
Cu2+、Zn2+对西藏不同区域野生赖草萌发特性和生理效应的影响
2017-09-13 05:19徐雅梅张卫红蔺永和刘大林苗彦军
草地学报 2017年5期
徐雅梅, 张卫红, 蔺永和, 刘大林, 苗彦军*
(1. 西藏农牧学院, 西藏 林芝 86000; 2. 扬州大学动物科学与技术学院, 江苏 扬州225009; 3. 兰州大学, 甘肃 兰州 730000)
西藏位于我国西南边陲,作为青藏高原的主体,具有“世界第三极”之称。该区域具有丰富的矿产资源,然而矿产开采过程中,矿区及周边土壤往往会受到Cu、Zn、Pb、Cd等重金属不同程度的污染[1-3],加之设施农业发展过程中化肥、农药的大量施用[4]及交通排放[5-6]等因素致使土壤遭到Cu、Zn等重金属的污染[7]。土壤系统中的重金属污染和防治一直是国际上研究的难点和热点[8],尤其是受青藏高原独特环境的影响,生态系统十分脆弱,一经破坏,不易恢复[9]。因此,对矿迹地和重金属污染区进行恢复治理刻不容缓,而进行重金属污染土壤恢复治理的一个重要途径是选育具有优良抗性的植物[10]。
赖草(Leymussecalinus)属于禾本科多年生草本植物,主要分布于我国华北、东北、西北和西南地区[11];具有广泛适应性和较高的抗逆性,能够适应青藏高原严酷的生存环境,是当地的重要优势乡土植物之一[12]。赖草具有强壮的根茎和迅速的分蘖繁殖能力,其在生态恢复过程中易形成单优群落[13];此外,赖草还具有适口性好、营养价值高和易被家畜所喜食等诸多优良特性,是重要的牧草资源[14]。目前,国内外关于赖草耐旱、耐寒及耐盐碱等抗逆性研究方面做了大量工作[12,15-16],但关于赖草在重金属胁迫下的响应研究并不多见,尤其是关于西藏不同区域野生赖草种子资源抗重金属离子胁迫的研究还未见详细报道。因此,本研究以采自西藏4个不同区域的赖草作为试验材料,通过观测不同浓度的重金属离子(Cu2+、Zn2+)处理下各赖草种子萌发、生长状况及生理指标的变化,以期为赖草在西藏重金属污染地治理中的运用和西藏赖草种子资源的评价与利用提供科学参考依据。
1 试验设计
1.1 试验材料
供试材料为采集于西藏4个不同区域的赖草种子,其编号和生境情况如表1所示,供试试剂为CuSO4·5H2O和ZnSO4·7H2O(分析纯)。
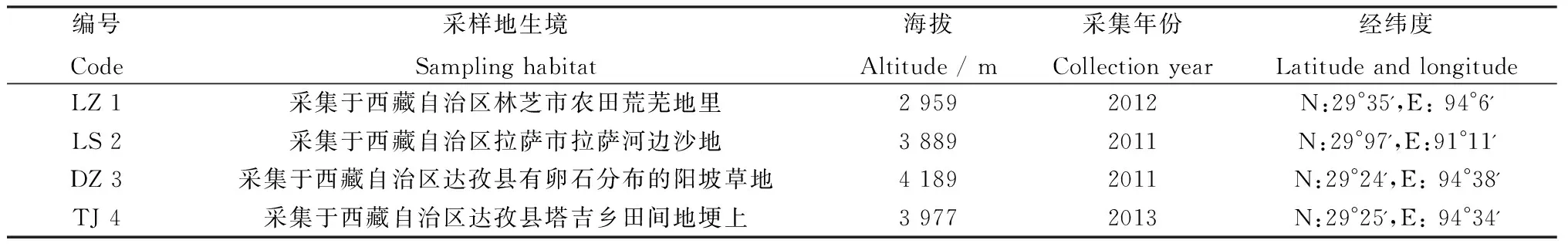
表1 试验种子编号及生境Table 1 Test seed code and habitat
1.2 试验方法
选取籽粒均匀饱满一致的4种赖草种子,用0.1%的高锰酸钾溶液消毒15 min,接着用蒸馏水冲洗3次,用滤纸吸干种子表面水分,接着在蒸馏水中浸泡8 h,使种子充分吸水。然后在发芽盒中放置2层滤纸,用蒸馏水做对照,每个发芽盒中分别用胶头滴管加Cu2+和Zn2+浓度分别为20,50,100,200,300,400,500 mg·L-1的CuSO4溶液和ZnSO4溶液至滤纸饱和(形成一层水膜)为止,然后将浸泡好的种子均匀摆置到发芽盒中,每种赖草种子100粒,每个重金属离子浓度梯度设置5个重复。从第二天开始,每日9:30和21:30用胶头滴管分别加对应溶液至滤纸饱和,使重金属离子浓度保持基本一致。
