
嗜盐四联球菌及其在发酵食品中的应用
2017-09-22 05:30:07王博周朝晖李铁桥卢丽玲陈坚堵国成方芳
食品与发酵工业 2017年8期
王博,周朝晖,李铁桥,卢丽玲,陈坚,4,堵国成,方芳*
1(江南大学 生物工程学院,江苏 无锡,214122)2(工业生物技术教育部重点实验室,江苏 无锡,214122) 3(广东珠江桥生物科技股份有限公司,广东 中山,528415) 4(粮食发酵工艺与技术国家工程实验室,江苏 无锡,214122)
嗜盐四联球菌及其在发酵食品中的应用
王博1,2,周朝晖3,李铁桥3,卢丽玲3,陈坚1,2,4,堵国成1,4,方芳1,2*
1(江南大学 生物工程学院,江苏 无锡,214122)2(工业生物技术教育部重点实验室,江苏 无锡,214122) 3(广东珠江桥生物科技股份有限公司,广东 中山,528415) 4(粮食发酵工艺与技术国家工程实验室,江苏 无锡,214122)
嗜盐四联球菌是一类存在于酱油、鱼酱、豆制品等多种发酵食品及糖浆中的耐盐或耐高渗的乳酸菌。它们在食品发酵过程中可提高食品中有机酸、醛类和酯类等风味物质含量,具有广泛的应用价值。该文对嗜盐四联球菌的分类、生理生化特性、鉴定、胁迫环境下适应机制以及在发酵食品中的功能特性等几个方面的研究进展进行了综述。
发酵食品;嗜盐四联球菌;分类;鉴别
1 嗜盐四联球菌简介
嗜盐四联球菌(Tetragenococcushalophilus)是一类存在于发酵食品及糖浆中的嗜盐或嗜高渗的乳酸菌[1],早期被归类为足球菌属(Pediococcus),称为嗜盐足球菌(Pediococcushalophilus)或酱油足球菌(Pediococcussoyae)[2]。Collins等在研究足球菌与气球菌(Aerococcus)系统发育关系中时发现,有些菌株与所研究的足球菌序列同源性较低,将它们重新分类并命名为四联球菌属(Tetragenococcus)[3]。四联球菌属目前已经识别了5个种,包括嗜盐四联球菌(T.halophilus)、盐水四联球菌(T.muriaticus)、韩国四联球菌(T.koreensis)、弧四联球菌(T.solitarius)、嗜高渗四联球菌(T.osmophilus)[4]。嗜盐四联球菌与足球菌等乳酸菌的系统发育关系见图1,由16S rDNA系统发育树可以看出,嗜盐四联球菌与肠球菌和气球菌亲缘关系较近。
嗜盐四联球菌是高渗发酵食品中常见的四联球菌属的菌种,不同来源的菌株在环境耐受性和生理生化特性上有一定的区别。JUSTÉ等通过遗传分析方法,包括随机扩增多态性DNA标记(random amplified polymorphic DNA,RAPD)、16S rDNA系统发育分析和重复序列PCR(repetitive sequence-PCR,rep-PCR)技术,将从不同来源筛选得到的耐盐嗜盐四联球菌和耐高渗透压嗜盐四联球菌归类为2个种群[5]。根据抗高渗透压耐胁迫等表现型并结合RAPD基因类型,也可将嗜盐四联球菌分为2个亚种,即分离自糖浆的Tetragenococcushalophilussubsp.flandriensis亚种,和分离自发酵鱼类和发酵豆制品的Tetragenococcushalophilussubsp.halophilus亚种。其中T.halophilussubsp.flandriensis在糖浆浓度达到69 °Bx时仍可生长,而T.halophilussubsp.halophilus在浓度高于65 °Bx后便不能正常生长[4]。
