
念青唐古拉山南坡高寒草甸生产力对温度和降水变化的敏感性及其海拔分异
2017-10-13 12:27赵景学罗天祥
生态学报 2017年17期
李 翔,王 忠,赵景学,罗天祥
1 中国科学院青藏高原研究所高寒生态学与生物多样性重点实验室, 北京 100101 2 中国科学院大学, 北京 100049 3 武汉大学生命科学学院, 武汉 430072
念青唐古拉山南坡高寒草甸生产力对温度和降水变化的敏感性及其海拔分异
李 翔1,2,王 忠3,赵景学1,2,罗天祥1,*
1 中国科学院青藏高原研究所高寒生态学与生物多样性重点实验室, 北京 100101 2 中国科学院大学, 北京 100049 3 武汉大学生命科学学院, 武汉 430072
目前人们仍不清楚不同海拔高寒草地植被生长对气候变化的敏感性差异及其与最适宜海拔分布中心的关系。利用西藏当雄县念青唐古拉山南坡7个海拔梯度固定样地的高山嵩草草甸地上净初级生产力(ANPP)观测数据(2009—2013),建立了ANPP与同期遥感植被指数(MODIS NDVI)的线性回归方程。基于长时间序列的NDVI数据,利用建立的回归方程估算了研究区2000—2013年的ANPP。结合沿海拔梯度的HOBO气象站数据(2006—2013)及当雄县气象站数据(2000—2013),分析了2000—2013年该地区高寒草甸ANPP对降水和温度变化的敏感性及其随海拔的变化规律。结果表明:(1)多年平均ANPP随海拔的变化均表现为先增加后降低的单峰分布格局,最大值出现在海拔4893—4942 m,说明在海拔梯度上存在一个最适宜高寒草甸植被生长的分布中心;(2)ANPP与生长季降水量(GSP)呈正相关关系,与生长季平均气温(GST)呈负相关关系,其相关斜率的绝对值(指示ANPP的降水敏感性和温度敏感性大小)与ANPP的海拔格局具有相反的变化趋势,即在最适宜高寒草甸植被生长的海拔分布中心附近,ANPP对降水和温度变化的敏感性最低,而在远离该分布中心的较高和较低海拔,ANPP对降水和温度变化的敏感性则相对较大。研究明确了高寒草甸ANPP对降水和温度变化的敏感性随海拔的分异性及其与高寒草甸最适宜海拔分布中心的关系,这有助于理解沿海拔梯度不同水热组合环境下高寒生态系统对未来气候变化的响应模式。
净初级生产力;敏感性;海拔梯度;高寒草甸;气候变化
Abstract: It remains unclear whether and how the sensitivity of alpine plant growth to climate change may vary with distance away from the altitudinal distribution center of a vegetation type. On the basis of measurements of aboveground net primary productivity (ANPP) in an alpineKobresiameadow across 7 altitudes along the southern slope of Nyainqentanglha Mountains in Tibet during 2009—2013, we established a linear regression model between ground ANPP and satellite vegetation index (MODIS NDVI). Using the long time-series MODIS NDVI data, we then applied the established model to estimate ANPP in the study area during 2000—2013. In combination with climatic data obtained from altitudinal HOBO weather stations (2006—2013) and Damxung weather station (2000—2013), we aimed to clarify the altitudinal pattern in the sensitivity of ANPP to changes in temperature and precipitation in alpine meadows. A unimodal pattern in multi-year mean ANPP was observed along the altitudes; ANPP increased with increasing altitude up to 4893—4942 m and then decreased, demonstrating the altitudinal distribution center of the alpine meadow at mid-altitude. ANPP was positively correlated with growing season precipitation but negatively correlated with growing season mean air-temperature during 2000—2013. The absolute values of the linear regression slopes (referred to as the sensitivity of ANPP to changes in precipitation and temperature) showed a pattern opposite to the change in ANPP along the altitudes. The precipitation and temperature sensitivities of ANPP were the lowest around the distribution center but higher at lower or higher altitudes with distance away from the distribution center. In conclusion, our data identified the altitudinal pattern in sensitivity of alpine meadow ANPP to climate change with a link to the altitudinal distribution center of the alpine meadow. Such knowledge is important for understanding the response of alpine meadow ecosystems to future climate change in different environments along an altitudinal gradient.
