
紫花苜蓿蓝光受体CRY2B过表达载体构建及转化
2017-09-13 05:05樊文娜李润林王占彬杜红旗赵凌平王成章
草地学报 2017年5期
樊文娜, 李润林, 王占彬, 杜红旗, 赵凌平, 王成章*
(1. 河南科技大学动物科技学院, 河南 洛阳 471003;2. 河南农业大学牧医工程学院, 河南 郑州 450002)
苜蓿(Medicagosativa)的秋眠性(Fall Dormancy)是苜蓿在秋季因日照长度变短和气温下降所表现出的一种适应性生长特性[1-2],是选择栽培苜蓿品种时考虑的首要指标。从本质上讲,秋眠性是对日照长度的反应,短日照是其主要的影响因子[3]。苜蓿作为长日照植物,晚春及夏季长日照条件下有利于其生长发育,秋冬季短日照则休眠,说明苜蓿秋眠存在光周期效应[4]。
目前,研究已发现的前3种光信号接受的光受体[5-8],即红光和远红光的受体——光敏色素(Phytochrome )、蓝光受体和紫外光受体(UV-receptor)。隐花色素(Cryptochrome)在植物中是很重要的感受光受体,是光形态建成反应的必需元素,但在苜蓿的秋眠中发挥的作用机理目前还未知。高等植物受蓝光调节的反应主要包括:向光性、抑制茎芽的生长、叶绿体的转移、刺激气孔张开和关闭,从而调节相关基因的表达。蓝光光受体——隐花色素可以调控植物开花时间以及昼夜节律循环[9],CRY2B(Cryptochrome2B)是主要接受和传递光信息的蓝光受体,并且紫花苜蓿转录组高通量测序结果差异基因的统计也证明,调控苜蓿秋眠的蓝光受体CRY2B的Log2(Fold Change)[10-11]是最大的,这证明CRY2B作为重要的光受体在调控苜蓿的秋眠中发挥着至关重要的作用。
本研究拟构建紫花苜蓿蓝光受体CRY2B过表达载体,过表达载体的构建本质上是基因重组的过程。构建紫花苜蓿蓝光受体CRY2B过表达载体是获得紫花苜蓿CRY2B转基因植株的关键步骤和基础。
1 材料与方法
1.1 试验材料
1.1.1植物材料 供试的紫花苜蓿(Medicago
sativaL.)品种为一级品种‘驯鹿’,由河南农业大学牧医工程学院草业科学实验室培养提供,采集叶片后迅速放入液氮速冻后-80℃保存。
1.1.2菌株 大肠杆菌(Escherichia.coli)DH5α,农杆菌菌株GV3301,均由河南农业大学草业科学实验室提供。
1.2 载体
克隆载体pMDl8-T(Takara);