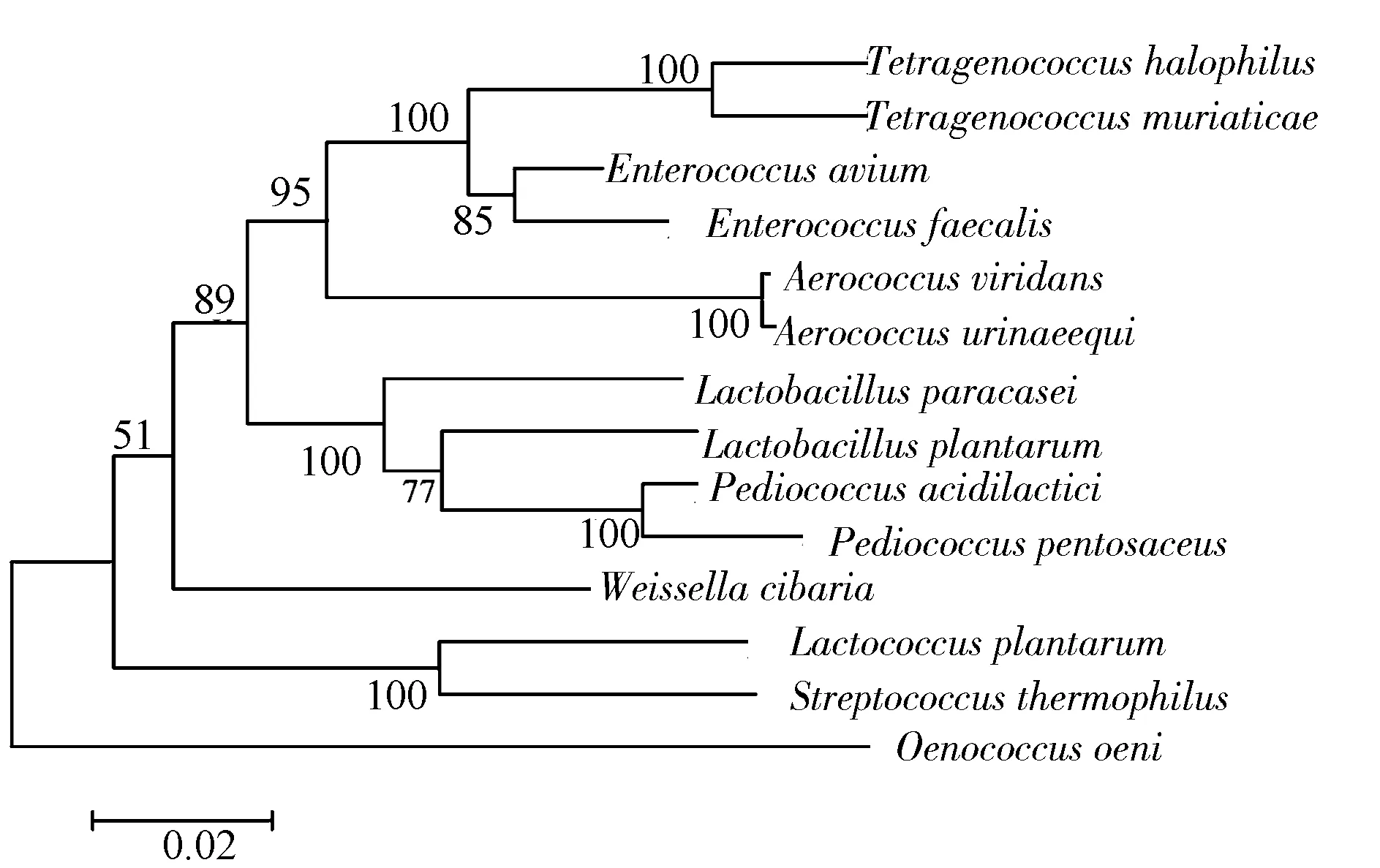
图1 基于16S rDNA测序构建嗜盐四联球菌系统发育树Fig.1 Phylogenetic tree of T.halophilus based on 16S rDNA sequencing
1.1嗜盐四联球菌的遗传特性和生理生化特性
嗜盐四联球菌是革兰氏阳性细菌,细胞不运动,非孢子形成球菌(直径约0.6~1 mm),显微镜下其细胞多呈四联球型,周围有胞外多糖,对提高菌株耐盐性有一定作用。菌株表现为氧化酶阴性,过氧化氢酶通常情况下为阴性,但部分嗜盐四联球菌在高铁血红素存在时,过氧化氢酶为阳性[6-8]。该属种的菌株为兼性厌氧,有氧条件不利于嗜盐四联球菌生长。嗜盐四联球菌菌落形态为白色,圆形,表面光滑,可在MRS液体培养基中缓慢生长(约3~5 d),在固体培养基中生长周期一般为5~7 d。其可生长的盐浓度范围为0%~25%(w/v),可生长温度范围为15~40 ℃,高于45 ℃不可生长,可在pH 5~9.6范围内生长。嗜盐四联球菌葡萄糖代谢终产物为乳酸,其基因组G+C含量为34%~37%[3-4]。
注:+:阳性;-:阴性。
通常所有嗜盐四联球菌均可发酵葡萄糖、果糖、甘露糖等糖类产酸;不可发酵淀粉、木糖等糖类;是否利用阿拉伯糖、甘油、棉子糖、海藻糖等碳源在不同菌株间情况不同。例如:分离自糖浆的嗜盐四联球菌均可发酵阿拉伯糖和乳糖产酸,而分离自腌鱼、酱油中的嗜盐四联球菌不能发酵这2种糖类,说明不同来源的嗜盐四联球菌对于糖类的发酵和利用有所区别[4]。此外,UDOMSIL等在泰国鱼酱中筛选到的嗜盐四联球菌M11、MS33、MRC10-1-3等菌株对于甘油和蔗糖的发酵情况也不相同。这说明相同来源的嗜盐四联球菌,不同菌株之间对于糖类发酵也有所区别[9]。
1.2嗜盐四联球菌的鉴定
嗜盐四联球菌与盐水四联球菌均为细胞是四联体形态的中度嗜盐乳酸球菌,共同存在于多种腌制品与发酵食品中。四联球菌属的这2种菌具有相似的生理特征与遗传学特征[10-12]。但是,嗜盐四联球菌一般在腌制品和发酵食品中起有益作用,盐水四联球菌则没有相关的有益功能。因此鉴别和区分2种菌并监测食品发酵过程及发酵体系中嗜盐四联球菌的存在具有重要意义。由于2种菌相似度较高,常用的细菌种属鉴定方法如16S rRNA基因序列比对分析较困难区分二者。KOBAYASHI等利用限制性内切酶片段长度多态性(restriction fragment length polymorphism,RFLP)方法成功区分了这2种菌[13],这种方法耗时较长且需使用特定仪器。KUDA等使用基质辅助激光解析电离飞行时间质谱(matrix-assisted laser desorption ionization-time of flight mass spectrometry,MALDI-TOF MS)有效地区分了这2种相近的四联球菌属的菌株:同种菌的质谱峰相似度高于78%,嗜盐四联球菌与盐水四联球菌的质谱峰相似度则为36%。采用这种方法获得的这2种四联球菌与其他乳酸球菌包括弧四联球菌的相似度均低于24%,可以高效且准确得将各菌种进行区分[14]。
2 嗜盐四联球菌与发酵食品