KeyWords: net primary productivity; sensitivity; altitudinal gradient; alpine meadow; climate change
在过去100年里,全球气温普遍升高,并预测未来将持续加剧[1]。然而,与温度普遍升高的趋势不同,降水的变化趋势存在很大的时空差异[2],从而导致不同地区和不同时段的温度和降水组合发生很大变化,这无疑会增加干旱半干旱地区生态系统生产力变化的复杂性。植被净初级生产力(NPP)是指某一段时间内单位面积上绿色植物总光合生产力减去其本身呼吸消耗之后剩余的部分[3-4]。NPP是生态系统功能的重要指标和碳循环的重要组成部分[5- 7],反映了气候变化和人类活动对陆地植被综合作用的结果[8]。在众多气候因子中,温度和降水及其组合是影响NPP时空变化的主要气候因子[9- 13],且不同区域NPP受到的主要限制因子也有所不同[13-14]。一个生态系统的脆弱性往往取决于生态系统对气候变化的敏感性和适应性(脆弱性=适应性‐敏感性)[1, 15]。因此,理解NPP对温度和降水变化的敏感性及其地理分异性将有助于我们科学评估陆地生态系统对未来气候变化的敏感性和脆弱性[1]。
通常认为高海拔地区的生态系统对气候变化的响应更为敏感[16-17]。海拔梯度在较小的地理范围内引起环境因子(如温度、降水等)发生较大的变化[18],这为我们研究不同水热组合环境下NPP对温度和降水变化的敏感性差异提供了理想的天然实验室。一般来说,在海拔跨度较大的情况下,随海拔的升高,温度逐渐降低而降水呈增加的趋势[13-14],这种变化趋势导致在不同海拔上具有不同的水热组合,在一定海拔高度存在一个最适宜植被生长和分布的水热组合环境。已有的研究表明,在干旱半干旱地区,高寒植被生长一般在低海拔受到干旱胁迫,而在高海拔出现低温胁迫,优势或关键物种盖度、群落物种丰富度及植被NPP等表现为随海拔先增加,达到一个峰值后再降低的单峰格局[13-14]。这说明植被生长和分布的环境限制因子沿海拔梯度发生变化,其对温度和降水变化的响应方式很可能存在差异。然而,目前尚不清楚在沿一个海拔梯度上,不同水热组合条件下NPP对温度和降水变化的敏感性具有怎样的变化特征。
作为世界第三极,青藏高原具有极为独特而脆弱的生态环境,其主体生态系统长期处于生存环境阈值下,对气候变化的响应具有高度的敏感性和脆弱性[16- 20]。位于高原中部的念青唐古拉山中段地区,地处高寒草甸到高寒草原过渡区,属半干旱气候区[21]。这里海拔高差大,从低海拔到高海拔气候干旱度逐渐降低,高寒草甸分布存在明显的海拔上限和下限,因而该地区为明确上述科学问题提供了很好的研究平台。近几十年来,遥感技术的迅速发展为在景观和区域尺度上研究植被生长的长期变化提供了可能。长时间序列的归一化植被指数(NDVI)能够反映植被生长的季节和年际变化特征[22],因而被广泛运用于分析植被生长状况的时空变化及其与气候变化的关系[23- 26]。
在这一研究中,系统收集整理了西藏当雄县念青唐古拉山南坡7个海拔梯度固定样地的高寒草甸地上最大生物量(即地上NPP)实测数据、沿海拔梯度的短期气象观测数据、当雄县气象站长期观测数据以及长时间序列MODIS NDVI数据,旨在阐明2000—2013年沿海拔梯度高寒草甸NPP对降水和温度变化的敏感性差异及其与最适宜海拔分布中心的关系,为理解不同水热组合环境下高寒植被生产力对气候变化的响应规律及评估在未来气候变化背景下高寒草地生态系统的脆弱性提供科学依据。
1 数据与方法
1.1 研究区概况
研究区域位于西藏自治区当雄县境内,地处青藏高原中部念青唐古拉山南坡,地理位置为30°23′ — 30°33′ N,90°57′ — 91°08′ E(图1)。该区域最高海拔约为5800 m,草线位置大致在海拔5200 m,从山脚当雄县气象站(4288 m)到峰顶的海拔跨度达1500 m。该地区气候属于高原亚寒带季风半干旱气候[21],雨热同期,干湿季明显。根据当雄县气象站记载(1963—2013),年平均气温为1.8℃,最冷月(1月)平均气温-9℃,最热月(7月)平均气温11.1℃。同期平均年降水量为442mm,90%的降水集中在5—9月植被生长季期间。50多年以来,该地区气温总体呈显著升高趋势,且最近十几年来升温速率明显加快。年平均气温、生长季平均气温和非生长季平均气温在1963—1999年分别以0.29、0.19、0.36℃/10a的速率升高,而在2000—2013年分别为0.66、0.96、0.44℃/10a。降水量在这两个时段的变化规律有所不同,1963—1999年间,全年降水量、生长季降水量和非生长季降水量均呈增加趋势,但生长季降水量的年际变化不显著;在2000—2013年间,全年降水量和生长季降水量均呈减少趋势,非生长季降水量略呈增加趋势,其中全年降水量和非生长季降水量的年际变化并不显著。基于沿海拔梯度HOBO自动气象站记录(2006—2013),随着海拔升高,温度逐渐降低,而降水表现为先增加后降低的格局,峰值出现在海拔5100 m左右[13]。研究区域处于高寒草甸向高寒草原的过渡带[27],植被类型以高寒草甸为主,较高海拔(4700—5200 m)是以高山嵩草(Kobresiapygmaea)和垫状点地梅(Androsacetapete)为优势种的高寒草甸,较低海拔(4300—4650 m)是以丝颖针茅(Stipacapillacea)和矮生嵩草(Kobresiahumilis)为优势种的草原化草甸,在海拔4300 m以下地带主要是由河流与低洼地形成的沼泽湿地。
