
铝胁迫条件下镧对百喜草幼苗根系生长的影响
2017-09-26 01:59罗涵夫宋华伟刘天增张巨明
草业学报 2017年9期
罗涵夫,宋华伟,刘天增,张巨明*
(1.华南农业大学林学与风景园林学院,广东 广州 510642;2.广东省草业工程技术研究中心,广东 广州 510642)
铝胁迫条件下镧对百喜草幼苗根系生长的影响
罗涵夫1,2,宋华伟1,2,刘天增1,2,张巨明1,2*
(1.华南农业大学林学与风景园林学院,广东 广州 510642;2.广东省草业工程技术研究中心,广东 广州 510642)
镧是稀土元素中最重要和最活泼的元素,具有促进种子萌发、提高作物产量和增强抵御逆境胁迫等作用。研究以华南地区水土保持常用的百喜草幼苗为材料,通过水培实验探究在酸性铝胁迫条件下镧对百喜草幼苗根生长的影响。实验在0.9 μmol/L Al3+的酸性铝胁迫水培条件下,以添加0.3 μmol/L的La3+作为处理,在水培的第2,4,6天,分别测定根系形态、根干鲜比、根茎比、根系活力。结果表明,在酸性铝胁迫水溶液中添加La3+,可提高幼苗的主根长、根表面积、根尖数、根体积、根干鲜比、根茎比、根系活力,说明在酸性铝胁迫条件下,镧元素可以促进百喜草幼苗根系的生长。
百喜草;根系;铝胁迫;镧
自然条件下,土壤中含有大量铝、锰氢氧化物,这些形态的铝、锰氢氧化物通常是难溶性的,所以对植物没有毒害。而土壤的酸化可以加快土壤中铝、锰氢氧化物的溶解,尤其是在土壤酸性较为严重的情况下,可以产生大量植物可以吸收形态的铝、锰化合物,引起土壤铝和锰等元素活动性增强,浓度增加。吸收形态的铝主要是以Al3+和单核羟基铝的形式存在,其含量会随着土壤pH值的降低呈快速增加的趋势。并且当土壤中活性铝水平达到一定的阈值,植物就会出现多种铝毒害的症状。主要表现为植物根系的结构遭到破坏,从而阻碍了根系对养分、水分的吸收以及根系的伸长,最终抑制了植物的生长、发育和繁殖[1]。酸性土壤中的铝胁迫被认为是引起植物生长缓慢甚至减产、草原以及森林退化的重要原因之一,因此提高植物耐铝性是植物抗逆研究的重大课题之一。
稀土元素对植物膜脂过氧化作用的研究表明,稀土元素能增加细胞的抗氧化酶类活性,从而减轻膜脂的过氧化损伤,保证细胞膜的正常功能[2]。添加适当浓度的镧溶液可以显著提高盐胁迫下黑麦草(Loliumperenne)根系过氧化物酶(peroxidase,POD)、超氧化物歧化酶(superoxide dismutase,SOD)和过氧化氢酶(catalase,CAT)的活性,提高抗坏血酸(ascorbic acid,AsA)和谷胱甘肽(glutathione,GSH)的含量,减少超氧阴离子(O2-)、过氧化氢(H2O2)和丙二醛(malondialdehyde,MDA)的含量。表明适当浓度的镧溶液能够提高植物细胞内活性氧清除能力,从而缓解盐胁迫产生的活性氧对黑麦草根系和叶绿体细胞膜造成的过氧化伤害,从而维持了植物膜系统的稳定性,降低细胞电解质的渗透率,维护植物正常生长和发育[3]。
