
马尾松谷胱甘肽过氧化物酶PmGPX基因的克隆及表达分析
2017-10-10 02:39罗群风吴东山杨章旗
浙江农林大学学报 2017年5期
陈 虎,贾 婕,罗群风,吴东山,杨章旗
(广西壮族自治区林业科学研究院 国家林业局马尾松工程技术研究中心/广西马尾松工程技术研究中心,广西 南宁 530002)
马尾松谷胱甘肽过氧化物酶PmGPX基因的克隆及表达分析
陈 虎,贾 婕,罗群风,吴东山,杨章旗
(广西壮族自治区林业科学研究院 国家林业局马尾松工程技术研究中心/广西马尾松工程技术研究中心,广西 南宁 530002)
克隆获得马尾松Pinus massoniana PmGPX基因,进行生物信息学分析以及在不同抗虫材料表达模式分析。在前期马尾松转录组测序获得该基因片段基础上,用cDNA末端快速扩增(RACE)技术获得基因全长,实时荧光定量聚合酶链式反应(qRT-PCR)技术分析该基因在不同抗虫材料中的表达模式。克隆获得基因cDNA全长序列,命名为PmGPX。该基因包含741 bp开放阅读框,编码246个氨基酸。序列分析表明:该基因具有GPX家族典型的保守结构域,等电点PI 8.29,蛋白分子量27.08 kDa,与针叶植物北美云杉Picea sitchensis的同源性较高,为91%。与单子叶和双子叶植物同源性较差,大部分在64%~80%,而在GPX家族典型保守结构域的同源性较高,为82%~100%。qRT-PCR技术分析表明:PmGPX在所有组织中均有表达,在针叶中表达量最高,嫩叶的表达量是根的33.45倍,在根、花和球果中的表达量较低,不同材料日表达模式相似,总体上随时间出现 “升—降—升”模式。该基因在抗虫材料中启动较早,且表达量高于对照。推测该基因参与了马尾松抗虫防御体系。图5参29
林木育种学;马尾松;谷胱甘肽过氧化物酶;PmGPX基因;克隆与表达
Abstract:The study PmGPX genes was played a role in Pinus massoniana insect resistance,that the great significance for in-depth insect-resistant resistance mechanism in Pinus massoniana.PmGPX,a cDNA region encoding glutathione peroxidase,was cloned in the transcriptome sequencing analysis of one insect-resistant material in Pinus massoniana.Analyses included an amino acid sequence and an expression (qRT-PCR)analysis.Results showed that the total length of open reading frame (ORF) was 741 bp encoding 246 amino acids.The amino acid sequence analysis showed three high conservative character fields of the GPX gene family for this gene.The isoelectric point of PmGPX was PI 8.29,and the protein molecular weight was 27.08 kDa.In genetic clustering,compared with some other plants,PmGPX showed a higher homology with Picea sitchensis (91%)but a low homology with other monocots and dicotyledonous plants (64%-80%).However,homologies of the three typical conservative GPX family domains(G1,G2,G3)reached a higher level with other plants (82%-100%).The expression analysis indicated that PmGPX was expressed in all kinds of tissues with higher expression levels in leaves (P<0.01),especially in young leaves (33.45 times more than the expression level in roots).Daily expression patterns were same in different insect-resistant materials and showed up-down-up-regu-lated trend the day.In insect-resistant material,expression of PmGPX reached a higher point earlier than that of control (P<0.05 in old leaves and tender stem).The expression analysis also indicated that PmGPX was involved in the insect-resistant defense system of P.massoniana.This study laid a foundation for further analysis of the PmGPX biological function and its role in insect-resistant defense with Pinus massoniana.[Ch,5 fig.29 ref.]