1.2 样地和样带的布设
于2006年在念青唐古拉山南坡(阳坡)沿海拔梯度建立了7个20 m×20 m固定样地,海拔跨度包括草甸的上、下限,高度分别为4390、4500、4640、4800、4950、5110、5210 m(图1)。在2009年,在每个固定样地机械布设了6个1 m×1 m的定位观测样方,每年7—8月调查样方内的物种组成、盖度和高度等。
坡向是重要的地形因子之一,其通过影响光照、温度及降水的空间分配进而影响到植物群落的组成与分布[28]。为理解坡向是否会影响植被生产力对温度和降水变化的敏感性及其海拔变化格局,并且考虑到固定样地实测数据的代表性,本研究着重选择3条宽2 km的海拔梯度样带进行剖析(图1),海拔跨度包括草甸的上、下限。样带1设置在7个固定样地所在的坡面,海拔范围4350—5250 m;样带2设置在样带1以西海拔跨度更大的坡面,海拔范围4350—5400 m;样带3设置在阳坡(南坡)对面的阴坡(北坡),海拔范围4250—5250 m,其最高海拔低于南坡,草线位置几乎接近峰顶。

图1 研究区地理位置、固定样地以及选取的3条海拔样带分布图Fig.1 Location of the study area, sampling plots and the three selected altitudinal transects
1.3 数据来源
1.3.1 固定样地最大生物量的测定
将各海拔固定样地的1 m×1 m固定样方划分为4个0.5 m×0.5 m的小样方,然后进一步划分为0.1 m×0.1 m的小格子,以小格子为单位进行群落调查(每年7—8月),定位观测物种组成、高度和盖度等变化。作为长期定位观测研究平台,固定样方的地上生物量采用基于盖度×高度的生物量体积回归方程法进行估测,以避免人为干扰并确保数据的连续性和可比性。为此,在每一海拔固定样地外另选取3个1 m×1 m样方,对每一样方左上角的0.5 m×0.5 m的小样方进行群落调查并以收获法实测地上生物量(用剪刀贴地表剪取植物地上部分,不包括枯死物),按物种建立实测生物量(干重)与盖度×高度的经验方程(综合7个海拔测定数据,具体方法和经验方程详见文献[13-14])。基于固定样方的物种组成、高度和盖度等连续观测数据(按0.5 m×0.5 m的小样方),利用已建立的生物量体积经验方程估算了2009—2013年7个海拔样地的地上最大生物量及其变化。由于研究区优势物种地上部分均为1年生,且群落调查和生物量收集时间都在其最大生物量时期,将地上最大生物量视为地上净初级生产力(Aboveground Net Primary Productivity, ANPP)[13-14]。
1.3.2 气象观测数据
沿海拔梯度气象观测数据来自每个固定样地旁边的Onset HOBO小型自动气象站记录(于2005年8月架设),数据采集间隔为每30min 1次,其中气温数据记录时间范围为2005年8月到2013年12月,降水数据记录时间范围为2006年7月至2013年12月。另外,我们还获取了当雄县气象站(距离4390m固定样地4 km,海拔4288m)的气温和降水数据(来自中国气象科学数据共享服务网:http://cdc.nmic.cn/home.do),时间分辨率为日值,时间范围为2000年1月至2013年12月。
1.3.3 NDVI和DEM高程数据
归一化植被指数(NDVI)被定义为近红外波段的反射值与红光波段的反射值之差与两者之和的比值,能够反映植被生长的季节和年际变化特征[22]。本研究使用的NDVI数据来自NASA官网LAADS Web数据中心(https://ladsweb.nascom.nasa.gov/data/)提供的16d最大值合成的MODIS植被指数产品(MODIS/Terra Vegetation Indices 16-Day L3 Global 250m SIN Grid,MOD13Q1),空间分辨率为250m,数据质量等级为5级,数据格式为EOS-HDF,时间覆盖为2000—2013年。使用MRT(MODIS Reprojection Tools)软件将下载的MOD13Q1数据产品进行格式和地图投影转换,把HDF格式转换为TIF格式,并将SIN投影转换为Albers等面积割圆锥投影。DEM高程数据来自SRTM DEM(数据来源于中国科学院计算机网络信息中心国际科学数据镜像网站(http://www.gscloud.cn)),分辨率为90m。
1.4 数据处理与分析
1.4.12000—2013年ANPP的估算

图2 2009—2013年沿海拔梯度NDVI与ANPP的回归关系 Fig.2 Relationship between NDVI and ANPP along an altitudinal gradient during 2009—2013误差线为标准误
