月25日青稞灌浆初期,高温13~20 ℃,低温1~6 ℃,日平均气温在8~13 ℃,在此期间,有22 d为阴雨天气,光合有效辐射低,青稞光合能力不足,导致干物质积累速度较慢,进而影响了淀粉积累和淀粉粒发育。环境条件对淀粉粒度分布有重要影响。例如高温使B型淀粉粒的数量下降,A型淀粉粒数量会增加[26-27]。水分充足条件下,B型淀粉粒表面积占比和体积占比会增加,而A型淀粉粒会减少。遮阳导致B型淀粉粒体积百分比显著减少,而A型淀粉粒增加[28]。西藏林芝地区低温多雨的环境可能导致青稞淀粉粒与遮阳条件下有相似的表现,即B型淀粉粒数量减少,A型淀粉粒数量增加,而在西藏拉萨、日喀则额、山南等日光充足、雨水较少地区,青稞籽粒B型淀粉粒数量可能会下降,而A型淀粉粒数量会增加。因此,对西藏青稞胚乳淀粉、淀粉粒还需要进一步研究。
[1]TANG H J,ANDO H,WATANABE K,etal.Physicochemical properties and structure of large,medium and small granule starches in fractions of normal barley endosperm [J].CarbohydrateResearch,2001,330(2):242.
[2] 冯继林,甲 干,向明华,等.藏区青稞考察与思考[J].大麦与谷类科学,2007(3):7.
FENG J L,JIA G,XIANG M H,etal.Investigation and thinking of hulless barley in Tibet [J].BarleyAndCerealSciences,2007(3):7.
[3]KANG M Y,SUGIMOTO K I,SAKAMOTO S,etal.Some properties of large and small starch granules of barley endosperm [J].AgricBiolChem,1985,49(5):1293.
[4]LI W,YAN S,YIN Y,etal.Starch granule size distribution in wheat grain in relation to shading after anthesis [J].JournalofAgriculturalScience,2010,148(2):183.
[5]WANG S J,LUO H,ZHANG J,etal.Alkali-induced changes in functional properties and in vitro digestibility of wheat starch:The role of surface proteins and lipids [J].JournalofAgriculturalandFoodChemistry,2014,62(16):3636.
[6]RAEKER M O,GAINES C S,FINNEY P L,etal.Granule size distribution and chemical composition of starches from 12 soft wheat cultivars [J].CerealChemistry,1998,75(5):721.
[7]LAMBROS F,ATHANASIA K,GEORGE K,etal.Study of the influence of surfactants on the size distribution and mass ratio of wheat starch granules by sedimentation/steric field-flow fractionation [J].FoodHydrocolloids,2008,22(6):961.
[8]TAKEDA Y,TAKEDA C,MIZUKAMI,etal.Structures of large,medium and small starch granules of barley grain [J].CarbohydratePolymers,1999,38(2):109.
[9]NI Y,WANG Z,YIN Y,etal.Starch granules size distribution in wheat grain in relation to phosphorus fertilization [J].JournalofAgriculturalScience,2012,150(1):45.
[10]LI W,SHAN Y,XIAO X,etal.Effect of nitrogen and sulfur fertilization on accumulation characteristics and physicochemical properties of A- and B-wheat starch [J].JournalofAgriculturalandFoodChemistry,2013,61(10):2418.
[11]AHMADI A,BAKER D A.The effect of water stress on the activities of key regulatory enzymes of the sucrose to starch pathway in wheat[J].PlantGrowthRegulation,2001,35(1):81.
[12]YU A,LI Y,NI Y,etal.Differences of starch granule distribution in grains from different spikelet positions in winter wheat [J].PlosOne,2014,9(12):e114342.
[13]WEI C X,ZHANG J,ZHONG F X,etal.Comparison of the starch granule and kernel structure between feed and malt barley varieties [J].JournalofTriticeaeCrops,2006,26(4):133.
[14]SALMAN H,BLAZEK J,LOPEZ R A,etal.Structure- function relationships in A and B granules from wheat starches of similar amylose content [J].CarbohydratePolymers,2009,75(3):420.