嗜盐乳酸菌是生产发酵食品的发酵剂的重要微生物组成,它们具有提高发酵食品产品质量或者缩短发酵周期等作用。嗜盐四联球菌是腌制类发酵食品中微生物的最重要组成之一,广泛存在于酱油、发酵豆制品、鱼酱和咸鱼等发酵食品中。将嗜盐四联球菌与碱性蛋白酶或风味蛋白酶结合加入发酵食品中,可以提高产品中挥发性物质和游离氨基酸的含量,起到提高产品风味的效果[15-16]。因此,了解嗜盐四联球菌对环境胁迫的适应机制,对于在发酵食品生产中应用嗜盐四联球菌,以及有针对性的改善食品风味和优化食品品质具有重要作用[17-20]。
2.1嗜盐四联球菌对环境胁迫的适应机制
发酵食品中,高的渗透压、酸胁迫等多重环境因素影响着微生物的生存与生理代谢。高渗透压环境使细胞内部水分流失,改变胞内物质浓度,影响细胞生理活动甚至造成细胞死亡。研究证明,嗜盐细菌通常有2种机制以适应外部环境的高渗胁迫:一是调节胞内离子浓度,即在胞内Na+含量升高时积累K+;二是细胞通过合成或者吸收相容性溶质来平衡环境中较高的离子浓度,相容性溶质包括氨基酸、糖类及其衍生物[21-23]。
与其他嗜盐菌相似,嗜盐四联球菌也是氯化物依赖性细菌。在高盐胁迫下,嗜盐四联球菌通过转运肉毒碱、甘氨酸甜菜碱或者将胆碱转化为甘氨酸甜菜碱来积累调渗物质[24]。热休克反应是存在于多种生物体内的应激反应,细胞在应激时合成的一组蛋白质为热休克蛋白。关于热休克蛋白编码基因在高渗透压条件下功能和表达的研究表明,GroESL,DnaK和ClpB充当功能性分子伴侣,提高了嗜盐四联球菌对高盐度的适应性[25-26]。LIU等在转录组学基础上对嗜盐四联球菌耐盐性进行了研究,结果表明,嗜盐四联球菌在不同盐浓度条件下,基因表达水平差异较大。在合适的盐浓度下,转录、翻译、膜系统、细胞分裂相关基因表达量明显提高,有助于细菌生长;在偏高的盐浓度下,表达量提高的主要为grpE,groES,clpE,hslV等编码热休克蛋白的基因和rpoD等编码ABC转运蛋白的基因。ABC转运蛋白利用ATP在细胞膜内外转运离子、糖类、氨基酸等多种亲水物质,可以起到调节渗透压的作用[23]。这些调节机制使嗜盐四联球菌具有适应外部高渗透压的能力,使其在发酵食品中有重要的应用价值。
酸性环境会影响微生物细胞膜的通透性和相关酶活,进而影响细胞的生长与代谢。研究证明,调节细胞膜脂肪酸组成是微生物应对外界胁迫因素的方法之一,而嗜盐四联球菌可通过提高细胞膜中不饱和脂肪酸含量来抵御酸胁迫[27-28]。氨基酸可以稳定细胞膜与蛋白质结构,避免细胞受到胁迫因素的迫害,例如:谷氨酸脱羧酶将胞外谷氨酸转化为γ-氨基丁酸,反应可消耗H+;精氨酸脱亚氨基(ADI)途径中,精氨酸被微生物转化为瓜氨酸和鸟氨酸,代谢产物包括NH3和ATP,都对细菌的耐酸性有一定贡献[29]。HE等证明以上氨基酸转化途径对增强嗜盐四联球菌耐酸性同样有作用,并通过研究证明,嗜盐四联球菌通过酸预适应处理后,耐酸性有显著提高[30]。
2.2嗜盐四联球菌对发酵食品风味的影响
风味物质是表征发酵食品的重要特征之一,主要包括芳香类风味物质和味感风味物质,根据化合物结构又可分为醇类、醛类、酯类、酮类、萜类化合物与氨基酸等。发酵食品的风味物质主要通过化学反应和生物方法合成。生物合成风味物质主要有2种途径,一是前体物质经过酶的催化作用转化为风味物质;二是利用微生物代谢葡萄糖、蛋白质等营养物质,来生产某些风味物质,即微生物发酵法生产风味物质[31]。