Key words:forest tree breeding;Pinus massoniana;glutathione peroxidase;PmGPX;cloning and expression analysis
谷胱甘肽过氧化物酶(glutathione peroxidase,GPXs)是一种重要的过氧化物分解酶,能清除活性氧,阻止脂膜氧化,起到保护细胞免受氧化胁迫的作用,具有重要的生理功能[1]。自CRIQUI等[2]从烟草Nicotiana tabacum中获得GPX基因后, 先后从柑橘Citrus[3], 番茄Lycopersicon esculentum[4], 水稻Oryza sativa[5], 香蕉 Musa nana[6], 荷花 Nelumbo nucifera[7], 枣 Ziziphus jujube[8]等植物中获得 GPX 基因。 近年来,对于植物GPXs的研究主要集中在生物胁迫和非生物胁迫(如高温、低温、干旱、重金属毒害、耐盐等)方面,尤其是GPXs在非生物胁迫中的功能作用,证实了GPX基因与植物抗逆性紧密相关。在非生物胁迫下,GPX基因在转录水平发生变化,参与相关氧化还原途径[9]。如苹果Malus MdGSTU1提高苹果的耐盐能力[10],过量表达NtGPX基因提高烟草的抗冻性[11],番茄LePHGPX基因的过量表达提高植株耐高温能力[12]。研究人员对拟南芥Arabidopsis thaliana GPX基因的功能研究取得较大进展,新的GPX被不断发掘(拟南芥目前已知8个),对其中7个GPX基因的研究证实,全部对非生物胁迫有应答反应,转基因可提高植物抵御胁迫的能力[13],如AtGPX3能显著增强植株的抗旱性[14],AtGPX1具有抗高温下的氧化胁迫能力[15]。目前虽有多种植物GPX基因被发现,并开展相关功能研究,但主要集中于单子叶和双子叶植物,对裸子植物的研究较少。马尾松Pinus massoniana属裸子植物松科Pinaceae松属Pinus植物,是中国南方重要的用材林树种[16],但在马尾松生长过程中常受到病虫害的危害,给林业经济带来严重损失,制约了马尾松人工林的健康发展。本研究在高通量测序分析基础上结合cDNA末端快速扩增(RACE)技术,获得谷胱甘肽过氧化物酶基因,对其进行生物学分析,利用实时定量聚合酶链式反应(qRT-PCR)技术研究其不同组织、不同抗性材料、不同时间表达模式,为探求马尾松虫害防御机制提供依据。
1 材料与方法
1.1 实验材料
植物材料包括抗虫马尾松品种 ‘松韵’ ‘Songyun’和对照GC101无性系,材料选自广西马尾松桐棉种源,采自广西马尾松种质资源库。该库栽植于2007年,树龄8年生,栽植株行距为4 m×4 m,根、茎、叶、花、果采自普通马尾松无性系,雌雄球花样品采自2015年2月,其他组织采自2015年4月。日变化叶和茎的材料采自2015年5月,采样时间分别在当日的7:00,10:00,13:00,16:00,19:00,所有样品采摘后放入液氮中保鲜,带回实验室存放于-80℃冰箱中备用。
1.2 实验方法