1.3 测定指标及方法
1.3.1萌发指标的测定 从第2 d开始,每天记录各处理下赖草种子萌发数,到第7 d时,以胚根露出种皮为萌发标志[17],统计发芽势(Gp)。发芽试验结束时(14 d)统计发芽率(Gr),测定胚芽长和胚根长,每个处理测5株;并将胚根、胚芽分开,用电子天平(精确到0.0001)称量胚根、胚芽鲜重,计算各处理下的相对发芽势、相对发芽率、活力指数。计算公式如下[18-19]:
① 发芽势(Gp)=前7 d累计发芽种子数/供试种子总数×100%
发芽率(Gr)=试验结束累计发芽种子数/供试种子总数×100%
② 发芽指数(GI)=处理后t日的发芽数/相应的发芽时间。
③ 根系耐受指数=(某浓度下的胚根长/对照处理的胚根长)×100%
④ 活力指数(V)=发芽指数×生物量(生物量=胚芽鲜重+胚根鲜重)。
1.3.2生理指标的测定 采用硫代巴比妥酸法[20]测定胚芽中丙二醛(MDA)的含量。
1.4 数据分析
采用Excel 2013对试验数据预处理后;再用SPSS 19.0进行方差分析,Duncan法进行多重比较;Excel 2013作图。
2 结果与分析
2.1 Cu2+、Zn2+对赖草发芽势的影响
由图1可知,不同重金属(Cu2+、Zn2+)对采自西藏4个不同地域的赖草发芽势的影响不同,同种重金属不同浓度对其发芽势的影响也不相同。其中LZ 1和TJ 4的发芽势随着Cu2+浓度的增加而减小;而LS 2和DZ 3的发芽势则均表现出先增大后减小的变化趋势,两者均在Cu2+浓度为20 mg·L-1时达到试验组最大值,依次为75.5%和44.71%,当Cu2+浓度超过20 mg·L-1时,发芽势随着Cu2+浓度的增大而显著减小(P< 0.05)。当Cu2+浓度为100 mg·L-1时,TJ 4的发芽势为0,Cu2+浓度为200 mg·L-1时,4个赖草的发芽势均为0。
Zn2+处理下,各赖草的发芽势均表现出低浓度促进,高浓度抑制的变化规律(图1)。LZ 1和LS 2在Zn2+浓度20 mg·L-1时发芽势均最高,依次为25.24%和72.67%;当Zn2+浓度超过20 mg·L-1之后,其发芽势随着Zn2+浓度的增大而减小。DZ 3和TJ 4在Zn2+浓度为100 mg·L-1时发芽势均达最高值,依次为43.28%和13.89%,当Zn2+浓度超过100 mg·L-1之后,发芽势随着Zn2+浓度的增大而减小。在同一浓度重金属离子(Cu2+、Zn2+)处理下,采自不同地方的赖草发芽势之间也表现出显著性差异(P< 0.05)。
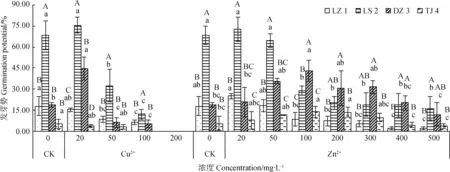