植物表达载体pCAMBIA3301含有CaM 35 S启动子和Nos polyA终止子(图1)。

图1 pCAMBIA3301结构图Fig.1 Diagram of Pcambia3301
1.3 试验方法
1.3.1CRY2B基因引物设计 引物设计前分析序列(紫花苜蓿基因组测序尚未完成,结合紫花苜蓿转录组测序信息,以模式植物蒺藜苜蓿CRY2B基因为模板 ,NCBI登录ID:XM-003589988.1),基因序列编码区696-2498(即CRY2B完整的开放阅读框),设计引物扩增出来的目的片段要包含此阅读框,如下:AATCTGAAAATAGGTATGAATAGGACCATAGTT-TGGTTTAGGAGGGACCTAA-GAATTGAGGACAA………TACCATAACAATGTTCCTCT-CCTCTTCTACTAATATAAAGTT-GTCATGGTAAGAACATAG(加粗部分划线为CRY2B起始密码子和终止密码子,目的基因过表达要求引物设计的长度从起始密码子和终止密码子,含完整的开放阅读框)。
利用Premier 5设计引物,选择过表达载体上NcolI、PmlI为酶切位点(同时对植物表达载体pCAMBIA3301和目的基因CRY2B进行双酶切),在设计好的CRY2B引物(696-2553)上加入酶切位点(划线部分为限制性酶切位点),为保证酶切质量,酶切位点前面为保护碱基(表1) ,并送生工生物(上海)股份有限公司。

表1 基因引物序列Table 1 The primers of CRY2B gene
1.3.2CRY2B基因克隆与测序 PCR产物经1.5%琼脂糖凝胶电泳检测后回收目的片段,进行PMD18-T克隆转化,菌液PCR检测为阳性后送上海生工测序。测序结果在http://blast.ncbi.nlm.nih.gov/Blast.cgi进行比对,CRY2B克隆成功。用质粒提取试剂盒提取质粒,保存并命名为PMD18-T-CRY2B。
1.3.3纯化回收的产物与PMD-18T载体的连接 采用北京百泰克生物技术有限公司琼脂糖凝胶DNA回收试剂盒回收,严格按照回收试剂盒的操作步骤进行,回收到的产物置于-20℃冰箱中保存备用。
纯化回收产物CRY2BPMD-18T载体连接前,使用BspHI/PmlI内切酶,分别双酶切CRY2B目的片段和植物表达载体pCAMBIA3301,为提高CRY2B和pCAMBIA3301载体两个目的片段的酶切效率和准确性,先将目的片段与PMD-18T载体连接,增加CRY2B目的片段的拷贝数。