百喜草(Paspalumnotatum)对环境的要求不高,在弱酸性的土壤中可以正常生长、繁殖,其根系能在含铝较高的土壤中生长,可以有效地减轻土壤水土流失、环境污染等灾害的发生[4]。在我国华南酸雨地区栽培种植后,发现百喜草具有高效的缓解金属毒害,明显改善土壤环境以及提供美化绿化等功能,同时它还具有易种植、易管理、抗逆性强等特点[5-6]。所以百喜草在我国华南地区被广泛用于边坡绿化及水土保持工程。
在我国南方红壤地区,土壤pH值过低,植物受铝毒害严重。尽管稀土元素的应用非常广泛,但其对酸性土壤铝胁迫条件下草坪草的作用研究很少。草坪草幼苗对环境条件十分敏感,苗期是草坪草能否建植成功的关键阶段。研究稀土元素镧对铝胁迫下百喜草幼苗生长的影响,不但为稀土元素在草坪中的应用,也为百喜草在我国南方酸性土壤地区的推广应用提供一定的科学依据。
1 材料与方法
1.1试验材料
百喜草幼苗。于2015年3月13日,用0.5%的NaClO消毒百喜草种子(由北京克劳沃有限公司提供),用蒸馏水冲洗数次。置于40 ℃的烘箱中烘干4 h,然后把烘干后的种子均匀地撒入放有双层滤纸的9 cm培养皿中,湿润,放入人工气候箱中,黑暗处理24 h,然后光照培养6 d。
1.2试验设计
挑选根系长度为(2.0±0.1) cm的幼苗,用海绵固定在直径为1 cm的泡沫板上,平均分成2份,分别放入装有营养液(5 L)的培养盆中。营养液中包含了0.2 mmol/L Ca(NO3)2,0.2 mmol/L KNO3,0.08 mmol/L MgSO4,0.04 mmol/L KH2PO4,9 μmol/L B,0.06 μmol/L Cu,0.02 μmol/L Mo,1.84 μmol/L Mn,0.16 μmol/L Zn和10 μmol/L Fe(Fe-EDTA)。其中一盆营养液中含有0.9 μmol/L的 Al溶液,标记为处理Al;另一盆营养液含有0.9 μmol/L的Al溶液和0.3 μmol/L的La溶液,标记为处理Al+La。营养液2 d更换一次。将培养盆放入人工气候箱中培养,设定的参数为白天温度(30±1) ℃,夜晚(25±1) ℃,光周期为16 h/8 h(光/暗),相对湿度为(80±5)%。