利用ArcGIS软件,提取出2009—2013年7个海拔对应样点的NDVI值。为了保持NDVI值与实测的地上生物量在时间上一致,分别采用2009—2013年第209天(7月28至8月12日最大值合成)的NDVI作为回归分析值。以每个海拔6个固定样方的地上生物量平均值作为该海拔的地上净初级生产力,记为ANPP。如图2所示,NDVI与ANPP表现为非常密切的线性关系(R2=0.80,P< 0.001,n=35),因而建立NDVI与ANPP的线性回归模型为:
ANPP=742.04×NDVI-79.42
每年最大NDVI值代表了植被生长盛期的信息。通过建立的回归模型,利用年最大NDVI值估算了整个研究地区过去14a(2000—2013)的地上净初级生产力。
1.4.2 ANPP对降水和温度变化的敏感性分析
基于估算的2000—2013年ANPP,统计分析ANPP对降水和温度变化的敏感性。气候因子选取与植被生长关系最为密切的生长季降水量(GSP)和生长季平均气温(GST),其中生长季定义为5—9月。通过比较分析发现,2006—2013年沿海拔梯度自动气象站观测的生长季降水量和平均气温的年际变化与当雄县气象站观测数据具有很好的一致性(图3),说明在距离当雄县气象站一定范围内,降水和气温都应该表现为相同的年际变化趋势。因此,在整个研究区范围内(距离当雄县气象站都不超过15km),可以利用当雄县气象站长期观测的气象数据来分析不同海拔ANPP对降水和温度变化的敏感性。
敏感性大小采用ANPP与气候因子(温度、降水)之间线性回归方程的斜率来表示,斜率绝对值表示ANPP对气候因子变化的敏感性大小,正、负斜率分别表示ANPP对气候因子的变化表现为正响应、负响应。
根据当雄县气象站观测数据,利用编写的IDL程序,计算研究区内每个像元ANPP与GSP和GST的线性回归斜率以及多年平均(2000—2013年)ANPP。利用ArcGIS 10.3软件,分别统计出3个海拔样带内每个像元的多年平均ANPP、回归斜率以及像元中心对应的DEM海拔高程值。利用三次多项式方程(y=ax3+bx2+cx+d),分别拟合了ANPP与海拔的非线性关系以及ANPP-GSP和ANPP-GST的关系斜率与海拔的相关模式。根据拟合的多年平均ANPP随海拔的变化曲线方程,利用求导数的方法找到曲线极大值所对应的海拔高度,并以此海拔作为高寒草甸最适宜海拔分布中心,该分布中心代表了高寒草甸生长最适宜的海拔地带。

图3 2006—2013年沿海拔梯度HOBO气象站观测数据与当雄县气象站观测数据的年际变化趋势比较Fig.3 Comparisons of interannual trends in growing season precipitation and growing season mean air-temperature obtained from altitudinal HOBO weather stations (2006—2013) and Damxung weather station (2000—2013)
2 结果与分析
2.1 ANPP的海拔分异规律
图4显示,随着海拔的升高,固定样地的实测ANPP与同期NDVI值均表现为先升高后降低的单峰格局,不同年份ANPP一般在海拔4950 m达到最大值(除了2012年外)。与ANPP海拔格局类似,NDVI随着海拔升高也呈先升高后降低的格局,不同年份NDVI最大值均出现在海拔4950 m。
在3个海拔梯度样带中,三次多项式方程很好地拟合了多年平均ANPP沿海拔梯度的变化,其中样带1海拔变化对ANPP变化的解释度最高,达到85%,其次分别是样带2和样带3,解释度分别为70%和48%(图5)。从拟合曲线可以看出,随着海拔的升高,多年平均ANPP呈现出先升高后降低的单峰格局(图5),这与固定样地的实测ANPP海拔变化格局一致。如图5所示,虚线表示ANPP随海拔的拟合曲线最大值出现的海拔高度,该海拔可视为高寒草甸最适宜海拔分布中心。样带1、2、3的高寒草甸最适宜海拔分布中心分别出现在海拔4893、4942、4856 m。
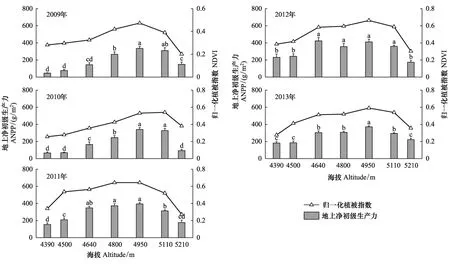
图4 2009—2013年实测ANPP和NDVI的海拔变化格局Fig.4 Altitudinal variations in observed ANPP and NDVI during 2009—2013不同字母表示不同海拔之间ANPP差异显著(P < 0.05),误差线为标准误
2.2 ANPP的降水和温度敏感性随海拔的变化规律
由当雄县气象站观测数据的年际变化趋势可知(图3),2000—2013年间,该区域GSP年际变化呈显著降低趋势(P< 0.05),GST呈显著增加趋势(P< 0.01)。3个海拔梯度样带ANPP与GSP呈正相关关系,其降水回归斜率为正值(图5),而ANPP与GST呈负相关关系,其温度回归斜率为负值(图5)。在高寒草甸最适宜海拔分布中心以下的低海拔地区,占81%的像元ANPP与GSP和GST关系达到显著相关(P< 0.05)或弱相关(P< 0.10),而在分布中心以上高海拔地区,达到显著相关(P< 0.05)或弱相关(P< 0.10)的像元数则相对较少(44%)(图5)。