[15]LI J H,VASANTHAN T,HOOVER RROSSNAGEL B G,etal.Starch from hull-less barley:IV.Morphological and structural changes in waxy,normal and high-amylose starch granules during heating[J].FoodResearchInternational,2004,37(5):417.
[16]KIM H S,HUBER K C.Impact of A/B-type granule ratio on reactivity,swelling,gelatinization,and pasting properties of modified wheat starch.Part I:Hydroxypropylation [J].CarbohydratePolymers,2010,80(1):94.
[17]TILLETT I J L,BRYCE J H.The Regulation of Starch Granule Sizein Endosperm of Developing Barley Grains [M].Oxford:Oxford University Press,1994:45.
[18]AO Z,JANE J.Characterization and modeling of the A- and B-granule starches of wheat,triticale,and barley [J].CarbohydratePolymers,2007,67(1):46.
[19]BECHTEL D B,WILSON J D.Amyloplast formation and starch granule development in hard red winter wheat [J].CerealChemistry,2007,80(2):175.
[20]PENG M,HUCL P,CHIBBAR R N.Isolation,characterization and expression analysis of starch synthase I from wheat(TriticumaestivumL.)[J].PlantScience,2001,161(6):1055.
[21]JAISWAL S,BAGA M,AHUJA G,etal.Development of barley(HordeumvulgareL.)lines with altered starch granule size distribution[J].JournalofAgriculturalandfoodChemistry,2014,62(10):2289.
[22]LI J H,VASANTHAN T,ROSSNAGEL B,etal.Starch from hull-less barley:I.Granule morphology,composition and amylopectin structure [J].FoodChemistry,2001,74(4):395.
[23]SHINDE S V,NELSON J E,HUBER K C.Soft wheat starch pasting behavior in relation to A-and B-type granule content and composition [J].CerealChemistry,2003,80(1):91.
[24]LI W,YAN S,SHI X,etal.Starch granule size distribution from twelve wheat cultivars in east China’s Huaibei region[J].CanadianJournalofPlantScience,2016,96(2):176.
[25]SINGH S,SINGH N,ISONO N,etal.Relationship of granule size distribution and amylopectin structure with pasting,thermal,and retrogradation properties in wheat starch[J].JournalofAgriculturalandFoodChemistry,2010,2010,58(2):1180.
[26]YAN S H,YIN Y P,LI W Y,etal.Effect of high temperature during grain filling on starch accumulation,starch granule distribution,and activities of related enzymes in wheat grains [J].ActaAgronomicaSinica,2008,34(6):1092.
[27]DAI Z,YIN Y,WANG Z.Starch granule size distribution from seven wheat cultivars under different water regimes[J].CerealChemistry,2009,86(1):82.
[28]AML A D,STARK J R,MORRISON W R,etal.The composition of starch granules from developing barley genotypes[J].JournalofCerealScience,1991,13(1):93
DevelopmentalCharacteristicsofStarchGranuleinEndospermofTibetanWinterHullessBarley
HOUWeihai,WANGJianling,DANBa,HUDang
(Xizang Agiricultural and Animal Husbandry College,Linzhi,Tibet 860000,China)
时间:2017-09-13
网络出版地址:http://kns.cnki.net/kcms/detail/61.1359.S.20170913.1139.028.html
S512.3;S311
A
1009-1041(2017)09-1232-08
2017-04-09
2017-05-05
国家自然科学基金项目(No.31560362); 西藏自治区重点研究项目(No.XZXTCX-2016)
E-mail:418267205@qq.com
王建林(E-mail:xzwangjl@126.com)
猜你喜欢
小学生学习指导(高年级)(2024年4期)2024-05-07 03:27:10
作物学报(2022年1期)2022-11-05 08:06:34
河北科技师范学院学报(2022年2期)2022-08-26 08:55:32
农业工程学报(2022年7期)2022-07-09 06:16:32
中国农业大学学报(2020年8期)2020-07-22 07:57:28
翠苑(2019年4期)2019-11-11 04:27:26
今日农业(2019年11期)2019-08-15 00:56:32
小学阅读指南·高年级版(2016年9期)2016-10-31 05:23:01
西藏科技(2016年8期)2016-09-26 09:00:24
中学生数理化·高二版(2008年11期)2008-06-17 09:25:42