1.2.1 RNA提取及PmGPX克隆 实验所有样品的RNA采用多酚多糖植物RNA提取试剂盒(天根公司)进行提取,具体步骤按说明书进行。提取完成后进行凝胶电泳检查质量,参照M-MLV逆转录酶合成cDNA,进行凝胶检查和浓度测定(采用核酸定量仪)后备用。根据前期马尾松转录组测序结果中PmGPX的部分 cDNA序列进行比对,根据获得的序列在 2端设计特异引物,3′端引物(5′-ATGGCATCCTCTTTCGCAGCATCT-3′), 5′端引物(5′-TGCAGCAAGCAGATTCTGTATATCC-3′)。 以不同组织等量混合cDNA为模板进行扩增。设定退火温度为56.0℃,具体聚合酶链式反应(PCR)扩增体系和参数参照Taq DNA聚合酶说明书进行操作,扩增产物经凝胶电泳检查获得目的条带后,用胶回收试剂盒进行回收纯化,连接、转化后进行菌液PCR检查正确后测序。
1.2.2 序列分析 采用在线SOSUI和Plant-mPLoc亚细胞定位,利用Proteomics Server,SignalP 4.1 Server,SOPMA,TargetP 1.1 Server,DAS软件分析基因等电点、信号肽、二级结构、亲水性和跨膜结构,利用Motif Scan和SMART分析基因的功能结构域。将PmGPX基因完整氨基酸序列在美国生物技术信息中心(NCBI)进行覆盖率和同源性比较,利用DNAMAN软件构建PmGPX与其他植物的进化树。
1.2.3 基因表达模式分析 根据马尾松内参基因筛选结果[17],利用UBI4基因和CYP基因作为内参基因,内参引物分别为UBI4F:AGCTCCGACACCATTGATAA,UBI4R:CCAAAGTACGTCCAT CTTCCA,CYPR:CAAGGGTTCGTCGTTCCAC,CYPF:GGCAAACTTCTCGCCGTA,同时设计PmGPX基因引物,PmGPX1F:TCAAACTCCCCGTCTTATGG,PmGPX1R:GCTCCAACTGAATCCT TGC。 参照 SYBR Premix Ex TaqⅡ(Prefect real-time)试剂盒,构建qRT-PCR体系和扩增程序,其中退火为58℃,采用45个循环作熔解曲线。设生物学重复 3 个·样品-1, 根据 2-ΔΔCt法[18]计算相对表达量。
2 结果与分析
2.1 基因全长获得
对马尾松不同抗虫材料高通量测序比对发现,有个基因在抗虫材料中大量表达(log2.Fold_1.1491),该基因在KEGG代谢途径中只参与谷胱甘肽代谢(glutathione metabolism)和花生四烯酸代谢(arachidonic acid metabolism)途径。这2个途径中只有该基因上调表达,通过对序列在NCBI进一步比对,确定为谷胱甘肽过氧化物酶(glutathione peroxidase)基因。通过进行扩增、比对,获得了该基因完整开放阅读框(ORF)序列,命名为PmGPX。该基因完整开放阅读框(ORF)共包括741 bp,编码246个氨基酸(图1)。

图1 PmGPX基因cDNA序列及氨基酸序列Figure 1 cDNA and amino acid sequence of PmGPX