3个海拔梯度样带均表现为在高寒草甸最适宜海拔分布中心附近,ANPP与降水和温度的回归斜率的绝对值最低,而在远离该分布中心的较高或较低海拔,其回归斜率的绝对值逐渐增大。综上所述,ANPP对降水和温度变化的敏感性与多年平均ANPP在海拔格局上具有相反的变化趋势,即在高寒草甸最适宜海拔分布中心附近,ANPP对降水和温度变化的敏感性最低,而在远离该分布中心的较高和较低海拔,ANPP对降水和温度变化的敏感性则相对较大。

图5 3条海拔样带多年平均ANPP、ANPP分别与GSP和GST的回归斜率随海拔的变化格局。Fig.5 Altitudinal variations in multi-year mean ANPP and the regression slopes of ANPP vs GSP and ANPP vs GST along the three altitudinal transects深灰色和浅灰色填充圆圈分别表示显著水平达到0.05和0.10,虚线表示高寒草甸的海拔分布中心
进一步分析了3个海拔梯度样带内所有像元ANPP的降水和温度敏感性与多年平均ANPP的关系。如图6所示,3条样带ANPP-GSP回归斜率与多年平均ANPP都呈显著负相关关系(P< 0.001),而ANPP-GST回归斜率与多年平均ANPP都呈显著正相关关系(P< 0.001)。因此,从敏感性的角度看,ANPP的降水和温度敏感性(回归斜率的绝对值)与ANPP均呈显著负相关关系(P< 0.001),即随着ANPP的升高,ANPP对降水和温度变化的敏感性逐渐降低。

图6 3条海拔样带多年平均ANPP分别与其降水回归斜率和温度回归斜率之间的关系。Fig.6 Relationships of multi-year mean ANPP to the regression slopes of ANPP vs GSP and ANPP vs GST along the three altitudinal transects
3 讨论
多数情况下,随海拔升高,温度降低而降水增加[18]。在干旱半干旱地区,水热组合因子沿海拔梯度的变化控制着高寒草地优势或关键物种的生长和分布[13-14, 29- 31]。因此,不同海拔高度的植被对气候变化的响应方式应该有所不同。本文研究结果显示,固定样地的实测ANPP与同期NDVI值随海拔均表现为先升高后降低的单峰格局(图4),3个海拔梯度样带的多年平均ANPP随海拔同样表现出先升高后降低的单峰格局(图5),说明在海拔梯度上存在一个最适宜高寒草甸植被生长的分布中心。ANPP对降水和温度变化的敏感性在这个分布中心附近最低,而远离该分布中心的较高和较低海拔则相对较大(图5)。
根据限制因子作用律,植被生物量和生产力一般在最适宜环境条件下达到最大,在远离该适宜区时呈降低趋势[3, 32-33]。植物的生长和分布依赖于各环境因子的综合作用,并且必然受到一种或几种关键因子的限制。在干旱半干旱地区,植被生产力的限制因子随海拔会发生转变。在低海拔地区,NPP主要受到低降水导致的干旱胁迫[13-14],同时相对较高的温度导致蒸散增加进一步加剧了植物的水分胁迫[34- 36]。在较高海拔地区,低温也会导致植物生长普遍出现水分或养分限制[37-38],从而导致较低的生产力[13]。在生理生态机制上,当土壤温度低于7℃时,叶同化CO2的日总量显著下降;而当土壤温度大于7℃时,植物光合生产力对温度的敏感性显著下降[39]。在本研究区念青唐古拉山南坡,Wang等[13]研究发现,水热组合因子决定了生产力沿海拔梯度的单峰分布格局,在海拔梯度上存在最适宜植被生长的水热组合环境,其生长季平均气温和土壤温度分别为5.4—5.9℃和7.4—8.5℃,水热组合指标即生长季降水与积温(≥5℃)的比值为0.80—0.84,与光合生产力机理模型一致[38]。根据联系叶氮含量和碳稳定同位素与土壤水量平衡的光合生产力机理模型,当植被蒸腾与蒸散量比值小于0.85时,土壤水分供给是生产力的主要限制因子,而当植被蒸腾与蒸散量比值大于该阈值时,土壤氮素养分供给是生产力的主要限制因子[38]。因此,在最适宜海拔分布中心附近,植物生长具有最适宜的水热组合环境,其所面临的环境胁迫相对较小,水热因子的年际波动很少会超出其最适宜的水热组合范围,因而其对水热因子年际变化的敏感性相对较低。而在远离最适宜海拔分布中心的高海拔和低海拔时,植物生长受到的环境胁迫较大(如低海拔地区高温和降水较少导致干旱胁迫,高海拔地区低温导致水分和养分胁迫),因而对水热因子年际变化的响应更为敏感。
此外,从物种多样性的角度看,一个高度稳定的生态系统往往同时具有较高的物种丰富度[40]。由于共存物种对限制资源的利用存在互补性,群落物种丰富度增加会增强生态系统的稳定性[41- 43]。在本研究区此前的研究结果显示,物种丰富度与ANPP的海拔格局类似,随海拔升高也呈先升高后降低的单峰分布格局,两者具有相同的海拔分布中心[13-14]。因此,在最适宜海拔分布中心,物种丰富度最高且ANPP最大,生态系统结构相对稳定,对环境变化的敏感性较低。