UDOMSIL等研究发现,嗜盐四联球菌在高盐环境中仍可水解蛋白质,并且具有胞内氨肽酶活性。在鱼酱发酵过程中添加嗜盐四联球菌可以显著提高鱼酱中总氨基酸和游离氨基酸的含量,使鲜味氨基酸,如谷氨酸等的含量有所提高。添加嗜盐四联球菌的鱼酱中,对风味有积极影响的挥发性物质如2-甲基丙醛、2-甲基丁醛、3-甲基丁醛和苯甲醛等的含量也明显增加[9,32]。WU从中国豆瓣酱中筛选到1株嗜盐四联球菌,并将其作为发酵剂接种至豆瓣酱发酵中用于强化豆瓣酱的风味。结果显示,接种嗜盐四联球菌后,豆瓣酱中的氨态氮、酸类、醇类等挥发性成分均有所提高,而亚硝酸盐含量减少了39.4%[7]。因此,通过添加嗜盐四联球菌,不仅可以提高发酵食品、调味品的风味,还能有效减少发酵食品中的有害物质。
很多发酵食品的生产过程都有酵母菌参与代谢。酵母菌以食品为原料的发酵代谢产物主要为醇类和醛类,嗜盐四联球菌的代谢产物则主要是乳酸等多种有机酸类,两者结合,可显著提高产品中酯类物质含量,起到改善产品风味的作用。例如在酱醪中强化酵母菌可以提高酱油中乙醇、2-苯乙醇、2-乙基丙酸酯、呋喃酮等物质含量,其中呋喃酮具有明显增香效果;而向酱醪中接种嗜盐四联球菌后,酱油中乙酸、甲酸、2-羟基丙酸甲酯、香草醛、甲硫基丙醛、糠醛和糠醇等挥发性成分含量都有提高。糠醛和糠醇分别具有甜味和焦糖风味,酱醪体系中由于接种嗜盐四联球菌使得有机酸含量提高,加速了糠醛的生成[33-36];CUI等通过先接种嗜盐四联球菌,后接种耐盐酵母菌的方式,使酱醪中醇类、酮类、吡嗪类物质含量明显提高,证实了嗜盐四联球菌对增加风味物质含量的贡献程度要大于酵母菌,将它与酵母菌结合使用,风味物质含量增加比例最高[37]。同样,将嗜盐四联球菌与酵母菌结合作为发酵剂发酵豆豉,除增加挥发性风味成分外,还可缩短发酵周期[38]。以上结果表明,嗜盐四联球菌在各类发酵食品中均有提高产品风味的作用,也可以与酵母菌等微生物组合使用,可以作为优良的发酵剂,具有广阔的应用前景。
2.3嗜盐四联球菌对发酵食品安全的影响
水产品和肉制品在发酵过程中由于乳酸菌对含氮物质的不完全利用会产生氨基甲酸乙酯、生物胺等胺类物质。布氏乳杆菌、短乳杆菌、明串珠菌属和足球菌都可产氨基脱羧酶导致发酵过程中产生组胺、色胺和酪胺等生物胺[39],多种乳酸菌中存在的组氨酸脱羧酶可催化组氨酸脱羧生成组胺。酪胺可导致人体高血压,组胺可引起一些过敏反应,国家标准规定组胺含量:鲐鱼≤100 mg/100g、其他鱼类≤30 mg/100 g;上海市地方标准暂定发酵肉制品中组胺限量为≤200 mg/kg[40]。氨基甲酸乙酯为2A类致癌物质(具有潜在致癌性),前体物质为尿素、氨甲酰磷酸、瓜氨酸等。而一些乳酸菌可以通过精氨酸脱亚氨基途径(Arginine deiminase pathway,ADI)将精氨酸转化为瓜氨酸,有潜在的危害性[8]。降低发酵食品中有害物质的含量有助于保障食品的安全性,所以应关注发酵食品中微生物的氨基酸代谢与有害物质形成之间的联系。
已经报道的嗜盐四联球菌全基因组中有完整的ADI代谢途径,廖淡宜在研究中发现筛选自酱醪的嗜盐四联球菌R23可以在高盐的环境中将精氨酸和瓜氨酸转化为鸟氨酸,对瓜氨酸的利用率为17.7%,瓜氨酸到鸟氨酸的转化率为100%,既可以降低有害物质含量,也可以提高产品风味[8]。但是不同嗜盐四联球菌菌株之间ADI代谢途径的基因型与表现型有一定区别,并且在实际应用期间受到低温等环境因素影响,降低氨基甲酸乙酯及其前体物质瓜氨酸的能力有限,需要进一步研究选择合适的菌株应用于生产[41]。