2.2 PmGPX基因序列分析
利用生物信息学软件对PmGPX编码的氨基酸序列进行分析,结果显示:该基因不含信号肽,有6个丝氨酸(Ser)和1个色氨酸(Try)磷酸化位点,2个糖基化位点;二级结构预测中:α-螺旋占30.49%,随机卷曲占38.62%,延伸链占20.33%,β-转角占10.57%;该基因等电点为8.29,蛋白分子量为27.08,在113~117个氨基酸处存在1个跨膜结构,亚细胞定位于叶绿体。
对PmGPX蛋白进行功能结构域显示,具有3个N-糖基化位点,4个N-酰基化位点,2个酪蛋白激酶Ⅱ磷酸化位点,4个蛋白激酶c磷酸化位点,二硫键氧化还原酶结构域、硫氧还蛋白结构域、固氮铁蛋白、抗氧化蛋白AhpC/TSA家族结构域、谷胱甘肽过氧化物酶结构域,以及谷胱甘肽氧化酵素活性部位和信号位点(图1)。
2.3 PmGPX蛋白同源性及进化
PmGPX核酸序列进行比对发现,序列同源性在62%~100%,相似度在74%~94%,与白云杉Picea glauca和北美云杉Picea sitchensis相似度最近,与葡萄Vitis vinifera,烟草,龙眼Dimocarpus longan等植物相似度为74%~78%。氨基酸序列比对发现,序列同源性在64%~100%,除与北美云杉相似度在91%外,与其他相似度为58%~81%,大部分为64%~80% ,与山嵛菜Eutrema yunnanense,亚麻芥Camelina sative,菜豆Phaseolus vulgaris的相似度低,总体上除与针叶类同源性较高外,其他都较低。但对谷胱甘肽过氧化物酶保守域同源性比对时发现,PmGPX氨基酸序列与其他植物具有很高的同源性,特别是在3个特征性结构域G1,G2,G3高度保守,PmGPX构成GPX催化三联体的3个氨基酸(Cys119,Gln150,Trp209), 具有植物 GPX 蛋白活性中心的3个保守半胱氨酸(Cys)残基 Cys119,Cys148,Cys167(图2)。
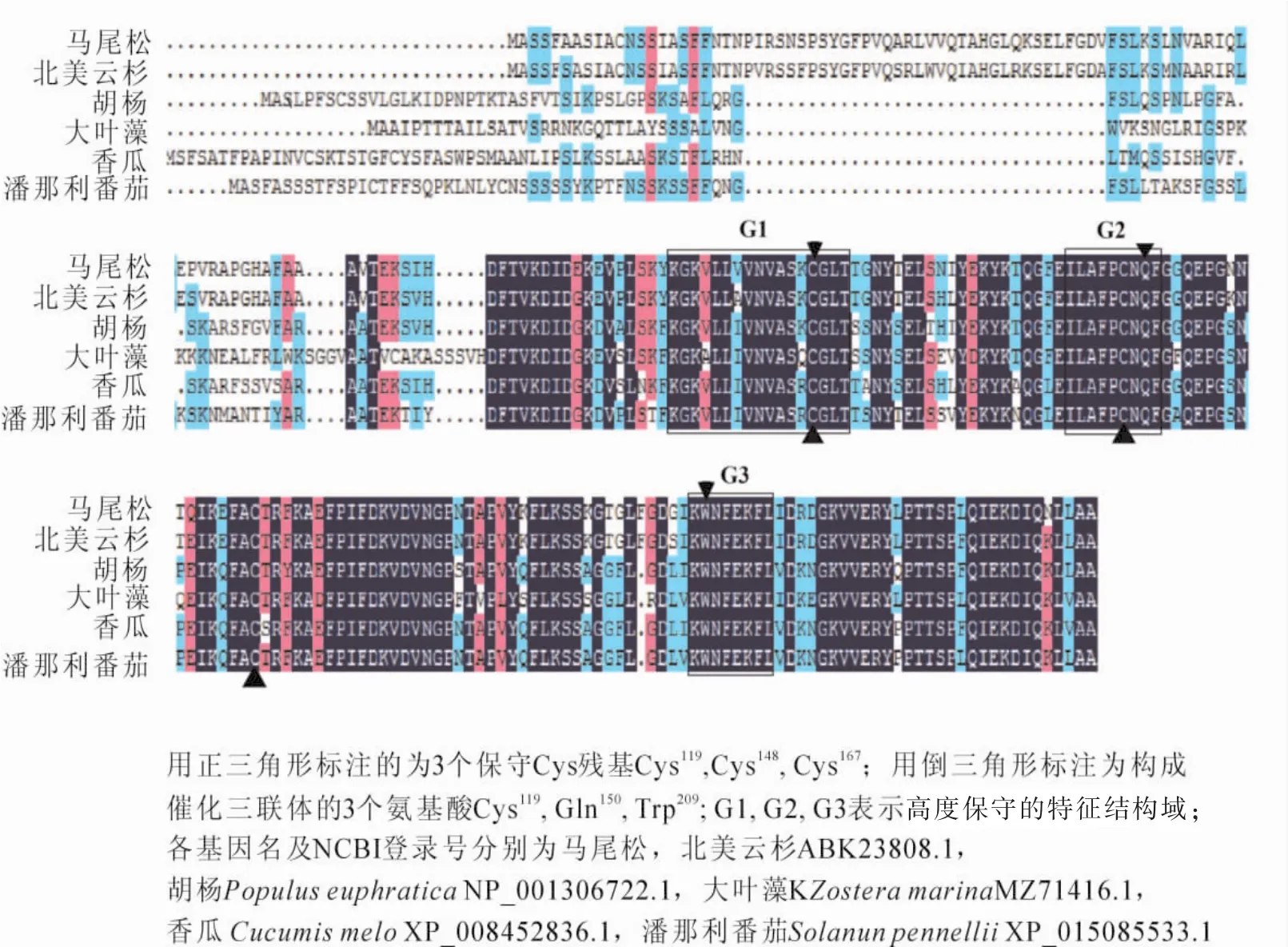
图2 PmGPX氨基酸序列比对Figure 2 Multiple sequence alignment of amino acid of the PmGPX proteins isolated
采用邻接法(neighbor-joining method)对20个植物的GPX基因构建进化关系树,马尾松与北美云杉聚为一类,并且与大叶藻的同源关系较近,与山嵛菜,亚麻芥,菜豆,无油樟Amborella trichopoda,小米Setaria italica和玉米Zea mays的同源关系较远。推测马尾松PmGPX基因与针叶植物,尤其是与云杉的该基因在进化过程是相似的,而与其他植物则存在较大差异,单子叶植物玉米、香瓜Cucumis melo同源关系较远,双子叶植物间相对的分化较小(图3)。
2.4 PmGPX基因表达分析