在水培的第2,4,6天,分别取出一部分培养液中的百喜草幼苗,用蒸馏水洗净,擦干外部水分之后,分离百喜草幼苗的根与茎,测定根系活力、根系干鲜重、根茎比和根系形态学等相关指标。
1.3指标测定及方法
1.3.1根系形态学参数 在透明有机玻璃盘中装入适量的蒸馏水,把分离好的根系放入水中,用镊子将重叠的根系充分展开,用Epson V700彩色图像扫描仪(法国)进行根系扫描,以像素300 dpi的黑白TIF图像文件存档,然后用WinRHIZO根系扫描分析软件对根系的根长、根表面积、根体积和根尖数进行分析。
1.3.2干鲜重、根茎比和干鲜比 将幼苗用蒸馏水洗净,擦干外部水分之后,分离叶片和根系,分别称量各自鲜重。然后105 ℃杀青,80 ℃烘干至恒重,分别称量其干重。根茎比=地上部干重/地下部干重。干鲜比=叶片(根系)干重/叶片(根系)鲜重。
1.3.3根系活力 分别取2,4,6 d的百喜草根系样品100个,浸没在0.4%氯化三苯基四氮唑(TTC)溶液和6 mmol/L磷酸缓冲液的等量混合液中,在28 ℃下暗保温4 h,然后加入1 mol/L硫酸溶液2 mL,停止反应(与此同时做一空白试验,先加硫酸,然后加入百喜草根样品,37 ℃下暗保温4 h后加硫酸,其溶液浓度和操作步骤同上)。然后把根取出,充分吸干水分后加2 mL乙酸乙酯和石英砂研磨均匀。用滤纸过滤后将提取液移入10 mL刻度试管,用乙酸乙酯多次清洗残渣,然后都倒入试管中,加乙酸乙酯定容至刻度线。用METASH(上海) UV5200型分光光度计在波长485 nm下测定,记录吸光值[7]。计算公式:
TTC还原强度=TTC还原量(μg)/时间(h)
1.4数据处理
用Excel 2010统计数据并制图表,应用SPSS 17.0统计分析软件进行方差分析,多重比较以及两个独立样本的t检验,各平均值采用邓肯式新复极差检验法(Dumcan’s法),在P<0.05水平下进行多重比较。
2 结果与分析
2.1对百喜草根系形态的影响
2.1.1主根长 如图1所示,Al+La和Al处理的百喜草主根长均随着时间的延长而增加,但在第2,6天测定时段Al+La处理的百喜草主根长都显著高于Al处理,表明La元素可以使百喜草幼苗的主根伸长。
2.1.2根表面积 如图2所示,Al+La和Al处理的百喜草根表面积均随着时间的延长呈增加的趋势,在处理的第6天,Al+La处理的百喜草根表面积显著高于Al处理,表明La元素可促进百喜草幼苗根表面积的增长。

图1 百喜草幼苗主根长变化Fig.1 The change of axial root length of bahiagrass seedling