图1 Cu2+、Zn2+对赖草发芽势的影响Fig. 1 Effect of Cu2+ and Zn2+ on the germination potential of Leymus secalinus注:图中不同大写字母表示同一金属浓度下不同来源赖草之间存在显著性差异(P<0.05);不同小写字母表示同种赖草在不同金属浓度之间存在显著性差异(P < 0.05),下同Note: Different capital letters indicate significant difference between the same metal concentration under different secalinus (P<0.05); different lowercase letters indicate significant differences between different metal concentrations of the same secalinus (P < 0.05); the same below
2.2 Cu2+、Zn2+对赖草发芽率的影响
由图2可知,不同浓度Cu2+处理下,LZ 1和TJ 4的发芽率随着Cu2+浓度的增大而降低,说明试验浓度下Cu2+对这两种赖草的萌发具有抑制作用;而LS 2和DZ 3的发芽率随着Cu2+浓度的增大呈现先上升后下降的变化趋势,均在Cu2+浓度20 mg·L-1时发芽率最高,依次为86%和48.4%;Cu2+浓度超过20 mg·L-1时,发芽率呈现下降趋势,且低于对照组发芽率。说明20 mg·L-1Cu2+浓度对LS 2和DZ 3的萌发具有促进作用,而Cu2+浓度超过20 mg·L-1时对这两种赖草萌发表现出抑制作用。
Zn2+处理下,各赖草发芽率均随着Zn2+浓度的增大呈先增高后降低的变化趋势(图2)。其中LZ 1和LS 2的发芽率均在Zn2+浓度50 mg·L-1时达到实验组最大值,依次为53.89%和87.42%,均高于对照组和Zn2+浓度20 mg·L-1时的发芽率;当Zn2+浓度大于等于100 mg·L-1时,发芽率迅速降低。表明Zn2+浓度低于50 mg·L-1时对发芽率具有促进作用,而Zn2+浓度大于等于100 mg·L-1时,则对其发芽率具有抑制作用。DZ 3和TJ 4在Zn2+浓度20 mg·L-1时的发芽率最高,超过20 mg·L-1时逐渐降低。但DZ 3的发芽率在Zn2+浓度200 mg·L-1时,仍显著高于对照组(P<0.05),Zn2+浓度达到300 mg·L-1时才低于对照。表明200 mg·L-1Zn2+浓度对赖草的发芽率具有促进作用,Zn2+浓度≥300 mg·L-1时表现为抑制作用。TJ 4的发芽率直至Zn2+浓度500 mg·L-1时仍高于对照,说明Zn2+浓度在20~500 mg·L-1之间均对其发芽率具有促进作用。

图2 Cu2+、Zn2+对赖草发芽率的影响Fig.2 Effect of Cu2+ and Zn2+ on the germination rate of Leymus secalinus
2.3 Cu2+、Zn2+对赖草发芽指数的影响