DH5α感受态细胞转化,用于扩繁、培养,进行PCR鉴定。
1.3.4提取阳性克隆DNA质粒 利用百泰克公司小量质粒提取试剂盒提取,严格按照提取质粒试剂盒的操作步骤进行。
1.3.5含有T载体的CRY2B片段质粒与pCAMBIA3301质粒双酶切 在冰上进行操作,反应液混匀后37℃酶切反应1 h。
1%的琼脂糖凝胶电泳观测酶切效果,对酶切正确的目的片段进行切胶纯化回收,以备连接反应。
1.3.6连接反应 将纯化回收酶切后的CRY2B目的片段连接到酶切后的pCAMBIA3301上,连接反应目的片段与载体的浓度摩尔比例为1:6~10。

表2 双酶切反应体系Table 2 The reaction system of restriction endonuclease
1.3.7连接产物转化及鉴定 pCAMBIA3301载体与CRY2B目的片段连接产物转化操作同上述DH5α感受态细胞转化步骤,由于载体pCAMBIA3301含有Kan、Amp的特性,所以配制含有Kan、Amp特性的LB培养基,转化后对含有目的片段菌液进行PCR检测。PCR检测为阳性克隆的菌液进行测序并提取质粒,长期保存,然后进行酶切检测。
1.3.8酶切检测 由于CRY2B基因上游添加的酶切位点是NcolI的同尾酶BspHI,因此重新整合的重组质粒不能用原来的酶切位点NcolI或PmlI去进行酶切鉴定,利用generunner分析可知,CRY2B基因片段上含有和pCAMBIA3301载体上相同的酶切位点EcoR1,此酶切位点不在pCAMBIA3301载体的表达区。利用EcorI进行酶切,若琼脂糖凝胶电泳得到10 000 bp和2 000 bp大小的两个片段(可根据pCAMBIA3301载体序列酶切位点的分析和CRY2B基因扩增长度判断),则说明目的片段pCAMBIA3301-CRY2B重组质粒构建成功,如果没有得到两个片段,则质粒重组不成功。
1.3.9农杆菌介导pCAMBIA3301-CRY2B转化苜蓿植株 质粒为上述试验构建的MBIA3301-CRY2B过表达载体,首先制备农杆菌GV3101感受态细胞,然后选择粒大饱满的‘驯鹿’紫花苜蓿种子,使用清水冲洗30 min,然后75% 的酒精浸泡3 min进行消毒,弃溶液,然后用0.1%HgCl2(氯化汞)溶液灭菌15 min,再用无菌水清洗3~5次,将种子置于多层无菌滤纸上直至吸去多余水分,用已灭菌的镊子将种子接种到MS培养基上(30 g·L-1蔗糖和2.5 g·L-1Phytagel)。将接种的培养基放入光照培养箱里,在白昼光周期为16 h/8 h,白昼温度为22/20℃的条件下培养,采用直接分化芽再生途径的方法进行农杆菌介导pCAMBIA3301-CRY2B重组载体转化紫花苜蓿[12-16]。