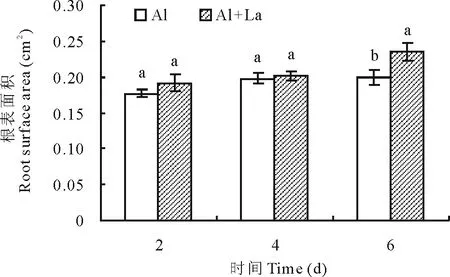
图2 百喜草幼苗根表面积变化Fig.2 The change of root surface area of bahiagrass seedling
Al:0.9 μmol/L Al,Al+La:0.9 μmol/L Al+0.3 μmol/L La.不同小写字母表示差异显著(P<0.05)。Different lowercase letters indicate significant differences atP<0.05 level.下同 The same below.
2.1.3根尖数 如图3所示,Al+La和Al处理的百喜草根尖数均随着时间的延长而呈增加的趋势,且第2,4,6天 Al+La处理的百喜草根尖数均高于Al处理,在第6天达到了显著水平,表明La元素有利于百喜草幼苗根尖数的增加。
2.1.4根体积 如图4所示,Al+La和Al处理的百喜草根体积均随着时间的延长而呈增加的趋势,且所有测定时段Al+La处理的百喜草根体积都高于Al处理,但是处理间的差异未达到显著水平。
2.2对百喜草干鲜重、干鲜比和根茎比的影响
2.2.1鲜重 随着测定时间的延长Al+La处理和Al处理的百喜草幼苗根系鲜重呈增加的趋势,且所有测定时段,Al+La处理的百喜草幼苗根系鲜重都显著高于Al处理(图5),分别高出了8.7%,20.9%,18.4%,表明在酸性铝胁迫条件下,La能促进根系生长,使根系鲜重增加。
2.2.2干重 随着测定时间的延长,Al+La处理和Al处理的百喜草幼苗根系干重均呈增加的趋势(图6),至处理的第6天,Al+La处理百喜草幼苗根系干重较第2天增重23.4%,而Al处理的百喜草幼苗根系干重仅增重3.6%,且所有测定时段,Al+La处理的百喜草幼苗根系干重都显著高于Al处理,表明在酸性铝胁迫条件下,La能促进根系干物质积累,使根系干重增加。

图3 百喜草幼苗根尖数的变化Fig.3 The change of root tips of bahiagrass seedling

图4 百喜草幼苗根体积变化Fig.4 The change of root volume of bahiagrass seedling
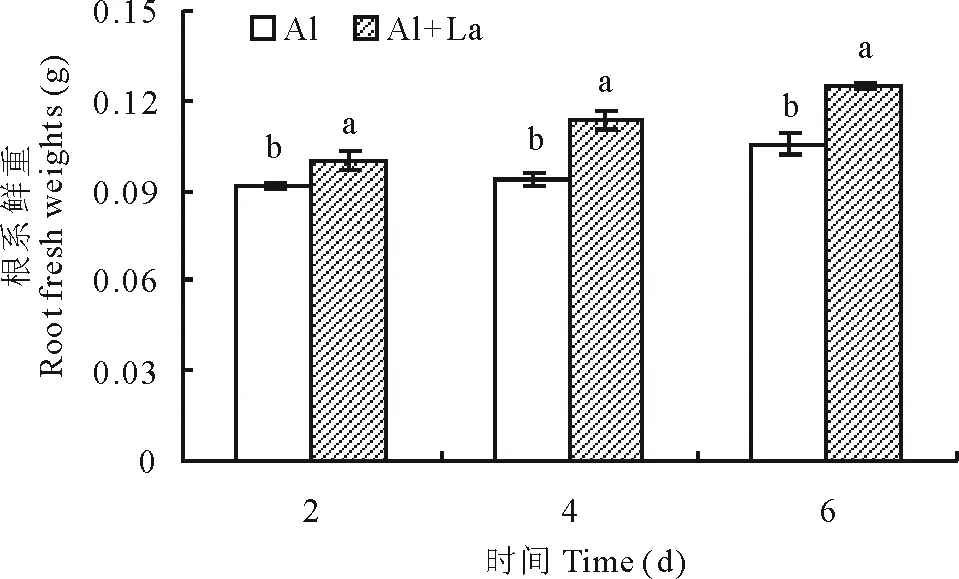
图5 百喜草幼苗根系鲜重变化Fig.5 The change of root fresh weight of bahiagrass seedling
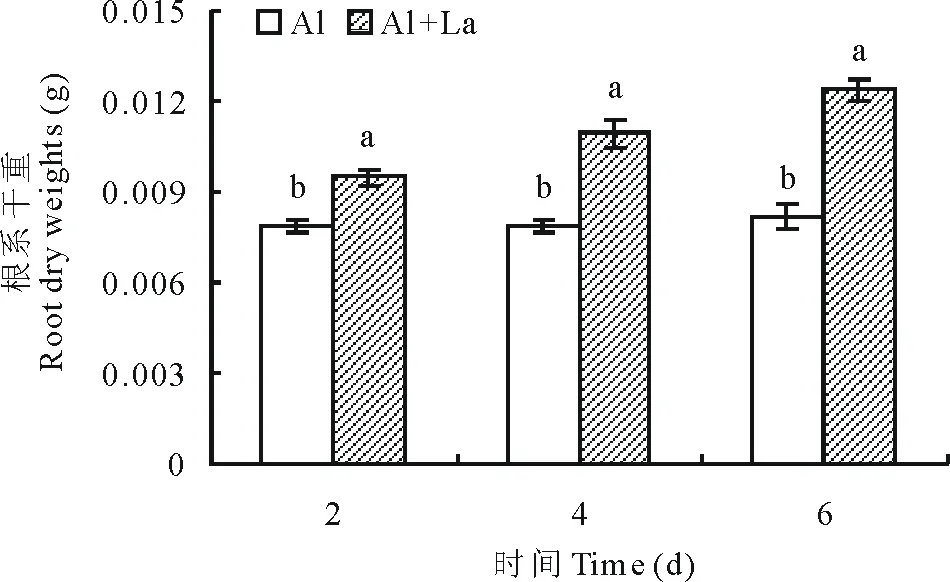