不同浓度的重金属离子(Cu2+、Zn2+)对采自西藏4个不同地域赖草的发芽指数有不同的影响,整体表现为低浓度提高其发芽指数,高浓度降低其发芽指数(图3)。Cu2+处理下,LZ 1和TJ 4的发芽指数随着Cu2+浓度的升高而降低,而LS 2和DZ 3的发芽指数随着Cu2+浓度的升高呈先增大,在Cu2+浓度为20 mg·L-1时达到最大值,当Cu2+浓度超过20 mg·L-1时呈现下降趋势。随Zn2+浓度增加,各赖草发芽指数均表现为先增大后减小的变化趋势,其中LZ 1和LS 2在Zn2+浓度为50 mg·L-1时发芽指数最高,DZ 3和TJ 4在Zn2+浓度为20 mg·L-1时发芽指数最高,此后发芽指数随着Zn2+浓度增大而降低。
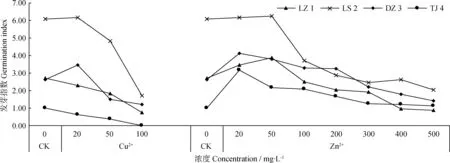
图3 Cu2+、Zn2+对赖草发芽指数的影响Fig.3 Effect of Cu2+ and Zn2+ on the germination index of Leymus secalinus
2.4 Cu2+、Zn2+对赖草胚芽的影响
2.4.1Cu2+、Zn2+对赖草胚芽长度的影响 由图4可知,不同浓度Cu2+处理下,LZ 1和TJ 4的胚芽长度均随着Cu2+浓度的增大而减少;LS 2和DZ 3的胚芽长随着Cu2+浓度的增大呈现先减少后增高的变化趋势,Cu2+浓度由0 mg·L-1增加至50 mg·L-1的时,胚芽长度随着Cu2+浓度的增大而降低,表现出同LZ 1和TJ 4相同的变化规律,而Cu2+浓度为100 mg·L-1时,LS 2和DZ 3的胚芽长度稍微有所增加,但差异不显著。不同浓度Zn2+处理下,4个赖草的胚芽长度整体上呈先增大后减小的变化趋势,但有些赖草在某些浓度上有所差别。其中Zn2+浓度为20 mg·L-1时,各赖草胚芽长度均大于其他处理下的胚芽长度;Zn2+浓度由20 mg·L-1增加到200 mg·L-1的过程中,DZ 3和TJ 4的胚芽长度随Zn2+浓度的增大而降低;300 mg·L-1Zn2+浓度处理下的胚芽较200 mg·L-1的胚芽长度有所增加;Zn2+浓度超过300 mg·L-1时,胚芽长度又呈现递减趋势。LZ 1和LS 2的胚芽长度由20 mg·L-1增加到400 mg·L-1的过程中呈递减变化趋势,但500 mg·L-1Zn2+浓度下较400 mg·L-1时的胚芽长度却稍有所增加。
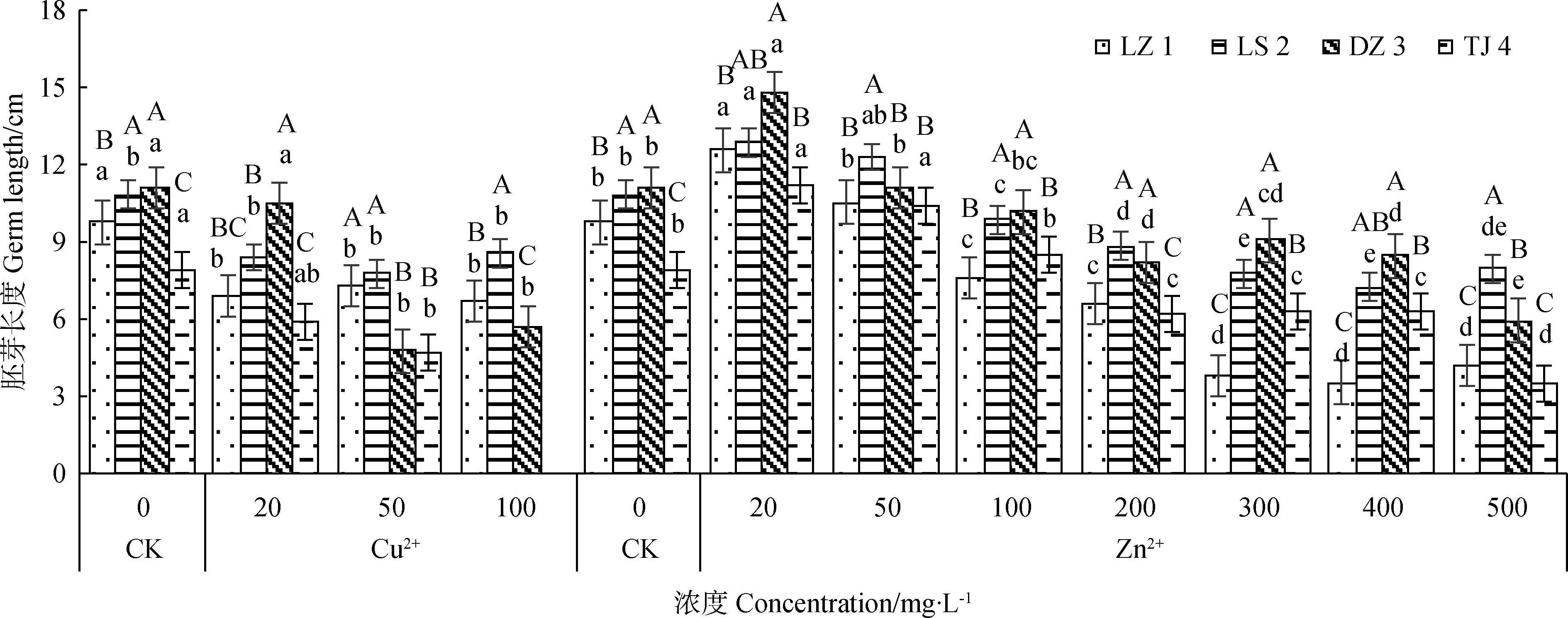
图4 Cu2+、Zn2+对赖草胚芽长的影响Fig.4 Effect of Cu2+ and Zn2+ on germ length of Leymus secalinus
2.4.2Cu2+、Zn2+对赖草胚芽鲜重的影响 由图5可知,Cu2+处理下,4个赖草的胚芽鲜重整体上均随着Cu2+浓度的升高而降低。LS 2和DZ 3在Cu2+浓度为100 mg·L-1时的胚芽鲜重较50 mg·L-1有所增加,但仍小于20 mg·L-1时的胚芽鲜重。Zn2+处理下,4个赖草的胚芽鲜重整体上均呈先增大后减小的变化规律。其中Zn2+浓度50 mg·L-1时,LS 2的胚芽鲜重大于其他浓度处理下的胚芽鲜重,为0.1136 g,表明该浓度对LS 2胚芽生长的促进作用最强;而其他3个赖草均在Zn2+浓度为20 mg·L-1时就达到试验组最大值。
