
惠阳胡须鸡IGF-Ⅰ和GHRL基因多态性研究
2017-09-13 09:20:29李红伟钟淡龙吴晓燕邹志冠
河南农业科学 2017年9期
李红伟,钟淡龙,陈 圆,吴晓燕,邹志冠
(1.惠州学院 生命科学系,广东 惠州 516007; 2.广东金种农牧科技股份有限公司,广东 惠州 516021)
惠阳胡须鸡IGF-Ⅰ和GHRL基因多态性研究
李红伟1,钟淡龙1,陈 圆1,吴晓燕1,邹志冠2
(1.惠州学院 生命科学系,广东 惠州 516007; 2.广东金种农牧科技股份有限公司,广东 惠州 516021)
以IGF-Ⅰ和GHRL基因为惠阳胡须鸡生长性状的候选基因,通过测序、酶切等方法对93日龄惠阳胡须鸡IGF-Ⅰ和GHRL基因多态性进行分析,并用GLM模型分析基因型与体质量的关联性。结果表明: IGF-Ⅰ基因在第181位发生碱基突变,A突变为G,属于同义突变,BsmⅠ酶切分析发现,IGF-Ⅰ基因只有1种基因型,表明该位点在胡须鸡保种群中已经纯合;GHRL基因在第124位发生碱基突变,T突变为G,改变了酶切位点,通过pflMⅠ酶切分析发现,GHRL基因有3种基因型,即AA型、AB型和BB型。统计结果表明,93日龄惠阳胡须鸡3种基因型个体间体质量无显著差异。
惠阳胡须鸡; IGF-Ⅰ基因; GHRL基因; 基因多态性
惠阳胡须鸡以特有的优良肉质和三黄胡须鸡的外貌特征而驰名,在育种、生产和外贸活鸡市场上都具有较高的经济价值,但存在生长速度慢等缺点。寻找与惠阳胡须鸡生长性状相关的候选基因,实施分子标记辅助选择育种十分必要。
IGF-Ⅰ全称胰岛素样生长因子,对动物生长发育有重要的调控作用,是调节动物生命活动的重要多肽生长因子之一[1]。Florini等[2]研究发现,IGF-Ⅰ基因通过影响肌纤维的分化、增殖和蛋白质沉积,间接影响肌肉的生长速度。赵秀华等[3]研究发现,IGF-Ⅰ基因部分单核苷酸多态性与京海黄鸡生长性状相关,认为可应用于标记辅助选择,加快育种进程。而刘大林等[4]研究认为,IGF-Ⅰ基因可能控制着鸡的生长性状和屠体性状或与控制生长性状、屠体性状的主效基因连锁,可以把IGF-Ⅰ基因作为鸡生长、屠体性状的辅助标记基因。生长素(ghrelin,GHRL)是促进生长激素(growthhormone,GH)分泌的关键因子,可以调节机体生长发育、增加食欲、调节代谢及能量平衡、促进胃酸分泌等[5-10]。动物饥饿时,血液中的GHRL量增加,在促进摄食的同时减少了体内脂肪的消耗,引起动物体质量增加[11]。巢湖鸭GHRL基因多态性与胴体质量、半净膛质量和全净膛质量等屠体性状有显著相关性[12]。何丹林等[13]研究表明,鸡GHRL基因C2100T位点多态性与鸡的部分生长性状存在显著相关性。Fang等[14]研究表明,GHRL基因外显子1处的8bp插入缺失多态性与鸡生长性状和屠体性状显著相关,而GHRL基因部分单核苷酸多态性与鸡腹脂质量、腿肌粗蛋白含量显著相关[15]。