实时荧光定量对根、茎、叶、花、果中PmGPX基因的表达特征进行分析表明,PmGPX基因在所有组织中都有表达,但表达量存在明显的差异。PmGPX基因主要表达于针叶和茎中,在针叶中的表达量要高于其他组织,在嫩叶中的表达量为根中表达量的33.45倍,针叶成熟度的不同表达量差异也明显,嫩叶中表达量是成熟叶的2.72倍。PmGPX基因在不同成熟茎中的表达量差异不明显,虽仅次于在叶中的表达量,但相差11.04倍,在根、雌雄花、果等4个组织中的表达量相差不大,表达量均较少(图4)。
为进一步研究PmGPX基因在不同材料1 d内的变化模式,采用qRT-PCR进行分析,结果如图5所示。在嫩叶和老叶中,2个材料PmGPX基因表达变化趋势相似,都是 “升—降—升”模式。在嫩叶中抗虫材料在10:00时表达量达到最大值,是7:00表达量的2.65倍,而对照中该基因从7:00-16:00的表达量变化不大,只有到19:00时才达到峰值。老叶中2种材料的表达规律一致,不同的是PmGPX在抗虫材料中的表达要高于对照,在13:00时表达量要高于对照。从茎中的表达模式分析,PmGPX在不同成熟茎和不同抗性材料中的变化趋势一致,随时间推移,表达量不断上升。不同的是在嫩茎中前4个时间点,2种材料的表达量基本相同,在19:00时抗虫材料的表达量才出现明显的上升。而在老茎抗虫材料中PmGPX的表达量始终高于对照,在13:00时两者的表达量相差3.48倍。
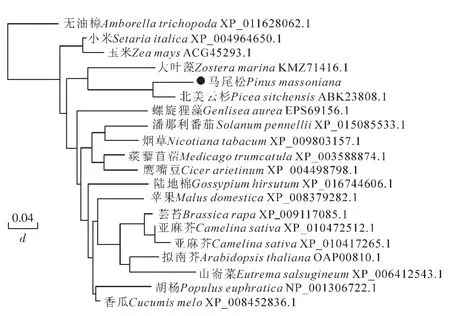
图3 PmGPX与其他GPX氨基酸序列的聚类分析Figure 3 Amino acid sequences cladogram analysis between PmGPX and other GPX
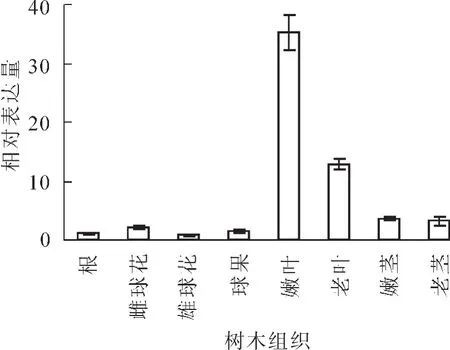
图4 马尾松PmGPX基因在不同组织的表达Figure 4 Expression of PmGPX gene in different organs by fluorescent quantitative RT-PCR
3 讨论
在马尾松抗虫材料转录组测序差异分析发现,在谷胱甘肽代谢存在唯一大量上调表达的基因,并且参与了花生四烯酸代谢途径,通过克隆比对确定为谷胱甘肽过氧化物酶基因,命名为PmGPX。该基因与针叶植物云杉同源性较高,在谷胱甘肽过氧化物酶保守域与其他植物同源性也较高,说明植物GPX基因在进化过程中虽然N端序列发现了改变,但在GPX基因功能结构域中的变化不大。另外,在PmGPX序列具有GPX中3个保守的特征性结构域,及GPX蛋白活性中心的3个保守半胱氨酸(Cys)残基。目前已知的植物GPX的活性位点由Cys作为催化残基来代替动物体内的硒代半胱氨酸[6,19],马尾松PmGPX也在Cys119处替代了动物PHGPXs中的硒代半胱氨酸残基,进一步证明PmGPX是GPX的家族成员。
PmGPX存在二硫键氧化还原酶结构域、硫氧还蛋白结构域、谷胱甘肽过氧化物酶结构域等功能结构域,在植物体内GPXs是含有巯基的过氧化物酶类,通过催化GSH来减少过氧化氢或有毒化合物,起到解毒和保护细胞免受破坏[20-21]。