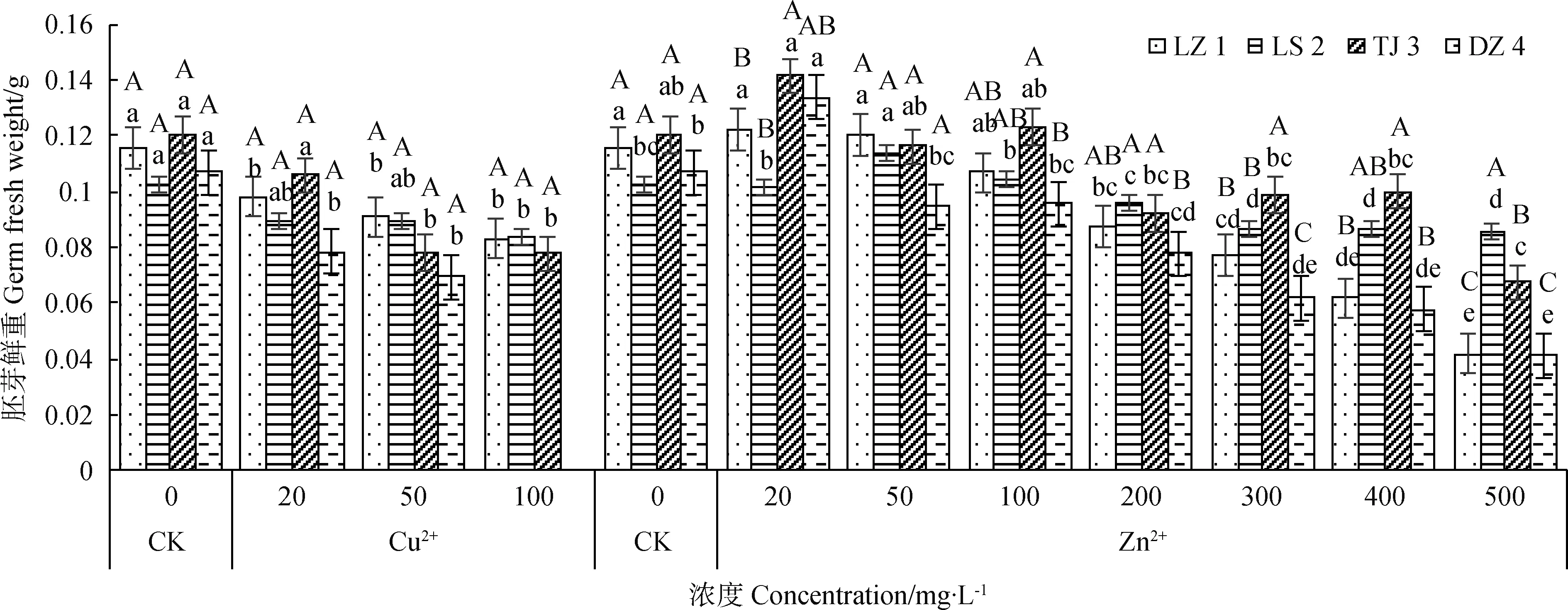
图5 Cu2+、Zn2+对赖草胚芽鲜重的影响Fig.5 Effect of Cu2+ and Zn2+ on the germ fresh weight of Leymus secalinus
2.5 Cu2+、Zn2+对赖草胚根的影响
2.5.1Cu2+、Zn2+对赖草胚根长度的影响 由图6可知,Cu2+处理下,4种赖草的胚根长均随着Cu2+浓度的升高而降低;Cu2+浓度越大,胚根越短,说明Cu2+对4种赖草的胚根伸长具有抑制作用,且不同浓度Cu2+处理间4种赖草的胚根长存在显著差异(P< 0.05)。Zn2+处理下,LZ 1的胚根长随着Zn2+浓度的增加呈现递减的变化规律;其他3种赖草的胚根长整体上随着Zn2+浓度的增加呈先增大后减小的变化趋势, Zn2+浓度为20 mg·L-1时其胚根长度均达到试验组最大值,说明该浓度对胚根长度具有促进作用;当Zn2+浓度高于50 mg·L-1时则表现出明显的抑制作用。同时,同种金属相同浓度处理下,不同来源赖草的胚根长度对该浓度重金属胁迫表现出不同的响应,且存在显著性差异(P< 0.05)。
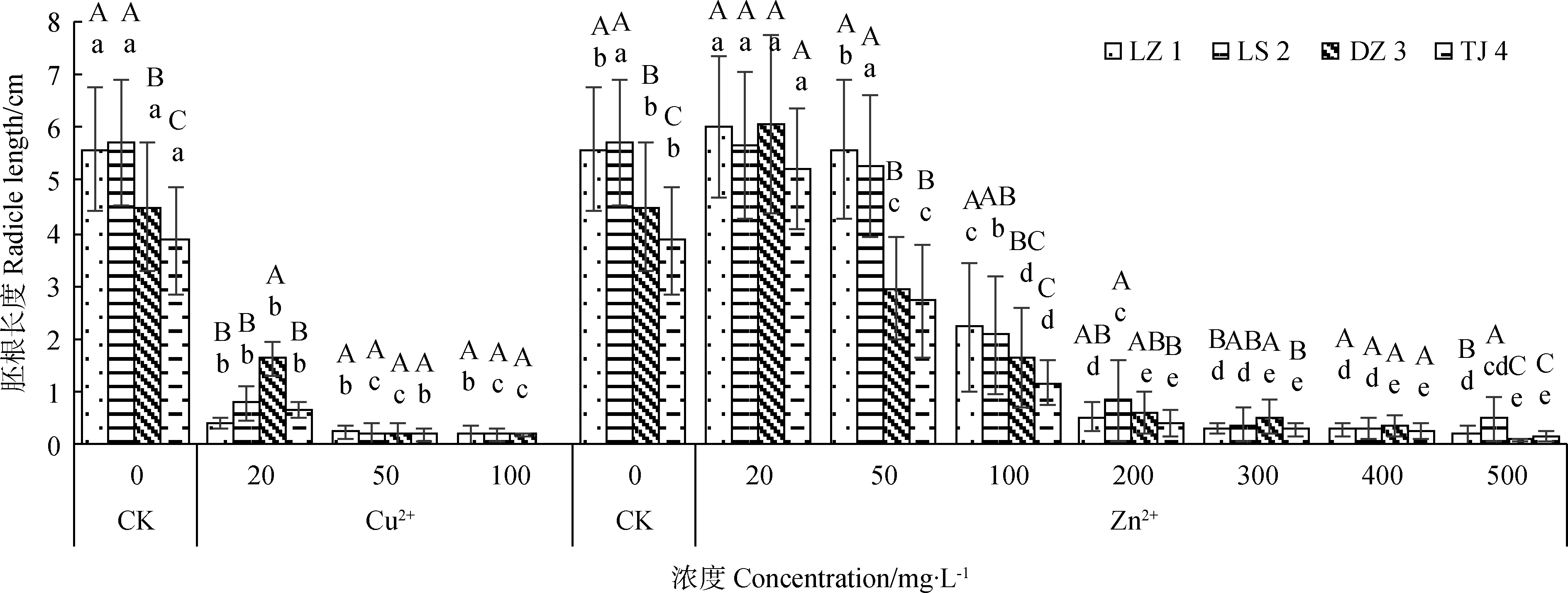
图6 Cu2+、Zn2+对赖草胚根长度的影响Fig.6 Effect of Cu2+ and Zn2+ on radicle length of Leymus secalinus
2.5.2Cu2+、Zn2+对赖草胚根鲜重的影响 由图7可知,Cu2+处理下,胚根鲜重随着Cu2+浓度的增大而降低,说明此时Cu2+对各赖草的胚根鲜重具有明显的抑制作用。Cu2+浓度由0 mg·L-1增加到20 mg·L-1时各赖草胚根鲜重迅速降低,Cu2+浓度为20~100 mg·L-1时,各赖草的胚根鲜重变化幅度则相对较小。不同浓度Zn2+处理下,各赖草的胚根鲜重整体上呈先增后减的变化趋势。Zn2+浓度为0~50 mg·L-1时各赖草胚根鲜重变化差异不显著,当Zn2+浓度为100 mg·L-1时,各赖草胚根鲜重量迅速降低,说明该浓度处理下各赖草的胚根鲜重具有明显的抑制作用。当Zn2+浓度超过100 mg·L-1之后,各赖草的胚根鲜重呈现缓慢的变化幅度。说明此浓度阶段的Zn2+对各赖草的胚根鲜重具有一定的抑制作用,但差异不显著。
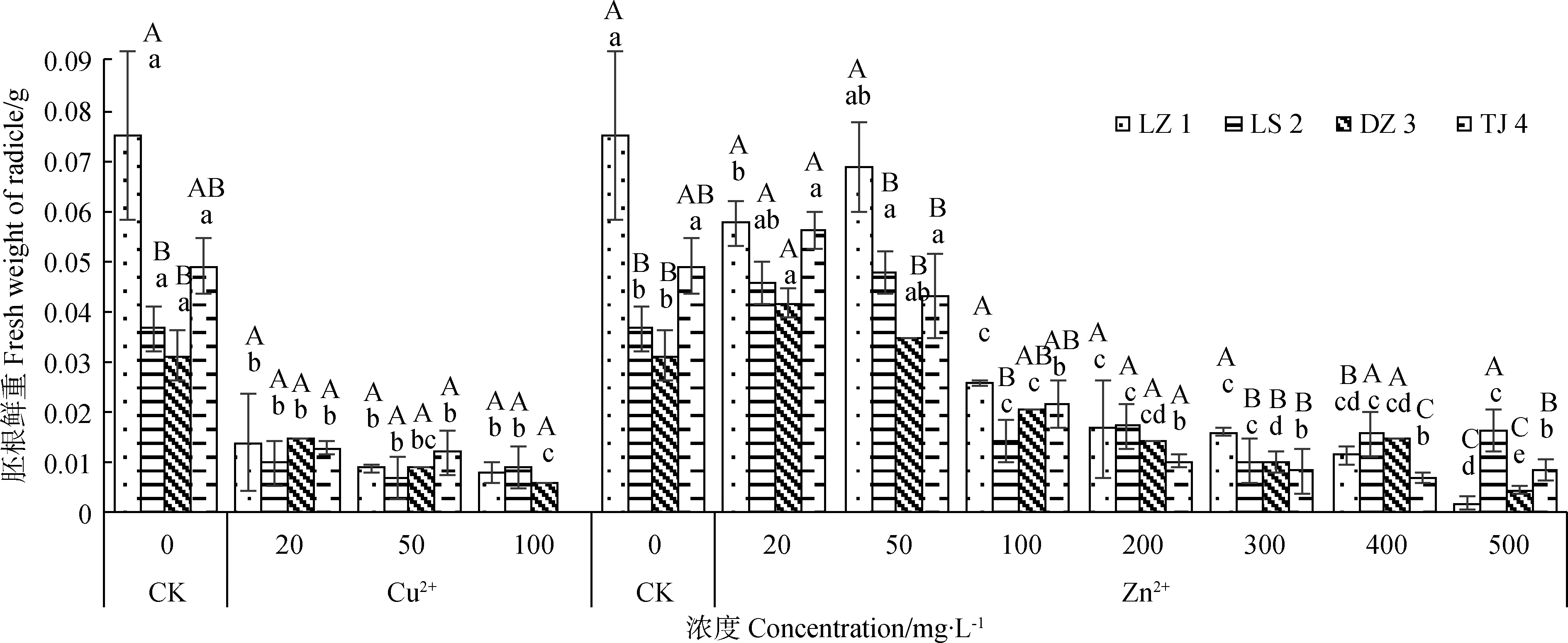
图7 Cu2+、Zn2+对赖草胚根鲜重的影响Fig.7 Effect of Cu2+ and Zn2+ on radicle fresh weight of Leymus secalinus
2.6 Cu2+、Zn2+对赖草根系耐受指数的影响