综上可知,IGF-Ⅰ和GHRL基因与鸡生长性状紧密相关。因此,以IGF-Ⅰ和GHRL基因为惠阳胡须鸡生长性状的候选基因,分析IGF-Ⅰ和GHRL基因多态性及其与体质量的关系,以期为今后惠阳胡须鸡育种中生长性状的分子标记辅助选择提供理论依据。
1 材料和方法
1.1材料
100份93日龄的惠阳胡须鸡血样采自广东金种农牧科技股份有限公司的惠阳胡须鸡保种场。将血样放入含肝素钠的采血管,-20 ℃保存。
1.2引物设计与PCR扩增
IGF-Ⅰ基因引物设计参照文献[3],GHRL基因引物设计参照文献[15]。引物由生工生物工程(上海)股份有限公司合成,引物序列见表1。
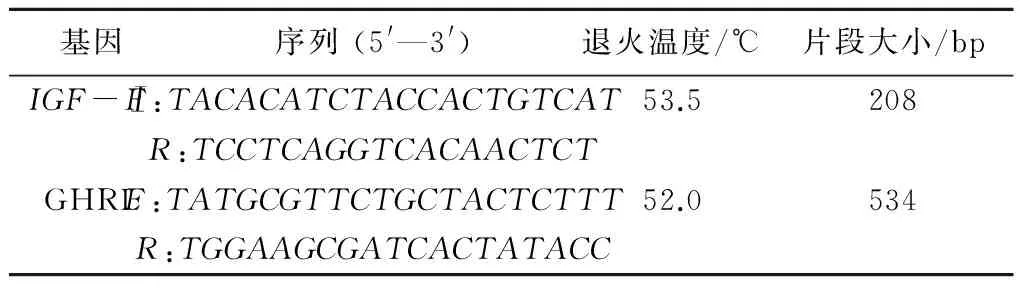
表1 IGF-Ⅰ基因和GHRL基因的引物序列
IGF-Ⅰ、GHRL基因的PCR扩增体系均为25μL:GreenMix12.5μL,无菌水10.5μL,上、下游引物各0.5μL,基因组DNA1μL。
IGF-Ⅰ基因扩增条件:94 ℃预变性5min;94 ℃变性30s,53.5 ℃退火30s,72 ℃延伸30s,共35个循环;最后72 ℃延伸10min;10 ℃保存。GHRL基因扩增条件:94 ℃预变性2min30s;94 ℃变性30s,52.0 ℃退火45s,72 ℃延伸45s,共32个循环;最后72 ℃延伸10min;10 ℃保存。
将上述PCR扩增产物纯化后送到广州华大基因公司,利用ABI-3T30仪器进行测序。
1.3基因型检测
IGF-Ⅰ基因酶切反应体系为20μL:BsmⅠ酶0.5μL,10×NEBuffer2μL,PCR产物17.5μL。65 ℃条件下反应15min。GHRL基因酶切反应体系为20μL:pflMⅠ酶0.5μL,10×NEBuffer2μL,PCR产物17.5μL。37 ℃条件下反应15min。酶切产物用1.2%琼脂糖凝胶电泳检测,用凝胶成像分析仪拍照并进行基因分型。
1.4数据统计与分析
统计不同基因型个体的数量,计算基因型频率,并对多态性片段的基因型分布进行卡方适合性检验。利用SPSS17.0统计软件中的GLM模型对基因多态性与体质量间的相关性进行最小二乘分析,统计结果均以平均值±标准误表示。
2 结果与分析
2.1IGF-Ⅰ、GHRL基因的PCR扩增结果
从图1和图2可以看出,IGF-Ⅰ、GHRL基因特异性扩增良好,没有出现非特异性条带和引物二聚体,扩增片段长度与预期大小相符,可以进一步用来测序和酶切鉴定。
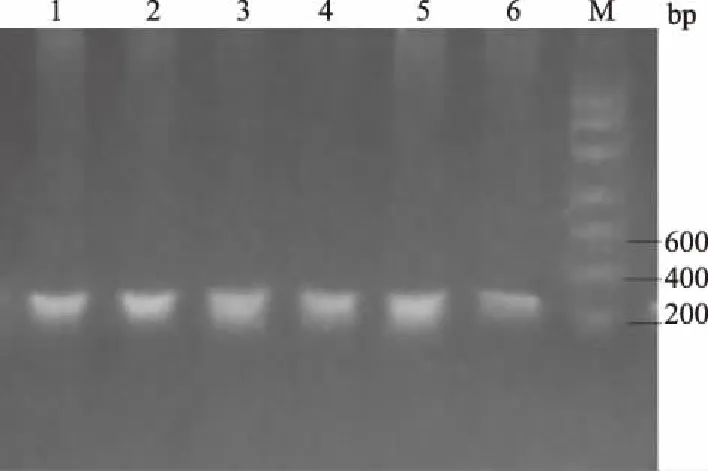
M.DNA Marker; 1—6.PCR扩增产物图1 IGF-Ⅰ基因的PCR扩增结果
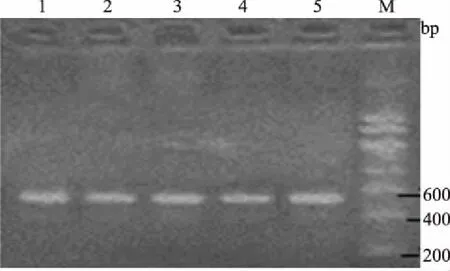
M.DNA Marker; 1—5.PCR扩增产物图2 GHRL基因的PCR扩增结果