对比不同坡向3个海拔梯度样带的结果,坡向并没有改变植被生产力对温度和降水变化的敏感性随海拔的变化格局(图5),这可能与季风气候效应有关。青藏高原高寒草甸和草原是适应寒冷和干旱气候的高原地带性植被,其形成和分布深受印度季风气候效应的影响。在西风与季风交互作用下,生长季早期既寒冷又干旱。前期研究发现,在雨热同期的季风气候效应下,为避免春季低温和干旱胁迫,高寒草地广布优势物种普遍形成一种能感应季风雨来临时间的返青物候适应策略[29],草地植被生长主要受生长季降水的控制[20]。由于印度洋水汽主要沿雅鲁藏布江大峡谷通道向高原内陆输送,同一峡谷相对分布的南坡与北坡具有相似的生长季降水变化格局[28]。因此,高寒草地生产力对季风降水的敏感性及其沿海拔梯度的单峰分布格局应该具有普遍性。研究结果进一步显示,ANPP对降水和温度的敏感性(其回归关系斜率的绝对值)与ANPP呈显著负相关关系(图6),即ANPP越大,其对温度和降水变化的敏感性越低,这与两者在海拔梯度上的分布格局是一致的。该结果指出,在植被生长状况较差的地区,高寒草地生态系统受到的环境胁迫较大,其对气候变化的响应更加敏感和脆弱。
据当雄县气象站记录,研究区最近14年(2000—2013年)生长季平均气温显著升高,生长季降水量显著减少,气候出现暖干化趋势(图3)。气候变暖可能会减缓高海拔地区植被生长的低温胁迫,但同时也会进一步加剧其在低海拔的干旱胁迫,进而可能使高寒嵩草草甸最适宜海拔分布中心沿海拔发生上移[13]。在嵩草草甸海拔分布中心以下的低海拔地区,降水减少和温度升高共同加剧了干旱胁迫,ANPP的降低趋势更加明显(图5)。因此,在暖干化气候变化背景下,高寒草甸生态系统将变得更加脆弱,并将加剧人类活动引起的草地退化[44- 46]。
4 结论
本文分析了西藏当雄县念青唐古拉山南坡高寒草甸生产力对降水和温度变化的敏感性及其随海拔的变化规律。研究结果表明:
(1)ANPP随海拔的变化整体上均表现为先增加后降低的单峰分布格局,表明在海拔梯度上存在一个最适宜高寒草甸植被生长的分布中心;
(2)ANPP对降水和温度变化的敏感性在最适宜的海拔分布中心附近最低,而在远离该分布中心的较高和较低海拔则相对较大。
研究结果揭示了不同海拔高寒草甸生产力对降水和温度变化的敏感性差异及其与最适宜海拔分布中心的关系,这有助于理解沿海拔梯度不同水热组合环境下高寒生态系统对未来气候变化的响应模式。
[1] IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Cambridge: Cambridge University Press, 2014.
[2] Bradley R S, Diaz H F, Eischeid J K, Jones P D, Kelly P M, Goodess C M. Precipitation fluctuations over northernhemisphere land areas since the mid- 19th century. Science, 1987, 237(4811): 171- 175.
[3] 罗天祥. 中国主要森林类型生物生产力格局及其数学模型[D]. 北京: 中国科学院研究生院(国家计划委员会自然资源综合考察委员会), 1996.
[4] 周广胜, 王玉辉. 全球生态学. 北京: 气象出版社, 2003.
[5] Guo R, Wang X K, Ouyang Z Y, Li Y N. Spatial and temporal relationships between precipitation and ANPP of four types of grasslands in northern China. Journal of Environmental Sciences, 2006, 18(5): 1024- 1030.
[6] Houghton R A, Hall F, Goetz S J. Importance of biomass in the global carbon cycle. Journal of Geophysical Research-Biogeosciences, 2009, 114(G2): G00E03.
[7] Loreau M, Naeem S, Inchausti P, Bengtsson J, Grime J P, Hector A, Hooper D U, Huston M A, Raffaelli D, Schmid B, Tilman D, Wardle D A. Biodiversity and ecosystem functioning: current knowledge and future challenges. Science, 2001, 294(5543): 804- 808.
[8] 张镱锂, 祁威, 周才平, 丁明军, 刘林山, 高俊刚, 摆万奇, 王兆锋, 郑度. 青藏高原高寒草地净初级生产力(NPP)时空分异. 地理学报, 2013, 68(9): 1197- 1211.
[9] Luo T X, Li W H, Zhu H Z. Estimated biomass and productivity of natural vegetation on the Tibetan Plateau. Ecological Applications, 2002, 12(4): 980- 997.