由图8可知,不同浓度Cu2+处理下,4个赖草的根系耐受指数随着Cu2+浓度增大而降低。不同浓度Zn2+处理下,LZ 1和LS 2的根系耐受指数均随着Zn2+浓度增大而降低,但DZ 3和TJ 4的耐受性指数先增大,当Zn2+浓度为20 mg·L-1时达到最高值,此后,两种赖草的根系耐受指数随着Zn2+浓度增大而降低,Zn2+浓度越大,根系耐受指数越低。由此可见,Cu2+和Zn2+对4个赖草根系耐受指数具有不同的影响,同种金属同浓度溶液对不同来源赖草的影响也不同。
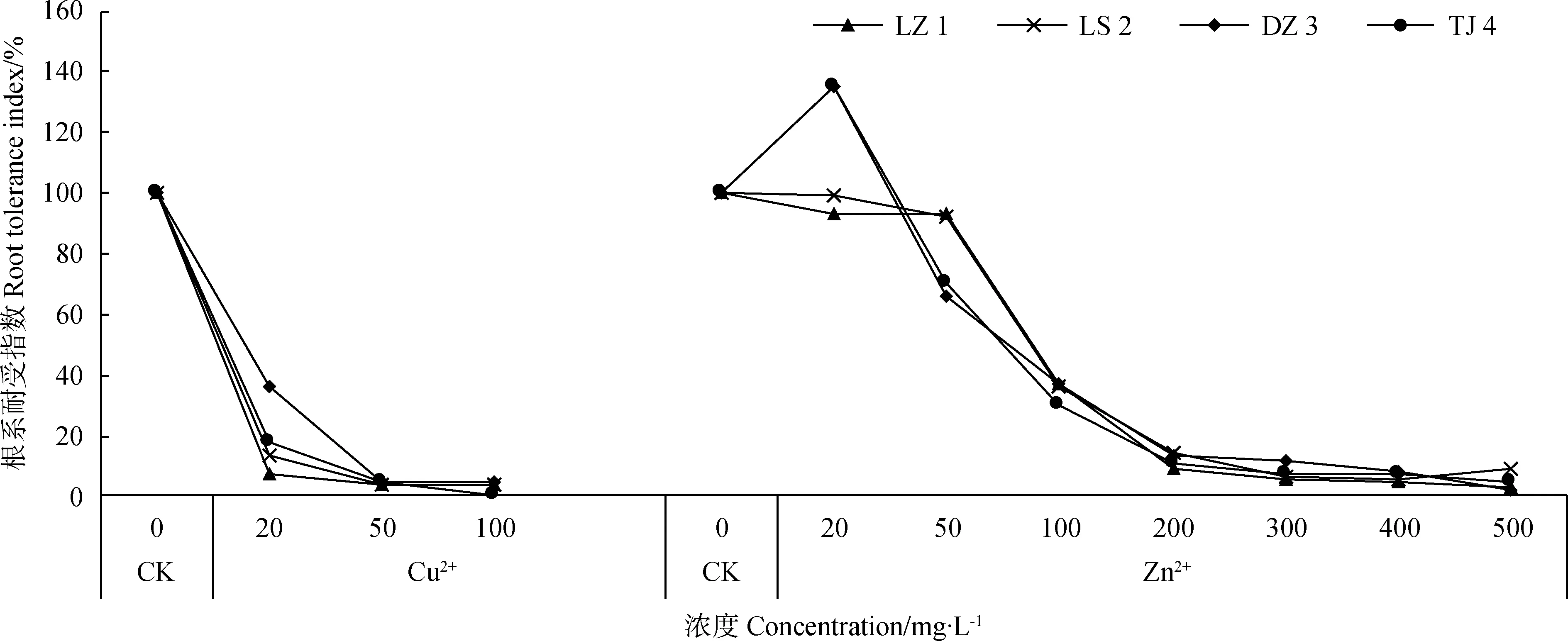
图8 Cu2+、Zn2+对赖草根系耐受指数的影响Fig.8 Effect of Cu2+ and Zn2+ on root tolerance index of Leymus secalinus
2.7 Cu2+、Zn2+对赖草种子活力指数的影响
由图9可知,不同浓度Cu2+处理下,DZ 3的种子活力指数先升高后降低,在Cu2+浓度为20 mg·L-1时达到最大值;其他3种赖草的种子活力指数均随着Cu2+浓度的增大而减小。LS 2的种子活力指数下降幅度最大,基本呈直线下降。不同浓度Zn2+处理下,4种赖草的种子活力指数随Zn2+浓度增加均呈先增大后减小的变化趋势,表明低浓度Zn2+提高种子活力指数,高浓度Zn2+降低种子活力指数。其中LZ 1和LS 2的活力指数在Zn2+浓度为50 mg·L-1时达到最大值,而DZ 3和TJ 4在Zn2+浓度为20 mg·L-1时就达到最大值,表明不同种赖草对Zn2+的耐受性不同。
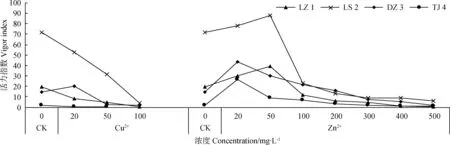
图9 Cu2+、Zn2+对赖草种子活力指数的影响Fig.9 Effect of Cu2+ and Zn2+ on seed vigor index of Leymus secalinus
2.8 Cu2+、Zn2+对赖草丙二醛(MDA)含量的影响
4种赖草的丙二醛(MDA)含量均随重金属离子(Cu2+、Zn2+)浓度的增大而增加,重金属离子浓度越大,MDA含量越高;表明重金属离子均对赖草胚芽细胞膜具有破坏作用,重金属浓度越大,破坏作用越强。在同浓度的重金属离子处理下,采自不同地区赖草的MDA含量不同,LZ 1的MDA含量变化幅度最大,Cu2+处理下增加了437.6 nmol·g-1,Zn2+处理下增加了597.9 nmol·g-1。LS 2的变化幅度最小,在Cu2+、Zn2+处理下分别增加了39.4 nmol·g-1和86.0 nmol·g-1。
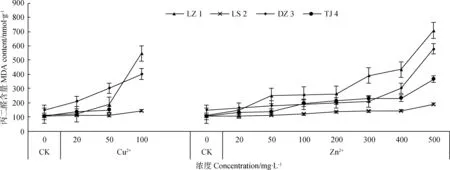
图10 Cu2+、Zn2+对赖草丙二醛含量的影响Fig.10 Effect of Cu2+ and Zn2+ on the MDA content of Leymus secalinus
3 讨论