MILLA等[13]推测拟南芥中的GPX基因分布于3条染色体上,定位在叶绿体、线粒体、胞质等亚细胞器中,如AtGPX2定位于细胞质。本研究PmGPX预测定位于叶绿体中,推测AtGPX1与抵御光氧化胁迫有关。活性氧在植物细胞代谢中主要来源于叶绿体、线粒体,亚细胞定位与功能有直接关系,马尾松PmGPX发挥的作用可能与光合或产生活性氧有关。
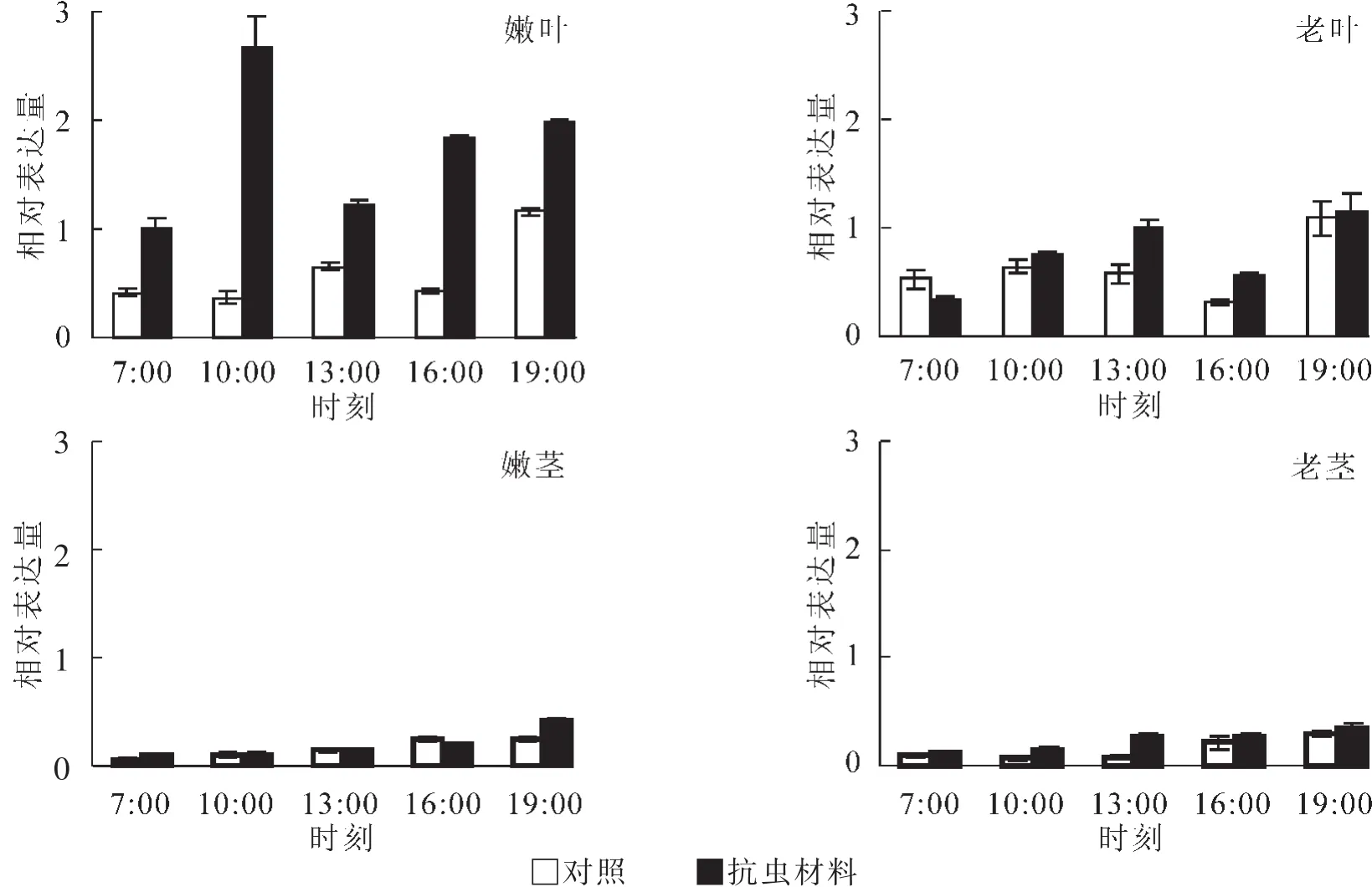
图5 荧光定量PCR检测PmGPX基因不同抗虫材料的表达Figure 5 Expression of PmGPX gene in resistant and susceptible varieties of Pinus massoniana by qRT-PCR
本研究中马尾松PmGPX在所有组织中均有表达,与已有研究结果一致,说明PmGPX在马尾松生长过程中起到清除细胞内活性氧(ROS)的作用[13],但PmGPX基因在叶和茎中的表达最高,这与拟南芥[13]和油菜Brassica napus[22]的研究结果一致,而香蕉MaGPX在花和根中表达量较高[6],白菜 Brassica pekinensis BrGPX6a在早期子叶胚中的表达量较高,在成熟胚中表达较低[23]。不同植物或同一植物GPX在不同组织中的表达均有所差别,可能与不同组织中活性氧的分布有关联[7,13,24]。由于PmGPX基因在叶中,尤其是嫩叶中的表达量最高,而在雌雄花和根中表达量极低,进一步验证了该基因亚细胞定位于叶绿体的可能。
PmGPX基因在不同抗性材料同一组织中的表达模式基本一致,主要区别在于抗性品种PmGPX基因表达量始终高于对照,并且随时间延长,表达量提升速度和达到峰值的时间均比对照要早。从选出的抗逆性材料来看,耐铝小麦Triticum sestivum和水稻品种,在受到铝胁迫后GPX活性明显提高,而对照则出现下降趋势[25-26],同样耐盐品种在盐胁迫下,本身含有或能够迅速提高植株的抗氧化能力,对照则较低[27]。从非生物胁迫下,香蕉MaGPX在盐、干旱等胁迫下表达量上升[7],拟南芥AtGPX1表达量的上升,会提高抵御光氧化胁迫[15]。
目前,有关GPX基因在植物抗虫功能的研究还较少,马尾松PmGPX基因在抗虫材料中的高表达,现有研究表明:植物中GSH合成和酶活性提高,能够增加植物对多种胁迫的抵抗能力,相反在环境胁迫下GPXs的mRNA水平也会提高,在谷胱甘肽代谢中GPX基因表达量提高,能够提高氧化谷胱甘肽量,从而转化成GSH, 提高植物抵御环境胁迫的能力[28-29]。马尾松PmGPX在抗虫材料中高表达,提高了防御虫害的能力,另外,PmGPX基因在抗虫材料花生四烯酸代谢中高量表达,推测该基因可能提高该途径15-oxoETE和5-oxoETE代谢物含量,从而提高丙酮含量,提高了防御抗虫的能力,但通过何种信号传导途径参与来起作用[14],有利于抵御虫害次生代谢物质的产生,起到抗虫效果还有待进一步研究。
[1] FLOHEÉ L,GÜNZLER W A.Assays of glutathione peroxidase [J].Methods Enzymol,1984,105(1):114 - 121.