图6 百喜草幼苗根系干重变化Fig.6 The change of root dry weight of bahiagrass seedling
2.2.3干鲜比和根茎比 所有测定时段的Al+La处理的百喜草根系干鲜比都显著高于Al处理(表1),且Al处理的百喜草根系干鲜比整体呈下降的趋势,而Al+La处理的百喜草根系干鲜比整体呈增加趋势。百喜草根茎比方面,Al处理整体呈现先减少后增加的趋势,而Al+La处理整体呈增加趋势,且第4,6天测定中,Al+La处理的百喜草根茎比显著高于Al处理。
2.3对百喜草根系活力的影响
如图7所示,随着测定时间的延长Al+La处理和Al处理的百喜草幼苗根系活力均呈增加的趋势,且在处理的第2,4天,Al+La处理的百喜草幼苗根系活力都显著高于Al处理,但值得注意的是,在处理的第6天, 处理间的差异不显著, 这表明在处理的前期,较高的根系活力。
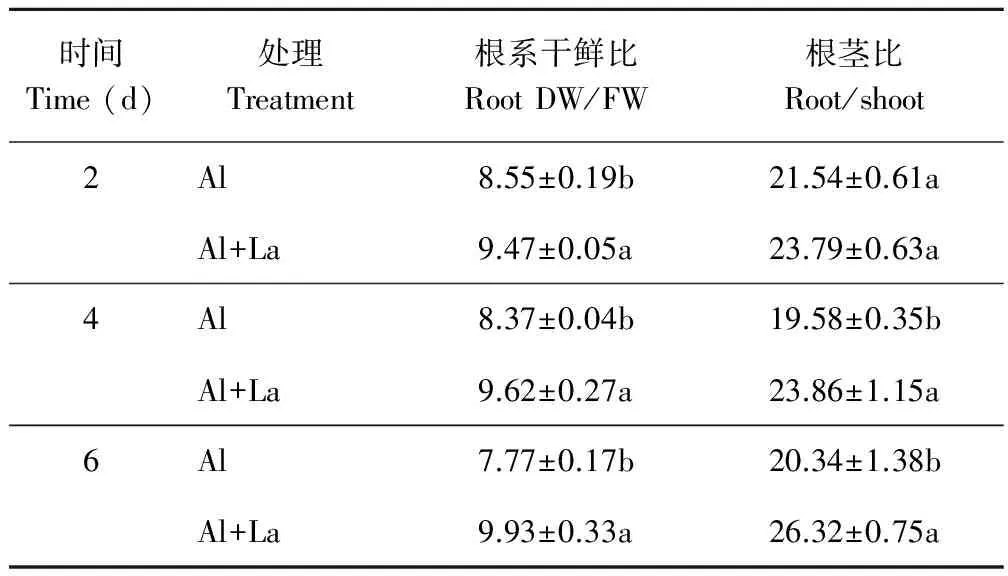
表1 百喜草根系干鲜比和根茎比变化Table 1 The change of root DW/FW ratio and root/shoot ratio of bahiagrass seedling %
注:同列不同小写字母表示差异显著(P<0.05)。
Note:Different lowercase letters within the same column indicate significant differences atP<0.05 level.生长在含有La3+的酸性铝溶液中的百喜草幼苗能维持
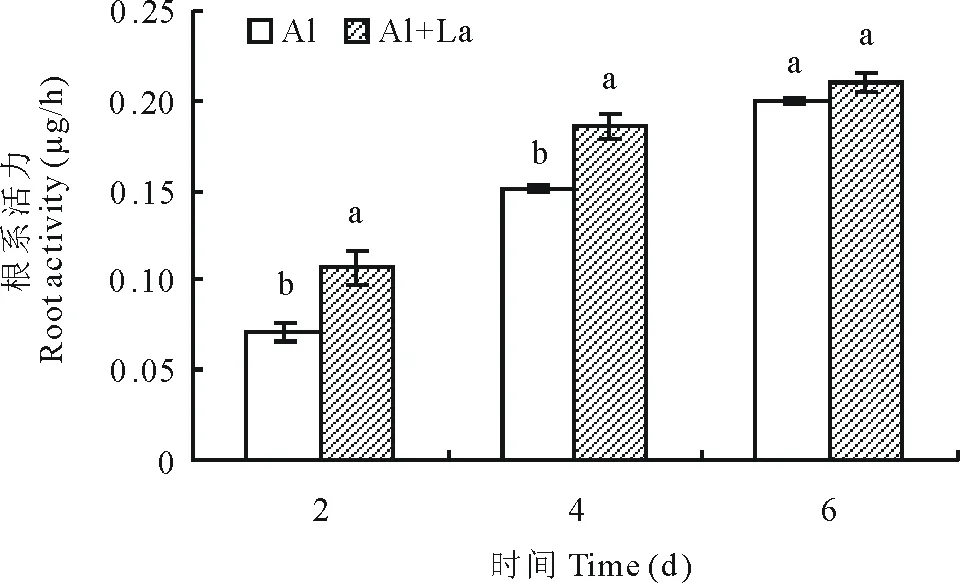
图7 百喜草幼苗根系活力的变化Fig.7 The change of root activity of bahiagrass seedling
3 讨论与结论
在逆境胁迫下,植物外部形态的变化是评价植物对逆境抗性强弱最直接和最可靠的指标之一[8]。通常在铝胁迫处理的一小段时间内,植物的根系生长便会受到铝毒抑制,这可能是由于铝毒抑制了植物根系细胞的伸长[9]。本研究发现,所有测定时段,Al+La处理百喜草幼苗的主根长、根尖数、根表面积和根体积都高于Al处理。且在第6天的测定中,达到了显著水平。说明铝能够抑制百喜草根尖细胞伸长和细胞分裂,从而引起根系变短、变细。而Al+La处理促进了百喜草幼苗根系的伸长,使根系的接触面积增加,有利于百喜草根系对养分和水分的吸收利用。这可能是由于适宜浓度的La3+可以提高百喜草幼苗根细胞质膜上ATP的活性,从而促进了百喜草幼苗的能量代谢和根系的吸收能力[10]。