发芽势、发芽率、发芽指数和种子活力不仅与牧草本身的生物学特性有关,而且与种子所处的生境具有密不可分的关系,是反映牧草出苗速率和出苗整齐度的重要指标[21-22]。赖草种子出苗率低或出苗不整齐,将导致作物大幅度减产或达不到重金属污染治理的效果[23]。研究过程中发现,Cu2+处理下,LS 2和DZ 3的发芽势、发芽率、发芽指数均呈先增大后减小的趋势,说明低浓度的Cu2+对这两种赖草种子的萌发具有促进作用;而高浓度Cu2+则表现出抑制作用,该结果与铜胁迫下黑麦草(Loliummultiflorum)[24]变化规律基本一致。LZ 1和TJ 4的发芽势、发芽率、发芽指数均随Cu2+浓度的增大而减小,说明本试验组浓度对这两种赖草只表现为抑制作用,未表现出低浓度促进萌发,高浓度抑制萌发的变化效应,该结果与铜胁迫下高羊茅(Festucaarundinacea)[24]和油菜(Brassicacampestris)[25]的变化规律一致。由此可以看出采自不同地域的野生赖草对Cu2+的耐性不同。
各赖草种子活力指数随Cu2+浓度的增大而减小,其主要原因是Cu2+处理下各赖草的胚芽、胚根鲜重均随Cu2+浓度的增大而减小,故此导致各赖草的种子活力指数呈由大到小的变化规律,这在本研究中得到验证,该结果与毛竹(Phyllostachysheterocycla)种子在Cu2+胁迫下的萌发特性相同[26]。胚芽、胚根的长度和鲜重及根系耐受指数的变化直接反映不同浓度重金属处理下赖草幼苗生长规律[27]。研究发现,Cu2+处理下各赖草的胚芽、胚根的长度和鲜重及根系耐受指数均随着Cu2+浓度的增大而减小,其中胚根长度和鲜重的减小幅度均大于胚芽,其原因可能是胚根是最早与Cu2+溶液直接接触的部位,受迫时间长,从而致使胚根受损[28];或者重金属离子抑制根尖细胞进行有丝分裂,致使根系生长发育受阻[29]。而胚芽未直接与重金属直接接触,故此对胚根的抑制作用大于胚芽,这与陈伟等[30]对Cu2+胁迫下4种草坪草种子萌发期胚根、胚芽变化的研究结果一致。
Zn2+处理下,各赖草种子萌发和幼苗生长的各项指标均表现“低浓度的促进作用和高浓度的抑制作用”[19]。表明低浓度Zn2+对各赖草萌发期的生长发育具有促进作用,但浓度较高时,表现出明显的抑制作用,其原因可能是高浓度的Zn2+对各赖草产生毒害作用,从而抑制了种子的萌发和幼苗的生长[31]。赖草种子发芽率的高低决定了其在重金属污染土壤恢复治理中的利用价值,研究发现,Zn2+浓度为20 mg·L-1时对各赖草的萌发和幼苗生长各指标均有促进作用,因此可采用该浓度的ZnSO4溶液浸种,以提高种子活力。研究中还发现,LS 2种子活力指数在Zn2+浓度为50 mg·L-1时达到试验组所有种子活力的最大值,且在Zn2+各浓度处理下活力指数也均较高,Cu2+处理下也是LS 2的种子活力最强,因此可以对LS 2进行驯化选育,以便作为Zn2+污染土壤恢复治理的选择材料。