SATOMI等分析嗜盐四联球菌在含有1%L-组氨酸的组氨酸培养基中可产生3 mg/mL的组胺[42]。UDOMSIL等研究发现嗜盐四联球菌在含有5%~25%氯化钠和0.25%组氨酸的改良GYP培养基中可合成0.06%~0.22 mg/mL的组胺,因此将该菌归类为中等组胺产生菌[9]。JEONG等对嗜盐四联球菌进行了安全评估,结果表明嗜盐四联球菌不同菌株产酪胺水平差别较大[43]。在鱼酱的发酵中添加嗜盐四联球菌,则可使样品中组胺和酪胺含量均减少,减少鱼酱中生物胺产生[32]。KUDA等从发酵水产品中筛选到两株嗜盐四联球菌HmF-131和HmS-129,HmF-131可使沙丁鱼在发酵过程中组胺含量升高,HmS-129则使组胺含量降低[44]。上述研究结果表明,合适的嗜盐四联球菌菌株不仅不会增加发酵食品中的生物胺,还能减少发酵食品中某些生物胺的生成。因此,发酵食品中组胺的产生可能与体系中其他微生物的氮源代谢相关。在实际生产中可选择能降低生物胺含量的嗜盐四联球菌株作为发酵剂,提高发酵食品的安全性。
2.4嗜盐四联球菌的益生作用
乳酸菌广泛存在于人体中,可以起到调节肠胃正常菌群、控制内毒素、抑制有害微生物及物质产生等多种益生作用,对人体的营养、生理功能和免疫反应等具有重要意义。近年来,一些报道指出嗜盐四联球菌可起到促进健康的益生菌作用。MASUDA等研究表明分离自酱油的嗜盐四联球菌Th221有助于调高Th1型免疫应答。通过体外和体内实验发现,嗜盐四联球菌可以促进小鼠腹腔巨噬细胞产生抗肿瘤及抗感染的白细胞介素-12,并随着介质中含盐量增加(≥10%,w/v),这一促进作用增强[45]。NISHIMURA等将嗜盐四联球菌Th221应用于治疗常年性变应性鼻炎(TAR)时发现,口服高剂量(60 mg/d的Th221实验组在实验结束后患者鼻炎严重程度和鼻塞症状有明显好转,试验期间患者的喷嚏流涕症状也有所改善,说明嗜盐四联球菌Th221可以改善TAR症状[46],有应用于改善过敏症状的潜力。
3 结论
对来源于发酵食品的嗜盐四联球菌的遗传特性和生理生化特性的深入研究,一方面可以丰富研究者关于耐盐菌的知识,另一方面可以为探究嗜盐四联球菌在食品发酵过程中发挥的作用与利用价值提供理论支持。最近对嗜盐四联球菌的研究表明,嗜盐四联球菌在高盐的发酵食品中可以提高挥发性成分含量,减少生物胺类有害物质的含量。但嗜盐四联球菌不同菌株间生理特性以及物质代谢情况有所差异,选择合适的菌株可以强化食品风味,提高食品安全性,甚至赋予食品一定保健功能,所以需要对其进行进一步的系统研究,以更好的应用于发酵食品生产。
[1] WILLEMS K A,WILLEMS M L,DARDENNE F,et al. Microbiological observations during storage of thick juice on a pilot and industrial scale[C].Cits Proceedings,Madrid 2003. Proceedings of theGeneral Assembly of the International Commission for Sugar Technology,Madrid,Spain,2003:18-21.