[2] CRIQUI M C,JAMET E,PARMENTIER Y,et al.Isolation and characterization of a plant cDNA showing homology toanimal glutathione peroxidases [J].Plant Mol Biol,1992,18(3):623 - 627.
[3] HOLLAND D,BEN-HAYYIM G,FALTIN Z,et al.Molecular characterization of salt-stress-associated protein in Citrus:protein and cDNA sequence homology to mammalian glutathione peroxidase [J].Plant Mol Biol,1993,21(5):923-927.
[4] DEPÈGE N,DREVET J,BOYER N.Molecular cloning and characterization of tomato cDNAs encoding glutathione peroxidase-like proteins [J].Eur J Biochem,1998,253(2):445 - 451.
[5] LI Wenjun,FENG Hai,FAN Jinghua,et al.Molecular cloning and expression of a phospholipid hydroperoxide glutathione peroxidase homolog in Oryza sativa [J].BBA Gene Struct Exp,2000,1493(1/2):225 - 230.
[6] 张丽丽,徐碧玉,刘菊华,等.香蕉谷胱甘肽过氧化物酶基因MaGPX的克隆和表达分析[J].园艺学报,2012, 39(8): 1471 - 1481.ZHANG Lili,XU Biyu,LIU Juhua,et al.Isolation and expression analysis of a cDNA encoding glutathione peroxidase from banana [J].Acta Hortic Sin,2012,39(8):1471 - 1481.
[7] DIAO Ying,XU Huaxue,LI Guolin,et al.Cloning a glutathione peroxidase gene from Nelumbo nucifera and enhanced salt tolerance by overexpressing in rice [J].Mol Biol Rep,2014,41(8):4919 - 4927.
[8] 肖蓉,罗慧珍,张小娟,等.干旱和盐胁迫条件下枣树谷胱甘肽过氧化物酶基因(ZjGPX)的差异表达及功能分析[J].中国农业科学, 2015, 48(14): 2806 - 2817.XIAO Rong,LUO Huizhen,ZHANG Xiaojuan,et al.Differential expression and functional analysis of glutathione peroxidase gene from jujube (ZjGPX) under drought and salt stresses [J].Sci Agric Sin,2015,48(14):2806 - 2817.
[9] AMTMANN A,BOHNERT H J,BRESSAN R A.Abiotic stress and plant genome evolution.Search for new models[J].Plant Physiol,2005,138(1):127 - 130.
[10] 安秀红,徐锴,厉恩茂,等.苹果抗性相关的谷胱甘肽转移酶基因MdGSTU1的生物信息学和表达分析[J].中国农业科学, 2014, 47(24): 4868-4877.AN Xiuhong,XU Kai,LI Enmao,et al.Bioinformatics and expression analysis of MdGSTU1 gene encoding a resistance-related glutathione transferase from apple [J].Sci Agric Sin,2014,47(24):4868 - 4877.
[11] ROXAS V P,LODHI S A,GARRETT D K,et al.Stress tolerance in transgenic tobacco seedlings that overexpress glutathione S-transferase/glutathione peroxidase [J].Plant Cell Physiol,2000,41(11):1229 - 1234.
[12] CHEN Shaorong,VAGHCHHIPAWALA Z,LI Wei,et al.Tomato phospholipid hydroperoxide glutathione peroxidase inhibits cell death induced by bax and oxidative stresses in yeast and plants [J].Plant Physiol,2004,135(3):1630-1641.
[13] MILLA M A R,MAURER A,HUETE A R,et al.Glutathione peroxidase genes in Arabidopsis are ubiquitous and regulated by abiotic stresses through diverse signaling pathways [J].Plant J,2003,36(5):602 - 615.
[14] MIAO Yuchen,LÜ Dong,WANG Pengcheng,et al.An Arabidopsis glutathione peroxidase functions as both a redox transducer and a scavenger in abscisic acid and drought stress responses [J].Plant Cell,2006,18(10):2749 - 2766.
[15]CHANG C C C,'SLESAK I,JORDÁ L,et al.Arabidopsis chloroplastic glutathione peroxidases play a role in cross talk between photooxidative stress and immune responses [J].Plant Physiol,2009,150(2):670 - 683.
[16] 杨章旗,舒文波.马尾松纸浆材育种研究进展及策略[J].广西林业科学,2012,41(1):27-32.YANG Zhangqi,SHU Wenbo.Breeding progress and strategies for pulpwood of Pinus massoniana [J].Guangxi For Sci,2012,41(1):27 - 32.