生物量是反映百喜草幼苗生长最直观、最重要的指标之一。本研究结果表明,所有测定时段内,Al+La处理百喜草幼苗的根系鲜重、根系干重都显著高于Al处理。许多研究已经证实,稀土元素La可以通过增强植物体内各种代谢酶活性、加强呼吸强度,促进化合物转化和生理代谢,从而促进植物生长发育,达到增产的作用[11-12]。所有测定时段内,Al+La处理的百喜草根系干鲜比显著高于Al处理,且Al处理干鲜比整体呈下降的趋势,而Al+La处理的干鲜比整体呈增加趋势。第4,6天测定中,Al+La处理的根茎比显著高于Al处理。这可能是因为适宜浓度的La3+处理后百喜草幼苗细胞加氧酶(Rubisco)羧化活力增强,而Rubisco又是光合作用中决定碳同化速率的关键酶,Rubisco活性的增加促进了幼苗的光合作用,导致植株干物质增加明显[13]。
根系活力通常包括了植物根系的吸收、物质交换、氧化还原等能力。植物根系活力的强弱直接反映了根系代谢能力的强弱,从而影响植株的生长发育及抗胁迫能力,是反映根系生命活动的一项重要指标[14]。TTC法通过测定与植物呼吸作用紧密的相关琥珀酸脱氢酶活性来表示植物的根系活力[15]。本试验研究发现,所有测定时段Al+La处理百喜草幼苗的根系活力都高于Al处理,且在第2,4天达到了显著水平,这与王玉凤等[16]在钙对NaCl胁迫下玉米(Zeamays)幼苗根系活力的结果基本一致,表明百喜草根系活力的升高可能是对逆境进行保护的一种反应。Al胁迫下百喜草幼苗的根系受到了伤害,自身保护能力降低。La3+溶液在一定程度上可以缓解Al胁迫对百喜草幼苗的伤害,可能是由于适当浓度的La3+处理能提高百喜草幼苗抗氧化系统对活性氧的清除能力,从而缓解Al胁迫对根系的过氧化伤害[3]。在胁迫处理的第6天时,Al处理的百喜草幼苗根系活力略低于Al+La,这可能是由于百喜草幼苗的根系在Al胁迫下,通过调节自身的生理反应,从而长出了新的根尖,对铝胁迫产生了一定的耐性。
通过研究在最适La3+浓度对铝胁迫下百喜草生长的影响,得出以下结论:Al+La处理后,百喜草幼苗的根长、根系表面积、根体积和根尖数、根系干鲜重和干鲜比均显著高于Al处理,Al+La处理的根茎比显著高于Al处理,说明适当浓度的镧元素处理可以缓解铝毒害对百喜草幼苗造成的伤害。
References:
[1] Hunter D, Ross D S. Evidence for a phytotoxic hydroxy-aluminum polymer in organic soil horizons. Science, 1991, 251(4997): 1056-1058.