[2] TANASUPAWAT S,DAENGSUBHA W.Pediococcusspecies and related bacteria found in fermented foods and related materials in Thailand[J]. Journal of General & Applied Microbiology,1983,29(6):487-506.
[3] COLLINS M D,WILLIAMS A M,WALLBANKS S. The phylogeny ofAerococcusandPediococcusas determined by 16S rRNA sequence analysis: description ofTetragenococcusgen. nov.[J]. Fems Microbiology Letters,1990,70(3):255-262.
[4] JUSTÉ A,VAN T S,VERRETH C,et al. Characterization ofTetragenococcusstrains from sugar thick juice reveals a novel species,Tetragenococcusosmophilussp. nov.and dividesTetragenococcushalophilusinto two subspecies,T.halophilussubsp.halophilussubsp. nov.andT.halophilussubsp.flandriensissubsp.nov.[J]. International Journal of Systematic & Evolutionary Microbiology,2012,62(Pt 1):129-137.
[5] JUSTÉ A,LIEVENS B,FRANS I,et al. Genetic and physiological diversity ofTetragenococcushalophilusstrains isolated from sugar- and salt-rich environments[J]. Microbiology,2008,154(9):2 600-2 610.
[6] GURTLER M,GANZLE M G,WOLF G,et al. Physiological diversity among strains ofTetragenococcushalophilus[J]. Systematic and Applied Microbiology,1998,21(1): 107-112.
[7] WU C D,LIU C L,HE G Q,et al. Characterization of a multiple-stress toleranceTetragenococcushalophilusand application as starter culture in Chinese horsebean-chili-paste manufacture for quality improvement[J]. Food Science &Technology Research,2013,19 (5):855-864.
[8] 廖淡宜,方芳,陈坚,等. 嗜盐四联球菌的分离及其精氨酸代谢[J]. 食品与生物技术学报,2016,35(1):48-53.
[9] UDOMSIL N,RODTONG S,TANASUPAWAT S,et al. Proteinase-producing halophilic lactic acid bacteria isolated from fish sauce fermentation and their ability to produce volatile compounds[J]. International Journal of Food Microbiology,2010,141(3):186-194.
[10] KUDA T,MIHARA T,YANO T. Detection of histamine and histamine-related bacteria in fish-nukazuke,a salted and fermented fish with rice-bran,by simple colorimetric microplate assay[J]. Food Control,2007,18(6):677-681.
[11] KUDA T,OKAMOTO K,YANO T. Population of halophilic bacteria in salted fish products made in the Loochoo Islands,Okinawa and the Noto Peninsula,Ishikawa,Japan[J]. Fisheries Science,2002,68(6):1 265-1 273.
[12] SATOMI M,KIMURA B,MIZOI M,et al.Tetragenococcusmuriaticussp. nov.a new moderately halophilic lactic acid bacterium isolated from fermented squid liver sauce[J]. International Journal of Systematic Bacteriology,1997,47(3):832.
[13] KOBAYASHI T,KIMURA B,FUJII T. Differentiation ofTetragenococcuspopulations occurring in products and manufacturing processes of puffer fish ovaries fermented with rice-bran[J]. International Journal of Food Microbiology,2000,56(2):211-218.
[14] KUDA T,IZAWA Y,YOSHIDA S,et al. Rapid identification ofTetragenococcushalophilusandTetragenococcusmuriaticus, important species in the production of salted and fermented foods,by matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS)[J].Food Control,2014,35 (1): 419-425.
[15] OGASAWARA M,YAMADA Y,EGI M. Taste enhancer from the long-term ripening of miso (soybean paste)[J]. Food Chemistry,2006,99(4):736-741.
[16] YONGSAWATDIGUL J,RODTONG S,RAKSAKULTHAI N. Acceleration of Thai fish sauce fermentation using proteinases and bacterial starter cultures[J]. Journal of Food Science,2007,72(9): 382-390.