2.2IGF-Ⅰ、GHRL基因测序结果
对IGF-Ⅰ基因的测序结果进行分析,发现1个突变位点,突变位点在第181位,由A突变为G,导致密码子GAA突变为GAG,但没有改变编码氨基酸,属于同义突变。同时,对突变位点限制性酶切位点分析发现,未突变的IGF-Ⅰ基因能被BsmⅠ酶切,突变后的IGF-Ⅰ基因则不能被切开。可见,突变改变了酶切位点。
对GHRL基因的测序结果进行分析,发现10个突变位点,分别在第124位(T→G)、125位(T→G)、234位(T→C)、262位(G→A)、410位(G→A)、421位(G→A)、450位(G→A)、480位(G→A)、510位(G→A)、519位(G→A)。这些突变位点都不在GHRL基因编码区上,因此不改变氨基酸序列。但对突变位点的限制性酶切位点分析发现,第124位的碱基突变影响了pflMⅠ酶切位点,未突变的GHRL基因能被pflMⅠ切开,而突变后的GHRL基因则
不能被切开。
2.3IGF-Ⅰ、GHRL基因的基因型分析结果
IGF-Ⅰ基因的PCR产物经BsmⅠ酶切后发现,只存在1种基因型(图3)。因此,不需要进行该基因型分布的卡方适合性检验以及基因型与体质量的关联分析。
GHRL基因的PCR产物经pflMⅠ酶切(图4),发现3种基因型,分别为BB、AB、AA基因型。其中,BB型样本41个,AB型18个,AA型41个。
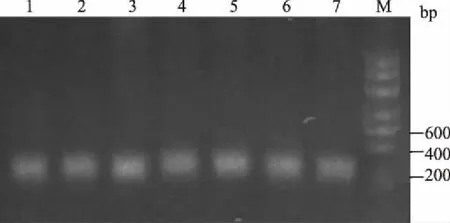
M.DNA Marker; 1—7.酶切产物图3 IGF-Ⅰ基因PCR产物的酶切结果
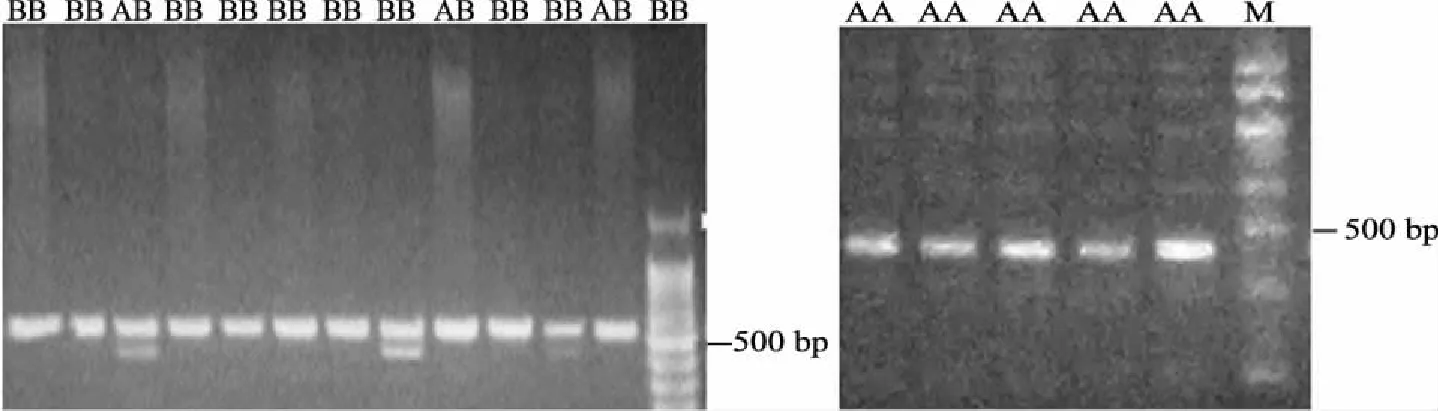
图4 GHRL基因PCR产物的酶切结果
2.4GHRL基因多态性分析
惠阳胡须鸡GHRL基因AA、AB、BB型3种基因型及等位基因的频率见表2,A和B等位基因的频率均为0.5;χ2适合性检验表明,该位点没有处于哈代-温伯格平衡状态,表明该基因没有处于遗传平衡状态。

表2 惠阳胡须鸡GHRL基因的基因型和等位基因频率
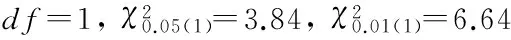
2.5GHRL基因型与惠阳胡须鸡体质量的关联分析
由表3可知,GHRL基因多态性与93日龄惠阳胡须鸡体质量不存在显著相关性,3种基因型个体体质量间的差异均不显著。

表3 GHRL基因多态性与生长性状的关联分析
注:同列相同字母表示差异不显著(P>0.05)。
3 结论与讨论
3.1IGF-Ⅰ基因与GHRL基因多态性