MDA是植物膜脂过氧化的终产物之一,其在植物体内的积累会对膜和细胞进一步造成伤害,MDA含量的高低可以反映该植物在不同浓度重金属离子胁迫下受到的伤害程度[32]。为了探究重金属离子是否会对植物产生毒害作用,本试验分别测定了重金属离子(Cu2+、Zn2+)处理下各赖草胚芽中的MDA含量。研究发现各赖草胚芽中MDA含量随重金属离子浓度的升高而增大,低浓度时MDA含量变化差异不显著,而高浓度时表现出显著性差异(P<0.05)。由此可见,无论重金属离子浓度的高低,都会对赖草产生一定的不利影响,重金属浓度越大,破坏作用越强。
本次试验还发现采自西藏4个不同地域野生赖草在同种金属同种浓度处理下的表现性状也有所差异。其原因可能是野生赖草在长期适应生境的过程中产生变异或者由于生境的不同从而导致遗产背景的不同[33],故同种金属同浓度处理下采集不同地域的赖草各指标间存在差异性。
据报道,脱落酸是限制赖草属牧草萌发和生长的一种重要激素[34],研究发现,相同处理下,LS2的发芽率高于TJ4,其原因可能是LS2采集于拉萨河边的沙地,水热条件充足,在长期适应该环境的过程中,由于逆境胁迫小,因此脱落酸含量减少,发芽率高。而TJ 4采集于塔吉乡田间地埂上,人为和各种外界影响因素较大,因此赖草通过增加脱落酸含量,降低发芽率来适应不利的生境。但易津等研究证明,随着赖草属种子贮存年限的增加,脱落酸含量逐渐降低,种子发芽率随之逐渐提高[35],试验材料LS 2和DZ 3是2011年收获的种子,LZ 1是2012年采集的,TJ 4是2013年采集的,这也符合上述理论的表述,至于具体原因尚待进一步研究。研究还发现低浓度的重金属离子对赖草的各项指标均有促进作用,其原因可能是,Cu2+和Zn2+均是植物生长的必须元素,低浓度的重金属给赖草的萌发提供了一个有利环境,抑制了种子萌发和幼苗生长过程中脱落酸的合成,促进了赖草种子萌发和幼苗生长。
4 结论
重金属Zn2+处理下,各赖草种子萌发和幼苗生长的各项指标均呈先增大后减小的趋势,表现出低浓度促进、高浓度抑制效应。重金属Cu2+处理下,LS 2和DZ 3的发芽势、发芽率、发芽指数也表现出相同的变化规律;但LZ 1和TJ 4的发芽势、发芽率、发芽指数及4个野生赖草的幼苗生长指标却均随Cu2+浓度的增大而减小。重金属离子(Cu2+、Zn2+)对胚根的影响大于对胚芽的影响。不同地方的赖草对不同重金属离子的抗性也不同,对采自于西藏4个不同地方的赖草种子活力指数和MDA含量进行比较,得出LS 2的发芽指标高于其他3个赖草,且对重金属离子(Cu2+、Zn2+)均具有较强的抗性,可以将其进行选育,以期培育出适合于青藏高原重金属污染土壤生态恢复的赖草品种。
猜你喜欢
今日农业(2022年6期)2022-07-05
山西农业科学(2021年8期)2021-08-17
青年文学家(2021年4期)2021-03-18
农民致富之友(2020年19期)2020-07-18
科学导报(2020年8期)2020-03-12
山西农业科学(2020年1期)2020-02-27
中国食品学报(2020年2期)2020-01-17
新农民(2019年21期)2019-11-09
家庭百事通·健康一点通(2019年11期)2019-09-26
潍坊学院学报(2016年2期)2016-12-01