1.3.9.1 农杆菌的侵染
(1)在含重组质粒pCAMBIA3301-CRY2B农杆菌的YEB(50 mg·L-1kan+100 mg·L-1rif)固体培养基上划线培养,放入28℃培养箱培养48 h左右。
(2) 2 d后,用牙签挑取单菌落,接种于含有50 mg·L-1kan和100 mg·L-1rif的20 mL YEB液体培养基中,28℃ 200~220 rpm震荡培养16~24 h, OD600=0.5~0.7,用于外植体的侵染。
(3)将切割好的子叶放入准备好的农杆菌菌液中侵染60 min(隔10 min轻微晃动1次),使农杆菌能够充分接触子叶节的伤口。接着将侵染过的子叶转移到干燥的无菌滤纸上,吸去表面多余的菌液,然后接种到共培养培养基上(光照培养箱24℃黑暗培养,3~4 d)。
1.3.9.2 芽诱导和筛选
共培养后的子叶转入芽诱导培养基中,注意把生长点插入培养基以保证子叶的生长方向,不定芽直立向上生长,并且子叶节处的伤口能够接触到培养基[17-20]。培养条件为光周期:16 h/8 h,温度:白昼25℃/22℃。转化成功的植株会在子叶节的伤口处直接分化出不定芽,且不定芽出现后会迅速生长。未转化成功的外植体会逐渐变黄、发褐甚至死亡。
1.3.9.3 诱导生根
选择芽诱导长势较好且已长有小根须的样本,在无菌条件下转入含有生根培养基(1/2MS+Yeastextract +carb)的三角瓶(瓶口较大,太小容易污染)中,用封口膜将锥形瓶口封紧,放入光照培养箱,设置培养条件,光照时间和温度:16 h(25℃)/黑暗时间和温度8 h(22℃)。诱导根生长30天左右,幼苗主根茁壮,接近于正常的植株。
1.3.9.4 炼苗移栽
当苜蓿转基因植株根部产生多条粗而壮的根须时,可以进行炼苗,即对三角瓶内的转基因幼嫩植株进行驯化,以适应外界生长条件[21]。打开三角瓶瓶口,加入一定量的灭菌去离子水(避免出现干燥应激)。在光照培养箱中生长4~6 d,将幼苗从三角瓶中取出,冲洗根部的培养基(一定要冲洗干净,避免病菌滋生使根部坏死)。之后将转基因苜蓿植株移栽于混有营养土/蛭石(1:1)的小花盆中。当苜蓿