SNPs指基因组中某个单核苷酸变异引起的DNA序列多态性,包括DNA中的碱基替换、颠换(嘧啶和嘌呤之间的替换)、单个碱基的插入或缺失等的变化。赵秀华等[3]在京海黄鸡中发现,IGF-Ⅰ基因外显子3序列的60bp处有A→G突变,属于同义突变,χ2独立性检验显示,京海黄鸡在该位点处于哈代-温伯格平衡状态。丁馥香等[16]在边鸡中发现,IGF-Ⅰ基因第3外显子区域存在2个碱基的突变,一个是同义突变,另一个突变导致了编码氨基酸的改变。Amills等[17]对鸡IGF-Ⅰ基因 5′侧翼区分析,发现了A→C突变。这些结果说明,鸡IGF-Ⅰ基因第3外显子为易发生突变的区域。本研究发现,在惠阳胡须鸡的IGF-Ⅰ基因第3外显子区域中检测到1个突变位点(A→G),属于同义突变,改变了BsmⅠ酶切位点。通过酶切鉴定,发现只有1种基因型,推测该基因位点在该胡须鸡保种群中已经纯合。
Nie等[15]在5个中外鸡品种和1个F2代群体中,通过对GHRL基因5′侧翼区扩增,发现了6个SNPs。方梅霞等[18]在杏花鸡×隐性白洛克鸡F2代资源群体中,用同样的引物对GHRL基因5′侧翼区进行扩增测序,发现相同的6个SNPs。本研究中,用同样的引物进行扩增测序,在GHRL基因5′侧翼区发现了10个SNPs,其中5个为未曾报道的新SNPs。以上结果提示,GHRL基因5′侧翼区为碱基突变的热点区域。有研究表明,非编码区存在着影响基因表达调控的元件,如启动子、增强子等,也是基因反式作用因子的特异结合区域[19]。本研究中发现的这些突变位点的变异是否影响GHRL的基因转录效率需要进一步研究。
3.2GHRL基因多态性对惠阳胡须鸡生长性状的影响
近年来,关于GHRL基因的多态性与动物机体生长、脂肪沉积等的相关性研究已经成为家禽育种领域的热点。罗开鹏等[20]研究表明,GHRL基因突变位点(C345T)影响贵州黑山羊生长性状,该位点有望作为山羊生长性状的一个标记辅助选择位点。鸡GHRL基因C2100T位点多态性与鸡的部分生长性状存在显著相关性[13]。
本研究分析了GHRL基因与93日龄惠阳胡须鸡体质量的关系,BB基因型个体的平均体质量较AA基因型高,但3种基因型个体间体质量无显著差异。由于本研究样本量偏少,且只有1个日龄的体质量数据,可能会影响分析结果,所以需进一步扩大样本数量,收集多个日龄的体质量数据,进一步分析惠阳胡须鸡3种基因型与生长性状的关联性,以确定优势基因型,为今后惠阳胡须鸡育种中生长性状的分子标记辅助选择提供理论依据。
[1]OksbjergN,GondretF,VestergaardM.Basicprinciplesofmuscledevelopmentandgrowthinmeat-producingmammalsasaffectedbytheinsulin-likegrowthfactor(IGF)system[J].DomesticAnimalEndocrinology,2004,27(27):219-240.
[2]FloriniJR,EwtonDZ,CoolicanSA.Growthhormoneandtheinsulin-likegrowthfactorsysteminmyogenesis[J].EndocrineReviews,1996,17:481-517.