[2] Zeng F, An Y, Zhang H T,etal. The effects of La (III) on the peroxidation of membrane lipids in wheat seedling leaves under osmotic stress. Biological Trace Element Research, 1999, 69(2): 141-150.
[3] Liu J X, Wang R J, Wang X. Effects of lanthanum on physiological characteristics in ryegrass seedling roots under NaCl stress. Chinese Journal of Grassland, 2010, 32(6): 46-50. 刘建新, 王瑞娟, 王鑫. 镧对NaCl胁迫下黑麦草幼苗根系生理特性的影响. 中国草地学报, 2010, 32(6): 46-50.
[4] Xia H P, Ao H X. Brief introduction of the major grass in Taiwan, China. Grassland and Turf, 2000, (1): 43-45. 夏汉平, 敖惠修. 我国台湾的主要禾草简介. 草原与草坪, 2000, (1): 43-45.
[5] Long Z F, Wang J L, Liu Z S,etal. Effects of bahiagrass different planting modes on water and soil conservation. Journal of Mountain Agriculture and Biology, 2004, 23(5): 408-411. 龙忠富, 汪俊良, 刘正书, 等. 百喜草不同种植模式的水土保持效应初探. 山地农业生物学报, 2004, 23(5): 408-411.
[6] Chu G L, Fu F S, Wang J G,etal. A study on the application of ecological-economical bahiagrass. Jiangsu Agricultural Science, 2011, 39(6): 292-294. 储国良, 傅反生, 王敬根, 等. 生态节约型草坪百喜草应用技术研究. 江苏农业科学, 2011, 39(6): 292-294.
[7] Wang X K. The Experiment Principle and Technique on Plant Physiology and Biochemistry[M]. Beijing: Higher Education Press, 2006: 174-176. 王学奎. 植物生理生化实验原理和技术[M]. 北京: 高等教育出版社, 2006: 174-176.
[8] Zhang Q F, Xu S, Li J L. A study on variations of physiological and biochemical indices of high temperature stress inFestucaarundinaceacv. Barlexas. Pratacultural Science, 2006, 23(4): 26-28. 张庆峰, 徐胜, 李建龙. 高温胁迫下高羊茅生理生化特性研究. 草业科学, 2006, 23(4): 26-28.
[9] Zhu M H, Cai M Z, Wu S H,etal. Effect of phosphorus on element uptake and transportation in buckwheat under aluminum stress. Journal of Soil and Water Conservation, 2009, 20(3): 183-186. 朱美红, 蔡妙珍, 吴韶辉, 等. 磷对铝胁迫下荞麦元素吸收与运输的影响. 水土保持学报, 2009, 20(3): 183-186.
[10] Pan T G, Wang Y Z, Ke Y Q,etal. Effects of rare earth spraying on activities of ATPase and enzymes in the rhizosphere of sugarcane. Acta Agronomica Sinica, 1993, 19(2): 133-138. 潘廷国, 王元贞, 柯玉琴, 等. 喷施稀土对甘蔗植株ATP酶和根际土壤酶活性的影响. 作物学报, 1993, 19(2): 133-138.
[11] Chen Y M, Bai H Y, Li Y R. Application of lanthanum in main crops and its physiological and biochemical function. Guangxi Agricultural Sciences, 2003, (6): 15-17. 陈远孟, 白厚义, 李杨瑞. 镧在主要农作物的应用及其生理生化作用. 广西农业科学, 2003, (6): 15-17.
[12] Deng G F. Effect of La-rich rare earth on rice yield increasing and its application prospect. Guangxi Agricultural Sciences, 2006, 37(5): 553-555. 邓国富. 富镧稀土对水稻的增产效果及应用前景. 广西农业科学, 2006, 37(5): 553-555.
[13] Liu C. The Relationships between Physico-chemical Characteristics of Rare Earth Elements and Photosynthesis as well as Their Underlying Mechanisms[D]. Suzhou: Soochow University, 2010. 刘超. 稀土元素的理化特性与光合作用的关系及其作用机制[D]. 苏州: 苏州大学, 2010.