[17] KIM T W,LEE J H,KIM S E,et al. Analysis of microbial communities in doenjang,a Korean fermented soybean paste,using nested PCR-denaturing gradient gel electrophoresis[J]. International Journal of Food Microbiology,2009,131(2-3):265-271.
[18] KUDA T,IZAWA Y,ISHII S,et al. Suppressive effect ofTetragenococcushalophilus,isolated from fish- nukazuke,on histamine accumulation in salted and fermented fish[J]. Food Chemistry,2012,130(3):569-574.
[19] ROLING W,VAN VERSEVELD H W. Characterization ofTetragenococcushalophilapopulationsin Indonesian soy mash (Kecap) fermentation[J]. Applied & Environmental Microbiology,1996,62(4):1 203-1 207.
[20] SUGIYAMA S. Selection of micro-organisms for use in the fermentation of soy sauce[J]. Food Microbiology,1984,1(4):339-347.
[22] GUNDE-CIMERMAN N,GUNDE-CIMERMAN N,GUNDE-CIMERMAN N,et al. Adaptation to life at high salt concentrations in archaea,bacteria,and eukarya[M]. Springer Netherlands,2005,48(11):2 181-2 186.
[23] LIU L C,SI L F,MENG X,et al. Comparative transcriptomic analysis reveals novel genes and regulatory mechanisms ofTetragenococcushalophilusin response to salt stress[J]. Journal of Industrial Microbiology & Biotechnology,2015,42(4):601-616.
[24] ROBERT H,LE M C,BLANCO C,et al. Glycine betaine,carnitine,and choline enhance salinity tolerance and prevent the accumulation of sodium to a level inhibiting growth ofTetragenococcushalophila[J]. Applied & Environmental Microbiology,2000,66(2):509-517.
[25] SUGIMOTO S,SARUWATARI K,HIGASHI C,et al. In vivo and in vitro complementation study comparing the function of DnaK chaperone systems from halophilic lactic acid bacteriumTetragenococcushalophilusandEscherichiacoli[J]. Bioscience Biotechnology & Biochemistry,2008,72(3):811-822.
[26] SUGIMOTO S,YOSHIDA H,MIZUNOE Y,et al. Structural and functional conversion of molecular chaperone ClpB from the gram-positive halophilic lactic acid bacteriumTetragenococcushalophilusmediated by ATP and stress[J]. Journal of Bacteriology,2006,188(23):8 070-8 078.
[27] DENICH T J,BEAUDETTE L A,LEE H,et al. Effect of selected environmental and physico-chemical factors on bacterial cytoplasmic membranes[J]. Journal of Microbiological Methods,2003,52(2):149-182.
[28] 何桂强,冯笑,吴重德,等. 多重胁迫对嗜盐四联球菌CGMCC 3792存活率及细胞成分的影响[J]. 食品工业科技,2016,37(5):182-186.
[29] TEIXEIRA J S,SEERAS A,SANCHEZ-MALDONADO A F,et al. Glutamine,glutamate,and arginine-based acid resistance inLactobacillusreuteri[J]. Food Microbiology,2014,42:172-180.
[30] HE G Q,WU C D,HUANG J,et al. Acid tolerance response ofTetragenococcushalophilus: A combined physiological and proteomic analysis[J]. Process Biochemistry,2015,51(2):213-219.
[31] 陈妍,陆利霞,熊晓辉. 微生物发酵法生产食品风味物质[J].中国调味品,2011,36(7):13-17.
[32] UDOMSIL N,RODTONG S,CHOI Y J,et al. Use ofTetragenococcushalophilusas a starter culture for flavor improvement in fish sauce fermentation[J]. Journal of Agricultural and Food Chemistry,2011,59(15):8 401-8 408.
[33] LEE K E,LEE S M,CHOI Y H,et al. Comparative volatile profiles in soy sauce according to inoculated microorganisms[J]. Bioscience,Biotechnology and Biochemistry,2013,77 (11): 2 192-2 200.
[34] HARADA R,YUZUKI M,ITO K,et al. Influence of yeast and lactic acid bacterium on the constituent profile of soy sauce during fermentation[J]. Journal of Bioscience & Bioengineering,2017,92(2):203-208.
[35] NOSAKA S,MIYAZAWA M. Characterization of volatile components and odor-active compounds in the oil of edible mushroom Boletopsisleucomelas[J]. Journal of Oleo Science,2014,63(6):577.
[36] LEE S M,SEO B C,KIM Y S. Volatile compounds in fermented and acid-hydrolyzed soy sauces[J]. Journal of Food Science,2006,71(3):146-156.