[10] Luo T X, Pan Y D, Ouyang H, Shi P L, Luo J, Yu Z L, Lu Q. Leaf area index and net primary productivity along subtropical to alpine gradients in the Tibetan Plateau. Global Ecology and Biogeography, 2004, 13(4): 345- 358.
[11] Knapp A K, Smith M D. Variation among biomes in temporal dynamics of aboveground primary production. Science, 2001, 291(5503): 481- 484.
[12] Huxman T E, Smith M D, Fay P A, Knapp A K, Shaw M R, Loik M E, Smith S D, Tissue D T, Zak J C, Weltzin J F, Pockman W T, Sala O E, Haddad B M, Harte J, Koch G W, Schwinning S, Small E E, Williams D G. Convergence across biomes to a common rain-use efficiency. Nature, 2004, 429(6992): 651- 654.
[13] Wang Z, Luo T X, Li R C, Tang Y H, Du M Y. Causes for the unimodal pattern of biomass and productivity in alpine grasslands along a large altitudinal gradient in semi-arid regions. Journal of Vegetation Science, 2013, 24(1): 189- 201.
[14] 王忠. 念青唐古拉山南坡高寒草甸生产力沿海拔梯度的变化机理研究[D]. 北京: 中国科学院研究生院, 2011.
[15] Young G, Zavala H, Wandel J, Smit B, Salas S, Jimenez E, Fiebig M, Espinoza R, Diaz H, Cepeda J. Vulnerability and adaptation in a dryland community of the Elqui Valley, Chile. Climatic Change, 2010, 98(1/2): 245- 276.
[16] Yao T D, Thompson L G, Mosbrugger V, Zhang F, Ma Y M, Luo T X, Xu B Q, Yang X X, Joswiak D R, Wang W C, Joswiak M E, Devkota L P, Tayal S, Jilani R, Fayziev R. Third pole environment (TPE). Environmental Development, 2012, 3: 52- 64.
[17] Diaz H F, Grosjean M, Graumlich L. Climate variability and change in high elevation regions: past, present and future. Climatic Change, 2003, 59(1/2): 1- 4.
[18] Körner C. The use of ′altitude′ in ecological research. Trends in Ecology & Evolution, 2007, 22(11): 569- 574.
[19] Liu X D, Chen B D. Climatic warming in the Tibetan Plateau during recent decades. International Journal of Climatology, 2000, 20(14): 1729- 1742.
[20] Zhao J X, Luo T X, Li R C, Li X, Tian L H. Grazing effect on growing season ecosystem respiration and its temperature sensitivity in alpine grasslands along a large altitudinal gradient on the central Tibetan Plateau. Agricultural and Forest Meteorology, 2016, 218- 219: 114- 121.
[21] 中国科学院青藏高原综合科学考察队. 西藏气候. 北京: 科学出版社, 1984.
[22] Huete A, Didan K, Miura T, Rodriguez E P, Gao X, Ferreira L G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sensing of Environment, 2002, 83(1/2): 195- 213.
[23] Ding M J, Zhang Y L, Liu L S, Zhang W, Wang Z F, Bai W Q. The relationship between NDVI and precipitation on the Tibetan Plateau. Journal of Geographical Sciences, 2007, 17(3): 259- 268.
[24] Mao D H, Wang Z M, Luo L, Ren C Y. Integrating AVHRR and MODIS data to monitor NDVI changes and their relationships with climatic parameters in Northeast China. International Journal of Applied Earth Observation and Geoinformation, 2012, 18: 528- 536.
[25] Pettorelli N, Vik J O, Mysterud A, Gaillard J M, Tucker C J, Stenseth N C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends in Ecology & Evolution, 2005, 20(9): 503- 510.
[26] Piao S L, Nan H J, Huntingford C, Ciais P, Friedlingstein P, Sitch S, Peng S S, Ahlström A, Canadell J G, Cong N, Levis S, Levy P E, Liu L L, Lomas M R, Mao J F, Myneni R B, Peylin P, Poulter B, Shi X Y, Yin G D, Viovy N, Wang T, Wang X H, Zaehle S, Zeng N, Zeng Z Z, Chen A P. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nature Communications, 2014, 5: 5018.
[27] 中国科学院青藏高原综合科学考察队. 西藏植被. 北京: 科学出版社, 1988.
[28] Liu X S, Luo T X. Spatiotemporal variability of soil temperature and moisture across two contrasting timberline ecotones in the Sergyemla Mountains, southeast Tibet. Arctic, Antarctic, and Alpine Research, 2011, 43(2): 229- 238.
[29] Li R C, Luo T X, Mölg T, Zhao J X, Li X, Cui X Y, Du M Y, Tang Y H. Leaf unfolding of Tibetan alpine meadows captures the arrival of monsoon rainfall. Scientific Reports, 2016, 6: 20985.
[30] Li R C, Luo T X, Tang Y H, Du M Y, Zhang X Z. The altitudinal distribution center of a widespread cushion species is related to an optimum combination of temperature and precipitation in the central Tibetan Plateau. Journal of Arid Environments, 2013, 88: 70- 77.