[14] Qman Y, Muhtar Z, Tayer A. Root activity and photosynthetic characteristics ofElaeagnusoxycarpaseedlings under drought stress.Chinese Journal of Applied Ecology, 2011, 22(7): 1789-1795. 齐曼·尤努斯, 木合塔尔·扎热, 塔衣尔·艾合买提. 干旱胁迫下尖果沙枣幼苗的根系活力和光合特性. 应用生态学报, 2011, 22(7): 1789-1795.
[15] Sechenbater, Wu H Y. Effect of different stress on roots activity and nitrate reductase activity inZeamaysL. Agricultural Research in the Arid Areas, 2001, 19(2): 67-70. 斯琴巴特尔, 吴红英. 不同逆境对玉米幼苗根系活力及硝酸还原酶活性的影响. 干旱地区农业研究, 2001, 19(2): 67-70.
[16] Wang Y F, Wang Q X, Shang L W. Effects of calcium on root activity and the content of osmotic adjustable organic substances of maize seedlings under NaCl stress. Journal of Maize Sciences, 2008, 16(2): 66-70. 王玉凤, 王庆祥, 商丽威. 钙对NaCl胁迫下玉米幼苗根系活力和有机渗透调节物质含量的影响. 玉米科学, 2008, 16(2): 66-70.
EffectoflanthanumonrootgrowthofPaspalumnotatumseedlingsunderaluminumstress
LUO Han-Fu1,2, SONG Hua-Wei1,2, LIU Tian-Zeng1,2, ZHANG Ju-Ming1,2*
1.CollegeofForestryandLandscapeArchitecture,SouthChinaAgriculturalUniversity,Guangzhou510642,China; 2.GuangdongEngineeringResearchCenterofGrasslandScience,Guangzhou510642,China
Lanthanum (La) is one of the most vital and active rare earth elements. It can promote seed germination, increase crop output, and enhance plant resistance to environmental stresses.Paspalumnotatum, a warm season turfgrass commonly used for conserving soil and water in Guangzhou, was selected to study the effect of La on root growth. Seven-day-old seedlings ofP.notatumwere grown in hydroponic conditions in an acid solution containing 0.9 μmol/L Al3+with and without 0.3 μmol/L La3+. Root morphology, the root dry weight/fresh weight ratio, and root activity were evaluated after 2, 4, and 6 d of treatment. The addition of La3+to the acidic Al3+solution resulted in increased axial root length, root surface area, number of root tips, root volume, and root activity ofP.notatumseedlings. These findings indicated that La can promote the root growth ofP.notatumseedlings under Al3+stress.
Paspalumnotatum; root system; aluminum stress; lanthanum
10.11686/cyxb2016475
http://cyxb.lzu.edu.cn
罗涵夫, 宋华伟, 刘天增, 张巨明. 铝胁迫条件下镧对百喜草幼苗根系生长的影响. 草业学报, 2017, 26(9): 208-213.
LUO Han-Fu, SONG Hua-Wei, LIU Tian-Zeng, ZHANG Ju-Ming. Effect of lanthanum on root growth ofPaspalumnotatumseedlings under aluminum stress. Acta Prataculturae Sinica, 2017, 26(9): 208-213.
2016-12-06;改回日期:2017-03-14
广东省科技计划项目(2012B020302002)资助。
罗涵夫(1992-),男,湖南郴州人,在读硕士。E-mail:82490247@qq.com*通信作者Corresponding author. E-mail:jimmzh@scau.edu.cn
猜你喜欢
现代园艺(2022年7期)2022-11-19
四川劳动保障(2021年3期)2021-06-09
今日农业(2020年13期)2020-08-24
哈哈画报(2018年4期)2018-09-10
现代园艺(2017年13期)2018-01-19
小火炬·智漫悦读(2017年7期)2017-09-29
中成药(2017年3期)2017-05-17
足球周刊(2016年15期)2016-11-02
中国农业文摘-农业工程(2016年5期)2016-04-12
金色年华(2016年8期)2016-02-28