[37] CUI R Y,ZHENG J,WU C D,et al. Effect of different halophilic microbial fermentation patterns on the volatile compound profiles and sensory properties of soy sauce moromi[J]. European Food Research and Technology,2014,239(2):321-331.
[38] HE G Q,HUANG J,LIANG R,et al. Comparing the differences of characteristic flavour between natural maturation and starter culture for Mucor-type Douchi[J]. International Journal of Food Science and Technology,2016,51(5): 1 252-1 259.
[39] BOVER-CID S,HOLZAPFEL W H. Improved screening procedure for biogenic amine production by lactic acid bacteria[J]. International Journal of Food Microbiology,1999,53(1):33-41.
[40] 刘景,任婧,孙克杰. 食品中生物胺的安全性研究进展[J]. 食品科学,2013,34(5):322-326.
[41] 廖淡宜. 酱醪中嗜盐四联球菌的分离及特性研究[D]. 无锡:江南大学,2015.
[42] SATOMI M,FURUSHITA M,OIKAWA H,et al. Analysis of a 30 kbp plasmid encoding histidine decarboxylase gene inTetragenococcushalophilusisolated from fish sauce[J]. Fisheries Science,2014,80(4):849-858.
[43] JEONG D W,HEO S,LEE J H. Safety assessment ofTetragenococcushalophilusisolates from doenjang,a Korean high-salt-fermented soybean paste[J]. Food Microbiology,2016,62:92-98.
[44] KUDA T,IZAWA Y,ISHII S,et al. Suppressive effect ofTetragenococcushalophilus, isolated from fish-nukazuke,on histamine accumulation in salted and fermented fish[J].Food Chemistry,2012,130 (3): 569-574.
[45] MASUDA S,YAMAGUCHI H,KUROKAWA T,et al.Immunomodulatory effect of halophilic lactic acid bacteriumTetragenococcushalophilusTh221 from soy saucemoromi grown in high-salt medium[J].International Journal of Food Microbiology,2008,121 (3): 245-252.
[46] NISHIMURA I,IGARASHI T,ENOMOTO T,et al.Clinical efficacy of halophilic lactic acid bacteriumTetragenococcushalophilusTh221from soy sauce moromi for perennial allergic rhinitis[J].Allergology International,2009,58(2):179-185.
Tetragenococcushalophilusanditsapplicationinfermentedfoods
WANG Bo1,2,ZHOU Zhao-hui3,LI Tie-qiao3,LU Li-ling3,CHEN Jian1,2,4,DU Guo-cheng1,4,FANG Fang1,2*
1(School of Biotechnology,Jiangnan University,Wuxi 214122,China) 2(Key Laboratory of Industrial Biotechnology,Ministry ofEducation,School of Biotechnology,Wuxi 214122,China) 3(Guangdong Pearl River Bridge Biological Limited Corporation,Zhongshan 528415,China) 4(Laboratory for Cereal Fermentation Technology,Wuxi 214122,China)
Tetragenococcushalophilusis a kind of halophilic or osmophilic lactic acid bacteria,which widely presents in soy sauce,fish sauce,bean products,sugar thick juice and other fermented foods.Strains from this species are proved to play important roles in formation of flavor substances such as organic acids,aldehydes and esters during food fermentation process.In this review,classification and identification ofT.halophilus,the physiological and biochemical characteristics,stress response as well as the functional properties ofT.halophilusin fermented foods were discussed.
fermented foods;Tetragenococcushalophilus; taxonomy;characterization
10.13995/j.cnki.11-1802/ts.014014
硕士研究生(方芳副教授为通讯作者,E-mail:ffang@jiangnan.edu.cn)。
国家自然科学基金(31371821);广东省科技计划项目(2015B020205002);江南大学自主科研计划重点项目(JUSRP51734B)
2017-02-10,改回日期:2017-04-21
猜你喜欢
天津农学院学报(2024年1期)2024-04-07 11:20:56
新世纪智能(高一语文)(2020年10期)2021-01-04 00:43:56
新世纪智能(高一语文)(2020年10期)2020-12-31 09:26:20
少儿美术(快乐历史地理)(2020年7期)2020-11-26 06:25:50
心肺血管病杂志(2019年4期)2019-06-27 07:36:18
国际呼吸杂志(2019年1期)2019-01-28 09:37:06
家庭影院技术(2019年1期)2019-01-21 02:25:02
现代检验医学杂志(2016年2期)2016-11-14 02:38:02
环境科技(2016年3期)2016-11-08 12:14:10
中国老区建设(2016年5期)2016-02-28 09:32:48