[31] Klimeš L, Doležal J. An experimental assessment of the upper elevational limit of flowering plants in the western Himalayas. Ecography, 2010, 33(3): 590- 596.
[32] 罗天祥, 赵士洞. 中国杉木林生物生产力格局及其数学模型. 植物生态学报, 1997, 21(5): 403- 415.
[33] Odum E P. Fundamentals of Ecology. Philadelphia, PA: Saunders, 1971.
[34] 李娜, 王根绪, 杨燕, 高永恒, 柳林安, 刘光生. 短期增温对青藏高原高寒草甸植物群落结构和生物量的影响. 生态学报, 2011, 31(4): 895- 905.
[35] Webb W L, Lauenroth W K, Szarek S R, Kinerson R S. Primary production and abiotic controls in forests, grasslands, and desert ecosystems in the United States. Ecology, 1983, 64(1): 134- 151.
[36] Epstein H E, Lauenroth W K, Burke I C. Effects of temperature and soil texture on ANPP in the U.S. great plains. Ecology, 1997, 78(8): 2628- 2631.
[37] Vitousek P M, Howarth R W. Nitrogen limitation on land and in the sea: How can it occur?. Biogeochemistry, 1991, 13(2): 87- 115.
[38] Luo T X, Zhang L, Zhu H Z, Daly C, Li M C, Luo J. Correlations between net primary productivity and foliar carbon isotope ratio across a Tibetan ecosystem transect. Ecography, 2009, 32(3): 526- 538.
[39] Wieser G, Tausz M. Trees at their upper limit: treelife limitation at the alpine timberline. Netherlands: Springer, 2007.
[40] Bai Y F, Wu J G, Xing Q, Pan Q M, Huang J H, Yang D L, Han X G. Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau. Ecology, 2008, 89(8): 2140- 2153.
[41] Tilman D, Wedin D, Knops J. Productivity and sustainability influenced by biodiversity in grassland ecosystems. Nature, 1996, 379(6567): 718- 720.
[42] Naeem S, Li S B. Biodiversity enhances ecosystem reliability. Nature, 1997, 390(6659): 507- 509.
[43] Naeem S, Thompson L J, Lawler S P, Lawton J H, Woodfin R M. Declining biodiversity can alter the performance of ecosystems. Nature, 1994, 368(6473): 734- 737.
[44] Chen B X, Zhang X Z, Tao J, Wu J S, Wang J S, Shi P L, Zhang Y J, Yu C Q. The impact of climate change and anthropogenic activities on alpine grassland over the Qinghai-Tibet Plateau. Agricultural and Forest Meteorology, 2014, 189- 190: 11- 18.
[45] 李克让, 曹明奎, 於琍, 吴绍洪. 中国自然生态系统对气候变化的脆弱性评估. 地理研究, 2005, 24(5): 653- 663.
[46] 郑度, 林振耀, 张雪芹. 青藏高原与全球环境变化研究进展. 地学前缘, 2002, 9(1): 95- 102.
AltitudinalvariationsinthesensitivityofalpinemeadowproductivitytotemperatureandprecipitationchangesalongthesouthernslopeofNyainqentanglhaMountains
LI Xiang1,2, WANG Zhong3, ZHAO Jingxue1,2, LUO Tianxiang1,*
1KeyLaboratoryofAlpineEcologyandBiodiversity,InstituteofTibetanPlateauResearch,ChineseAcademyofSciences,Beijing100101,China2UniversityofChineseAcademyofSciences,Beijing100049,China3CollegeofLifeScience,WuhanUniversity,Wuhan430072,China
中国科学院先导专项B“青藏高原多圈层相互作用及资源环境效应”子课题(XDB03030402);国家自然科学基金面上项目(31170451);国家973计划课题“全球变化影响下自然生态系统的脆弱性及评价指标”(2010CB951301)
2016- 06- 14; < class="emphasis_bold">网络出版日期
日期:2017- 04- 24
10.5846/stxb201606141147
*通讯作者Corresponding author.E-mail: luotx@itpcas.ac.cn
李翔,王忠,赵景学,罗天祥.念青唐古拉山南坡高寒草甸生产力对温度和降水变化的敏感性及其海拔分异.生态学报,2017,37(17):5591- 5601.
Li X, Wang Z, Zhao J X, Luo T X.Altitudinal variations in the sensitivity of alpine meadow productivity to temperature and precipitation changes along the southern slope of Nyainqentanglha Mountains.Acta Ecologica Sinica,2017,37(17):5591- 5601.
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18
青海草业(2022年2期)2022-07-23
散文诗(2021年22期)2022-01-12
环球时报(2019-05-23)2019-05-23
活力(2019年21期)2019-04-01
少儿美术(快乐历史地理)(2018年7期)2018-11-16
东北林业大学学报(2018年7期)2018-07-20
河北林业科技(2016年5期)2016-11-08
生态学报(2016年12期)2016-08-09
中国水土保持科学(2016年3期)2016-07-19