[17] CHEN Hu,YANG Zhangqi,HU Ying,et al.Reference genes selection for quantitative gene expression studies in Pinus massoniana L.[J].Trees,2016,30(3):685 - 696.
[18] LIVAK K J,SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmethod [J].Methods,2001,25(4):402 - 408.
[19] HERBETTE S,ROECKEL-DREVET P,DREVET J R.Seleno-independent glutathione peroxidases.more than simple antioxidant scavengers [J].FEBS J,2007,274(9):2163 - 2180.
[20] MAY M J,VERNOUX T,LEAVER C,et al.Glutathione homeostasis in palnt:implications for environmental sensing and plant development[J].J Exp Bot,1998,49(321):649 - 667.
[21] NOCTOR G,GOMEZ L,VANACKER H,et al.Interactions between biosynthesis,compartmentation and transport in the control of glutathione homeostasis and signalling [J].J Exp Bot,2002,53(372):1283 - 1304.
[22] 胡茂龙,浦惠明,龙卫华,等.甘蓝型油菜谷胱甘肽过氧化物酶基因的克隆及表达分析[J].江苏农业学报,2011, 27(5): 950 - 956.HU Maolong,PU Huiming,LONG Weihua,et al.Cloning and expression of glutathione peroxidase 2 gene in rapeseed (Brassica napus L.) [J].Jiangsu J Agric Sci,2011,27(5):950 - 956.
[23] 齐增园,陶鹏,李必元,等.白菜谷胱甘肽过氧化物酶基因GPX的鉴定与分析[J].浙江农业学报,2016,28(1): 64 - 69.QI Zengyuan,TAO Peng,LI Biyuan,et al.Identification and analysis of GPX genes in Chinese cabbage [J].Acta A-gric Zhejiang,2016,28(1):64 - 69.
[24] YANG Xiaodong,LI Wenjun,LIU Jinyuan.Isolation and characterization of a novel PHGPx gene in Raphanus sativus[J].BBA Gene Struct Expr,2005,1728(3):199 - 205.
[25] DARKÓ É,AMBRUS H,STEFANOVITS-BÁNYAI É,et al.Aluminium toxicity,Al tolerance and oxidative stress in an Al-sensitive wheat genotype and in Al-tolerant lines developed by in vitro microspore selection [J].Plant Sci,2004,166(3):583 - 591.
[26] MA Baohui,WAN Jianmin SHEN Zhenguo.H2O2production and antioxidant responses in seeds and early seedlings of two different rice varieties exposed to aluminum [J].Plant Growth Regul,2007,52(1):91 - 100.
[27] SHALATA A,MITTOVA V,VOLOKITA M,et al.Response of the cultivated tomato and its wild salt-tolerant relative Lycopersicon pennelli to salt-dependent oxidative stress:the root antioxidative system [J].Physiol Plant,2001,112(4):487 - 494.
[28] ESHDAT Y,HOLLAND D,FALTIN Z,et al.Plant glutathione peroxidases [J].Physiol Plant,1997,100(2):234 -240.
[29] 陈坤明,宫海军,王锁民.植物谷胱甘肽代谢与环境胁迫[J].西北植物学报,2004,24(6):1119-1130.CHEN Kunming,GONG Haijun,WANG Suomin.Glutathione metabolism and environmental stresses in plants [J].Acta Bot Boreal-Occident Sin,2004,24(6):1119 - 1130.
Cloning and expression analysis of the PmGPX gene encoding glutathione peroxidase in Pinus massoniana
CHEN Hu,JIA Jie,LUO Qunfeng,WU Dongshan,YANG Zhangqi
(Engineering Research Center of Masson Pine of State Forestry Administration/Engineering Research Center of Masson Pine of Guangxi,Guangxi Forestry Research Institute,Nanning 530002,Guangxi,China)
S791.2
A
2095-0756(2017)05-0856-08
2016-07-05;
2016-09-16
国家自然科学基金资助项目(31660219);广西 “八桂学者”专项经费资助项目(2011A015);广西自然科学基金资助项目(2014GXNSFBA118104)
陈虎,高级工程师,从事林木遗传育种研究。E-mail:chenhubeijing-2008@163.com。通信作者:杨章旗,教授级高级工程师,博士,从事林木遗传育种研究。E-mail:yangzhangqi@163.com
猜你喜欢
河北科技师范学院学报(2022年2期)2022-08-26
现代养生·上半月(2021年10期)2021-09-24
化工管理(2021年21期)2021-08-16
林业科技(2020年3期)2021-01-21
中国药剂学杂志(网络版)(2019年4期)2019-07-17
水产科学(2019年3期)2019-05-30
分析化学(2018年4期)2018-11-02
分析化学(2018年7期)2018-09-17
江苏农业科学(2016年10期)2017-02-05
民生周刊(2016年19期)2016-10-25