植株长出健壮茎叶时,即可进行转基因阳性苗的鉴定。
2 结果与分析
2.1 CRY2B克隆PCR扩增效果
根据克隆的基因以及所需的酶切位点设计CRY2B引物,进行PCR扩增,用1%的琼脂糖凝胶电泳检测。结果如图2所示,目的片段条带清晰明亮,可进行目的片段纯化回收。

图2 CRY2B PCR 扩增Fig.2 Electrophoresis of the product of CRY2B PCR amplificationM: DNA分子量标记;1-2: CRY2BM: DNA ladder; 1-2: CRY2B
2.2 pCAMBIA3301载体与CRY2B目的片段连接的菌液PCR检测
将纯化回收酶切后的CRY2B目的片段连接到酶切后的pCAMBIA3301连接产物,转化到DH5α大肠杆菌过夜培养。根据克隆的CRY2B基因以及所需的酶切位点所设计的引物,以菌液为模板,进行PCR扩增检测。扩增出如图3清晰明亮的目的片段,说明目的片段已经成功连接到pCAMBIA3301载体上,可以进行测序鉴定与酶切反应。
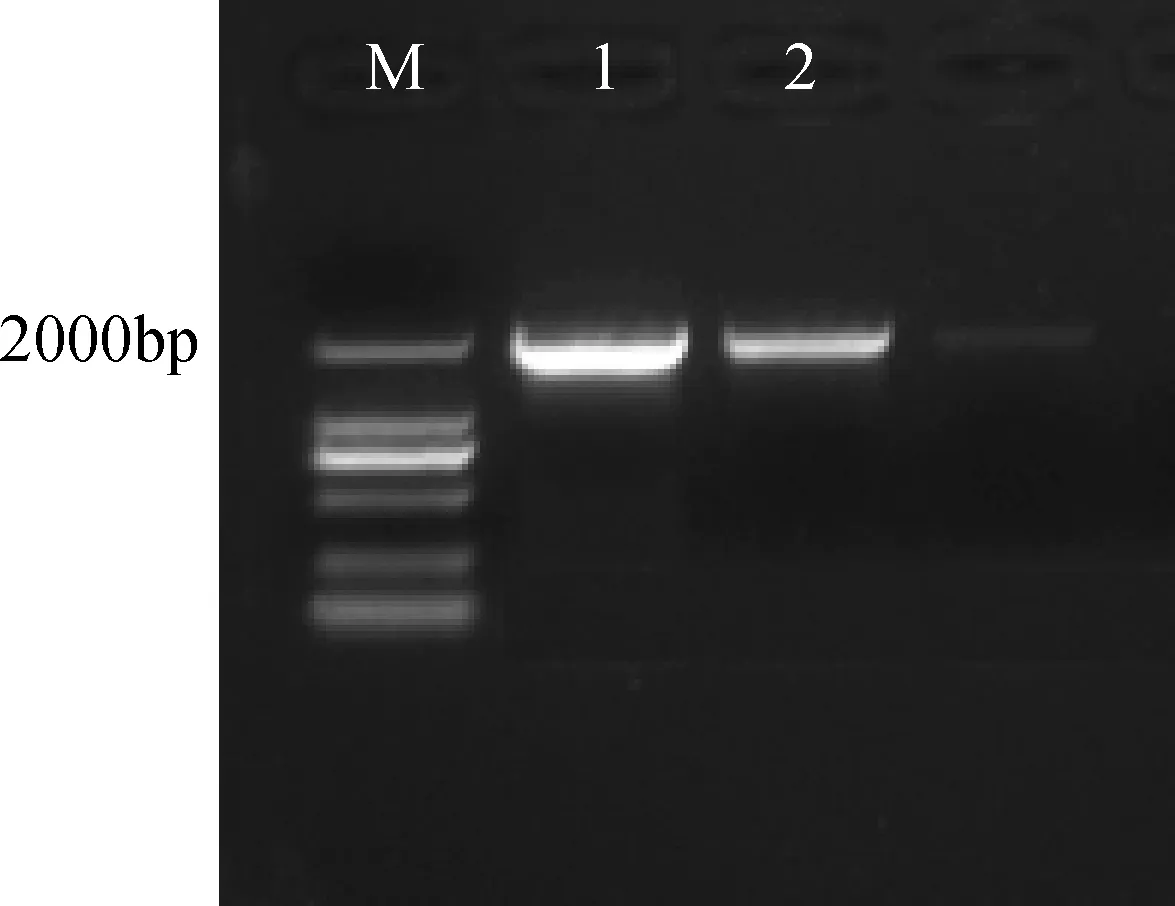
图3 CRY2B 菌液PCR 扩增Fig.3 CRY2B bacteria PCR amplificationM: DNA分子量标记;1-2: CRY2BM: DNA ladder; 1-2: CRY2B
2.3 pCAMBIA3301载体与CRY2B目的片段连接产物的酶切效果与分析
pCAMBIA3301载体与CRY2B目的片段连接产物转化操作同上述DH5α感受态细胞转化,由于载体pCAMBIA3301含有Kan、Amp的特性,所以配制含有Kan、Amp特性的LB培养基,转化后对含有目的片段的菌液进行PCR检测,检测为阳性克隆的菌液,然后进行EcoRI酶切鉴定检测。如图4酶切出现一个10 000 bp大小和一个2 000 bp大小的片段条带。
对酶切鉴定出现正确片段的菌液送往生工生物工程(上海)股份有限公司测序,测序结果和目的片段序列一致(图5),表明pCAMBIA3301-CRY2B重组质粒构建成功,保存pCAMBIA3301-CRY2B阳性克隆的菌液或提取阳性克隆菌液的质粒保存。