[3] 赵秀华,王金玉,张跟喜,等.IGF-Ⅰ与IGFBP-1基因对京海黄鸡生长性状的遗传效应分析[J].畜牧兽医学报,2012,43(1):152-158.
[4] 刘大林,王金玉,魏岳,等.京海黄鸡IGF-Ⅰ基因与生长和屠体性状的关联分析[J].中国畜牧杂志,2009 (11):9-12.
[5]KojimaM,HosodaH,DateY,et al.Ghrelinisagrowth-hormone-releasingacylatedpeptidefromstomach[J].Nature,1999,402:656-660.
[6]HayashidaT,MurakamiK,MogiK,et al.Ghrelinindomesticanimals:Distributioninstomachanditspossiblerole[J].DomesticAnimalEndocrinology,2001,21(1):17-24.
[7]KojimaM,KangawaK.Ghrelin,anorexigenicsignalingmoleculefromthegastrointestinaltract[J].CurrentOpinioninPharmacology,2002,2(6):665-668.
[8]TschopM,SmileyDL,HeimanML.Ghrelininducesadiposityinrodents[J].Nature,2000,407:908-913.
[9]NakazatoM,MurakamiN,DateY,et al.Aroleforghrelininthecentralregulationoffeeding[J].Nature,2001,409:194-198.
[10]WrenAM,SmallCJ,AbbottCR,et al.Ghrelincauseshyperphagiaandobesityinrats[J].Diabetes,2001,50(11):2540-2547.
[11] 张爱玲,张丽,杨明明,等.鸟枪法筛选枯草芽孢杆菌基因强启动子及对黄牛GHRL基因的表达[J].华南农业大学学报,2011,32(3):92-96.
[12] 李俊营,詹凯,许月英,等.巢湖鸭Ghrelin基因外显子3的单核苷酸多态性及其对屠体性状的影响[J].安徽农业科学,2010,38(14):7379-7381.
[13] 何丹林,方梅霞,聂庆华,等.鸡Ghrelin基因C2100T位点与生长和脂肪性状的相关性 [J].广东农业科学,2007(4):73-81.
[14]FangM,NieQ,LuoC,et al.An8bpindelinexon1ofGhrelingeneassociatedwithchickengrowth[J].DomesticAnimalEndocrinology,2007,32(3):216-225.
