
1株PRRSV类NADC30毒株的分离鉴定及序列分析
2017-09-13 09:20崔丹丹王傲杰王新港彭志锋王川庆
河南农业科学 2017年9期
崔丹丹,王傲杰,王新港,周 峰,彭志锋,王川庆
(河南农业大学 牧医工程学院,河南 郑州 450002)
1株PRRSV类NADC30毒株的分离鉴定及序列分析
崔丹丹,王傲杰,王新港,周 峰,彭志锋,王川庆*
(河南农业大学 牧医工程学院,河南 郑州 450002)
为了解猪繁殖与呼吸综合征病毒(PRRSV)流行株的变异情况,将PRRSV阳性病料处理后接种Marc-145细胞,成功分离到1株PRRSV毒株,将其命名为HENZXC-4,并对其全基因组序列进行测定和分析。结果表明,HENZXC-4为未发生重组的类NADC30毒株,其NSP2蛋白存在131个氨基酸不连续缺失,且其GP5蛋白存在1个氨基酸插入。
猪繁殖与呼吸综合征病毒; 分离鉴定; 类NADC30株; 序列分析
猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS)是由猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)引起的一种以妊娠母猪繁殖障碍及各生长阶段猪的呼吸道症状为特征的传染病,最早于1987年在美国报道,目前已成为严重威胁全球养猪业的主要疫病之一[1]。PRRSV基因组全长约15 kb,包含至少10个开放式阅读框(open reading frames,ORFs),具有高度变异的特点,在进化过程中可通过突变、缺失、插入、重排及重组等方式进行遗传变异,进而导致病毒的多样性。
我国作为世界上养猪大国之一,自1995年底报道出现PRRS后,养猪业已被该病困扰长达20多年。我国分离到的第1株PRRSV为经典毒株CH-1 a[2]。2006年,高致病性PRRSV(HP-PRRSV)引发了一场猪的“高热病”,其NSP2蛋白与经典毒株相比存在不连续的30个氨基酸的特征性缺失,为该类毒株的特征性缺失。随后,该类毒株逐渐成为国内PRRSV优势流行毒株[3-7]。然而,近年来针对该优势流行毒株的弱毒活疫苗的广泛使用造成了一些疫苗回复株在田间流行的现象[8]。此外,自2012年周峰等[9]首次发现与美国NADC30毒株高度同源的国内毒株以来,陆续有学者报道在我国其他地方有类NADC30毒株出现,且逐渐成为继HP-PRRSV之后的又一田间优势流行毒株,该类毒株在NSP2蛋白存在131个氨基酸不连续缺失,并易与其他类型PRRSV毒株发生重组[10-15]。目前,我国出现了多种类型PRRSV毒株共存的局面,且各类毒株仍在不断地变异,加大了该病的防控难度。分析PRRSV遗传变异情况,对指导PRRS防控工作具有重要意义[16-18]。鉴于此,本研究分离了PRRSV,并对其进行全基因组序列分析,以了解河南省PRRSV流行株的变异特征,旨在为PRRS的防控及疫苗研发提供参考。
1 材料和方法
1.1病料采集
PRRSV阳性病料采自河南省驻马店市某HP-PRRSV疫苗免疫猪场病死猪只的肺脏、脾脏及淋巴结等,取适量研磨后于-70 ℃冰箱保存备用。
1.2菌株、毒株及载体
pMD18-T克隆载体购自宝生物工程(大连)有限公司;E.coliDH5α感受态细胞及PRRSV阳性对照毒株(HP-PRRSV毒株HENZZ-8)由河南农业大学传染病实验室保存。
1.3主要试剂及细胞
RNAiso Plus试剂、随机引物、dNTP、RNA酶抑制剂(RNase Inhibitor)、鼠源反转录酶(M-MLV)、DL2000 DNA Marker、5′ Full RACE 试剂盒和3′ Full RACE试剂盒等均购自宝生物工程(大连)有限公司;琼脂糖购自Sigma公司;2×ExTaqMaster Mix(含染料)购自北京康为世纪生物科技有限公司;DMEM培养基、胰酶购自Gibco公司;胎牛血清(FBS)购自北京四季青公司;DAPI染色液及FITC标记的羊抗鼠IgG荧光抗体购自武汉博士德生物工程有限公司;抗美洲型PRRSV N蛋白单克隆抗体由河南农业大学田克恭教授惠赠;Marc-145细胞由河南农业大学杨国宇教授惠赠;其他试剂均为国产分析纯。
1.4引物设计与合成