图4 pCAMBIA3301-CRY2B质粒酶切鉴定Fig.4 pCAMBIA3301-CRY2B plasmid restrictionenzyme digestion analysis1: pCAMBIA3301-CRY2B 酶切;M: DNA 分子量标记;1 pCAMBIA3301-CRY2B restriction enzymeanalysis; M: DNA ladder

图5 克隆片段与CRY2B基因片段序列比对Fig.5 Sequence alignment between cloning sequence and CRY2B fragment
2.4 重组质粒PCR检测
将重组质粒pCAMBIA3301-CRY2B导入农杆菌GV3101感受态细胞,在含有50 mg·L-1Kan + 50 mg·L-1rif的YEB培养基上涂板,28℃培养48小时。以菌液为模板,进行PCR扩增,阳性菌落的PCR琼脂糖凝胶电泳图如图6所示。挑选的单菌落都为阳性菌落,表明植物表达载体pCAMBIA3301-CRY2B已转化进农杆菌GV3101中,可用于后期紫花苜蓿的遗传转化。
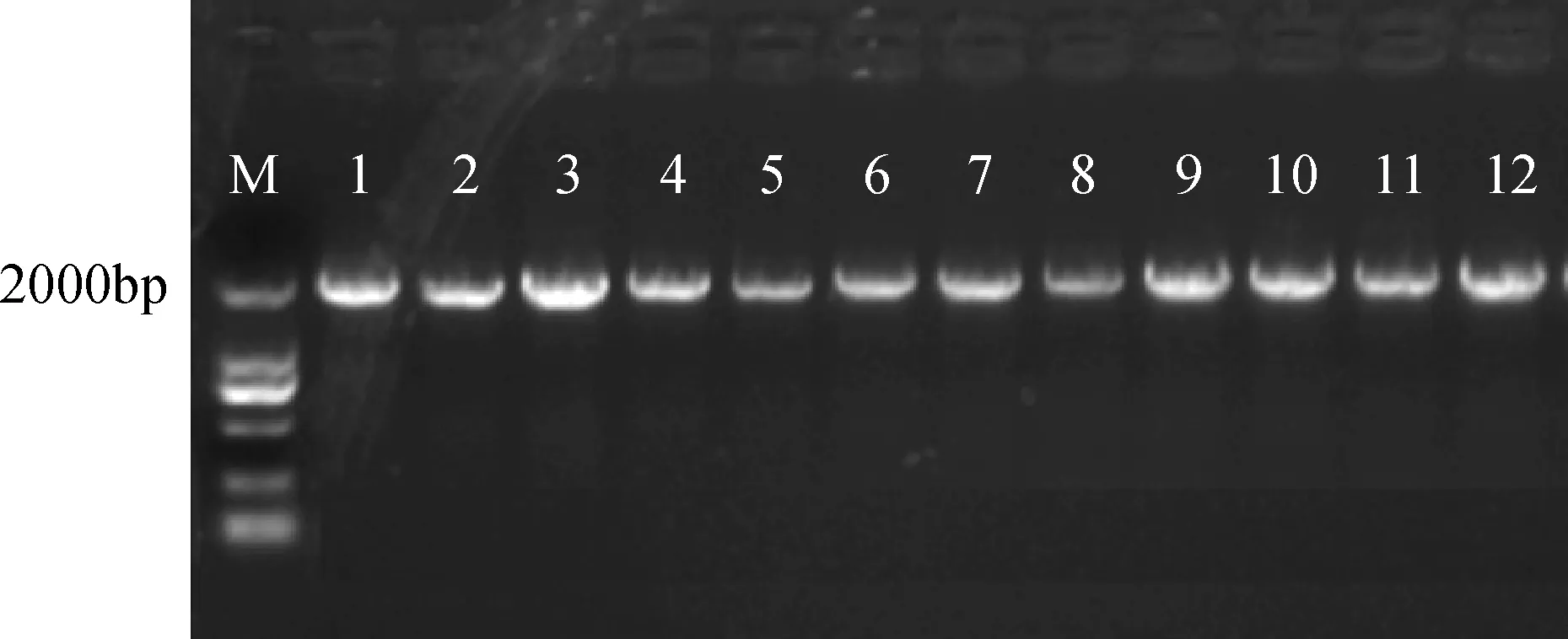
图6 pCAMBIA3301-CRY2B质粒PCR扩增Fig.6 pCAMBIA3301-CRY2B plasmid PCR amplification1-12 CAMBIA3301-CRY2B PCR扩增;M: DNA 分子量标记;1-12 CAMBIA3301-CRY2B PCR amplification; M: DNA ladder
2.5 酶切鉴定重组质粒
利用上述菌液提取质粒进行酶切鉴定,选用pCAMBIA3301载体和CRY2B共有的酶切位点EcoRI,酶切鉴定结果如图7所示,酶切出两条正确的基因片段。
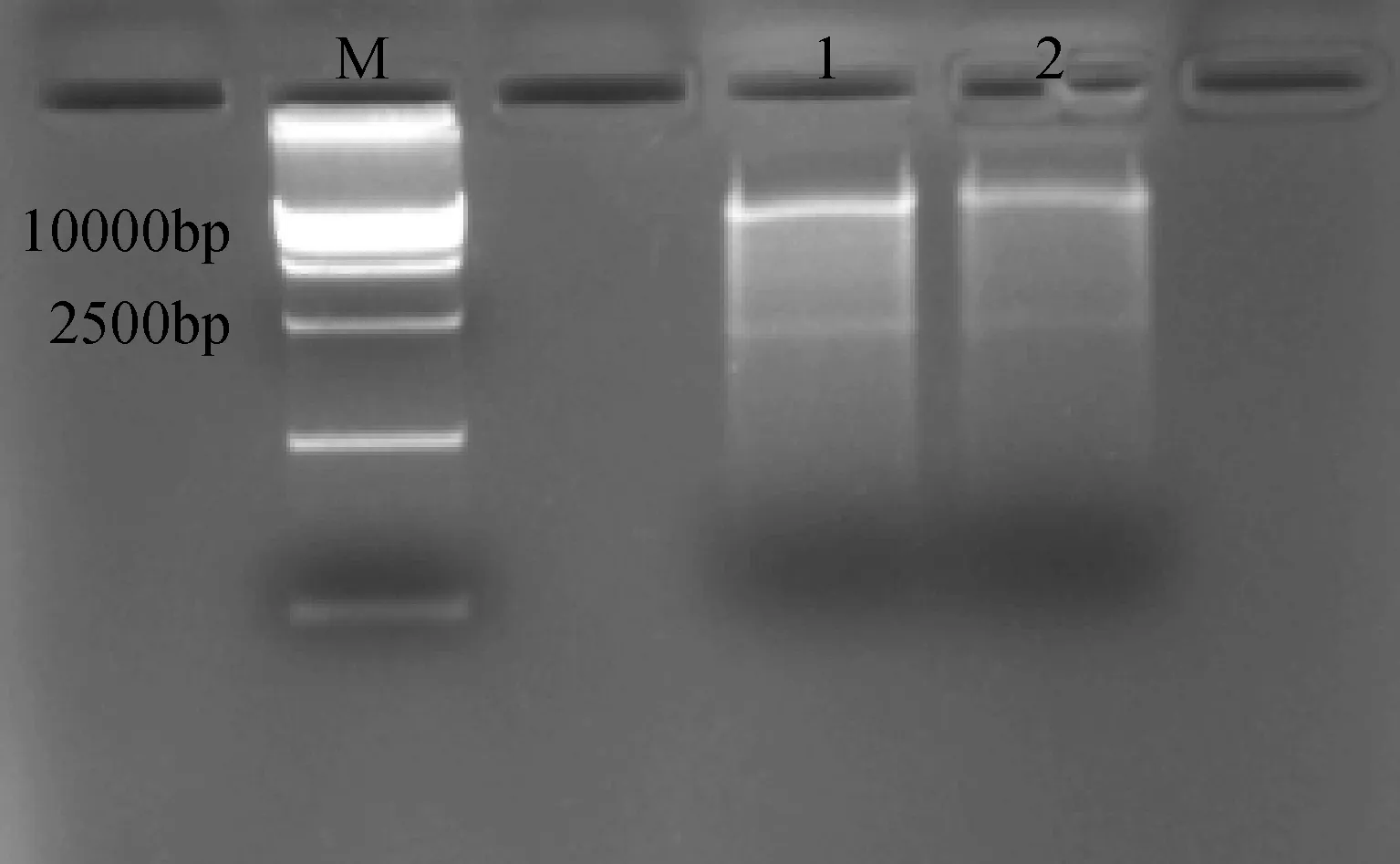
图7 pCAMBIA3301-CRY2B农杆菌质粒酶切鉴定Fig.7 pCAMBIA3301-CRY2B Agrobacteriumplasmid restriction enzyme analysis1-2: pCAMBIA3301-CRY2B 酶切;M: DNA 分子量标记1-2:pCAMBIA3301-CRY2B restriction enzyme analysis; M: DNA ladder
2.6 pCAMBIA3301-CRY2B转基因植株诱导芽的生长