[15]NieQ,FangM,XieL,et al.Molecularcharacterizationoftheghrelinandghrelinreceptorgenesandeffectsonfatdepositioninchickenandduck[J].JournalofBiomedicineandBiotechnology,2009.doi:10.1155/2009/567120.
[16] 丁馥香,张跟喜,王金玉,等.边鸡胰岛素样生长因子Ⅰ基因(IGF-Ⅰ)外显子 3 的多态性及其与繁殖性能的关系[J].农业生物技术学报,2010,18 (2): 313-317.
[17]AmillsM,JiménezN,VillalbaD,et al.Identificationofthreesinglenucleotidepolymorphismsinthechickeninsulin-likegrowthfactor1and2genesandtheirassociationswithgrowthandfeedingtraits[J].PoultryScience,2003,82(10):1485-1493.
[18] 方梅霞,徐海平,谢亮,等.GHRL基因 5′侧翼区多态性对鸡生长和屠体性状的影响[J].中国农业科学,2011,44(12):2567-2574.
[19]LatchmanDS.Generegulation-Aeukaryoticperspective[M].London:StanleyThornesLtd.,1998.
[20] 罗开鹏,宋桃伟,孙岩岩,等.山羊GHRL基因多态性及其与体重、体尺性状的关系研究[J].广东农业科学,2014,41(6):162-165.
StudyonSingleNucleotidePolymorphism(SNP)ofIGF-ⅠandGHRLGeneinHuiyangBeardedChickens
LIHongwei1,ZHONGDanlong1,CHENYuan1,WUXiaoyan1,ZOUZhiguan2
(1.DepartmentofLifeScience,HuizhouUniversity,Huizhou516007,China;2.GuangdongJinzhongAgricultureandAnimalHusbandryCo.,Ltd.,Huizhou516021,China)
In this study,IGF-ⅠandGHRLgeneswerechosenascandidategenesofgrowthtraitsinHuiyangbeardedchicken.Sequencingandenzymedigestionwereappliedtoassessthesinglenucleotidepolymorphismofthetwogenesandthegenerallinearmodelwasusedtoanalyzeassociationbetweendifferentgenotypesandbodyweightonthe93rddayinHuiyangbeardedchicken.Theresultsshowedthatonemutation(A181G)wasfoundinIGF-Ⅰgene,butthemutationwassynonymous.OnlyonegenotypewasdetectedbyBsmⅠenzymedigestioninIGF-Ⅰgene,whichindicatedthatthemutationsitehadbeenhomozygousintheconservationpopulationofHuiyangbeardedchicken.Theresultsalsoshowedthatonemutation(T124G)wasdetectedinGHRLgene.ThemutationchangedthecuttingsiteofpflMⅠenzyme.Threegenotypes(AA,AB,BB)oftheGHRLgeneweredetectedbythepflMⅠenzymedigestion.TheassociationanalysesshowedthattherewasnosignificantdifferenceamongindividualswiththreegenotypesinHuiyangbeardedchickenonthe93rdday.
Huiyangbearedchicken; IGF-Ⅰgene; GHRLgene;geneticpolymorphism
2017-03-16
广东省普通高校特色创新项目(2014KTSCX178);惠州学院博士启动基金项目(156020021)
李红伟(1972-),男,云南禄丰人,讲师,博士,主要从事家禽遗传育种研究。E-mail:lhwcau@163.com
S831
:A
: 1004-3268(2017)09-0152-04
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:33:26
艺术品鉴(2021年14期)2021-06-05 09:41:52
——第一届广东省吉他(惠阳)邀请赛暨惠阳吉他文化艺术节举行
岭南音乐(2017年3期)2017-07-18 11:59:40
西南农业学报(2016年6期)2016-04-16 05:12:47
神州民俗(2016年19期)2016-02-02 15:42:46
法医学杂志(2015年4期)2016-01-06 12:36:36
现代检验医学杂志(2015年6期)2015-02-06 01:44:02
实验动物与比较医学(2014年5期)2014-02-28 14:53:10
河南医学研究(2014年7期)2014-02-27 14:53:42
中国糖料(2013年1期)2013-01-22 12:28:23