以子叶节为外植体,适宜的Kan抗性筛选浓度为100 mg·L-1,适宜的子叶节芽诱导培养基为MS+1.0 mg·L-16-BA+0.1 mg·L-1NAA+100 mg·L-1Kan+400 mg·L-1Carb,pCAMBIA3301-CRY2B转基因无菌苗子叶经侵染之后,共培养4天,在诱导芽培养基上进行芽诱导,14~18 d左右子叶长出新芽(图8)。

图8 诱导芽的生长Fig.8 The growth of induced shoots
2.7 pCAMBIA3301-CRY2B转基因植株诱导根的生长
适宜的子叶节根诱导培养基为:1/2MS+0.2 g·L-1YE+100 mg·L-1Kan+400 mg·L-1Carb。pCAMBIA3301-CRY2B长出新芽的个体移入生根培养基中生长30~40 d,无菌苗苜蓿生根情况如图9所示。

图9 pCAMBIA3301-CRY2B转基因植株诱导根的生长Fig.9 The growth of induced roots with transgenic-pCAMBIA3301-CRY2B
2.8 炼苗与再生植株移栽
当再生的芽生长健壮达到约4 cm时,即可转移到生根培养基上进行生根诱导。待根部足够发达,出现多条根时,开始炼苗。约3~5 d后,小心取出小植株,把根部冲洗干净,种植到小花盆中。炼苗初期采用透明薄膜罩盆保持较高的湿度,待植株生长稳定后,即可摘除,仍保持湿润环境,保证幼嫩转基因植株较大的成活率(图10)。

图10 炼苗和再生植株移栽Fig.10 Plantlets exercising and transplanting of regenerated plantlets
3 讨论与结论
对拟南芥(Arabidopsisthaliana)的研究发现,隐花色素和光敏色素具有拮抗和互补的作用[22],研究表明,光受体基因通过光周期参与了对苜蓿秋眠的调控[23-27],在对PHYA、PHYB沉默的转基因植株研究中发现,CRY2B的表达和PHYB的表达近乎完全一致[23]。秋眠型紫花苜蓿CRY2B在夏季高温休眠条件下基因表达量没有发生变化,但秋眠条件下呈显著增长[23],因此在对紫花苜蓿秋眠调控机制的研究中,CRY2B基因功能的研究至关重要。
本试验CRY2B过表达载体构建成功后直接转化紫花苜蓿。通过在CRY2B基因的引物两端加入NcolI的同尾酶BspHI(使用同尾酶可产生相同的粘性末端)和PmlI位点,以提取的苜蓿RNA为模板进行PCR扩增得到CRY2B基因片段。选择过表达载体上NcolI、PmlI作为双酶切的酶切位点(载体上没有BspHI酶切位点),NcolI酶切(A/CATGT),为了使氨基酸正确翻译,CRY2B引物添加NcolI酶切位点时要使用同尾酶BspHI(T/CATGA)。然后将目的片段CRY2B和过表达载体pCAMBIA3301经过双酶切,使其产生相同的黏性末端。将切下的目的基因CRY2B的片段插入质粒的切口处,首先碱基互补配对结合,两个黏性末端吻合在一起,利用NEB公司T4连接酶分别将酶切后基因片段与酶切后pCAMBIA3301质粒整合到一起,构建pCAMBIA3301-CRY2B重组质粒,然后经过PCR检测、碱基序列测序和酶切鉴定确定成功构建了pCAMBIA3301-CRY2B过表达载体,植物表达载体pCAMBIA3301-CRY2B已转化进农杆菌GV3101中,可用于后续转基因紫花苜蓿的遗传转化。
近年来,紫花苜蓿载体构建并对烟草进行转化研究较多[28-31],但转化方式不尽相同。本试验采用直接分化芽再生途径转化紫花苜蓿,以萌发早期幼苗的子叶节为外植体,经农杆菌侵染后无需组织培养,外植体细胞不经过脱分化形成愈伤组织的阶段,而是直接分化出不定芽,进而生长发育为完整植株。该途径的优点是:植株再生周期短,遗传变异较小,转入的外源基因遗传稳定性高。郑宝仁等[32]以子叶节为外植体,在MS培养基上添加不同的激素组合直接进行不定芽的诱导,最终得到离体再生植株。Kumar等[33]研究发现对茎尖外植体可以直接进行不定芽的诱导(最多时一个外植体可以诱导出35个不定芽),然后再加入适当比例的NAA和活性炭即可诱导生根进而得到完整植株,并且该流程能够适用于5个印度苜蓿品种。本试验以紫花苜蓿子叶节为外植体,利用农杆菌介导pCAMBIA3301-CRY2B成功转化苜蓿植株,为进一步研究CRY2B调控苜蓿秋眠性分子机制奠定基础。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
现代畜牧科技(2021年9期)2021-10-13
现代畜牧科技(2021年4期)2021-07-21
畜牧兽医科技信息(2021年9期)2021-03-05
疯狂英语·新悦读(2020年7期)2020-07-30
癌变·畸变·突变(2020年1期)2020-02-12
草业科学(2018年12期)2018-12-27
中国三峡(2017年4期)2017-06-06
为了孩子(3~7岁)(2016年6期)2016-05-14
中国兽药杂志(2014年3期)2014-11-29