使用郭天准等[19]设计的引物P3-F/R(F: 5′-GTGTCAGGCATTGTGGCTGTG-3′,R: 5′- CA-TTATTGGCGTGTAGGTGATAGAAAA-3′)对细胞培养物进行RT-PCR检测,使用周峰[20]设计的针对类NADC30毒株的10对特异性引物对全基因组进行分段扩增,基因组5′ 端和3′ 端分别采用5′ Full RACE 试剂盒 和3′ Full RACE 试剂盒进行扩增(表1)。引物均由生工生物工程(上海)股份有限公司合成。

表1 PRRSV全基因组扩增引物
1.5PRRSV流行毒株的分离与鉴定
将保存的PRRSV阳性病料经离心、过滤后取1 mL 滤液接种于生长至单层的Marc-145细胞,置于37 ℃、5% CO2培养箱内孵育1 h后,加入5 mL含2%FBS的DMEM培养基继续培养,并观察细胞病变(CPE),2~4 d收集细胞培养物。盲传3代后进行RT-PCR检测,若出现CPE且PCR检测结果为阳性,则将该细胞培养物继续传代3次,观察CPE的稳定性并进行RT-PCR检测,同时进行间接免疫荧光(IFA)试验对培养物进行鉴定。
1.5.1 细胞培养物总RNA的提取及病毒核酸检测 取细胞上清250 μL,参照RNAiso Plus说明书提取总RNA,然后进行RT-PCR扩增。反转录反应体系(20 μL):RNA 模板1 μg、随机引物(20 pmol/μL) 1 μL、dNTP(10 mmol/μL) 1 μL、5×Buffer 4 μL、反转录酶M-MLV(200 U/μL) 0.5 μL、RNA酶抑制剂(40 U/μL) 0.5 μL,并用1‰的DEPC水补充至20 μL。反应条件:42 ℃ 1 h,95 ℃ 5 min,得到的cDNA用于PCR扩增。PCR反应体系(50 μL):2×ExTaqMaster Mix 25 μL、上下游引物(20 pmol/μL)各1 μL、cDNA模板5 μL,补加1‰的DEPC水至50 μL。反应条件:94 ℃预变性5 min;94 ℃ 50 s,56 ℃ 30 s,72 ℃ 1 min,35个循环;最后72 ℃延伸10 min。取5 μL PCR产物用1%琼脂糖凝胶进行电泳检测。以不加病毒的细胞培养物作为阴性对照,以1‰ DEPC水的模板为空白对照。
1.5.2 IFA鉴定 在12孔板中用常规方法进行IFA试验,试验结束后每孔即刻加入100 μL DAPI染色液并室温避光作用5 min进行细胞核染色,PBS清洗,晾干后置于荧光显微镜下观察。同时设立PRRSV阳性对照组(HENZZ-8毒株)和阴性对照组(不加病毒)。
1.6PRRSV分离株生长曲线绘制
PRRSV分离株按MOI=0.01接种Marc-145细胞,在接种后每隔12 h(直至96 h)分别取样进行TCID50测定,绘制出各毒株生长曲线。同时设立PRRSV阳性对照组HENZZ-8。
1.7PRRSV分离株全基因组的分段扩增、克隆及测序
用PRRSV全基因引物扩增出分离株基因组的各片段,将这些片段的PCR产物回收纯化后与pMD18-T克隆载体连接,转化E.coliDH5α感受态细胞,经氨苄青霉素筛选及PCR鉴定后,将阳性菌液送生工生物工程(上海)股份有限公司测序。
1.8PRRSV分离株全基因组的遗传变异分析
用DNAStar软件中的MegAlign对分离株的全基因进行序列比对和相似性分析,利用MEGA 7.0软件中的邻接法对分离株进行全基因遗传进化分析并绘制系统进化树,利用RDP 4.0进行重组分析。
1.9PRRSV分离株NSP2及GP5氨基酸序列分析
用DNAStar软件中的MegAlign对由NSP2和 ORF5基因推导的氨基酸进行比对分析。
2 结果与分析
2.11株PRRSV流行株的分离与鉴定
经病毒传代及RT-PCR(图1)和IFA(图2)鉴定阳性后,成功分离出1株PRRSV毒株,命名为HENXC-4。初代培养60 h出现CPE,连续传代3次后可出现稳定且典型的CPE。
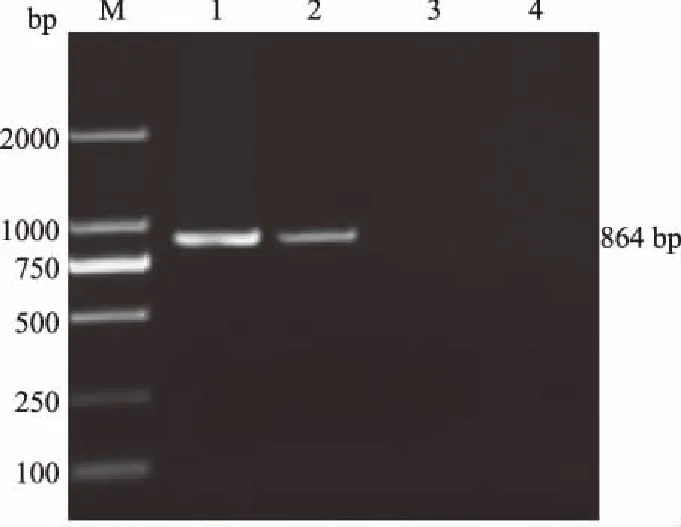
M.DL2000 DNA Marker; 1.阳性对照; 2.细胞培养物; 3.阴性对照; 4.空白对照图1 细胞培养物的RT-PCR检测结果
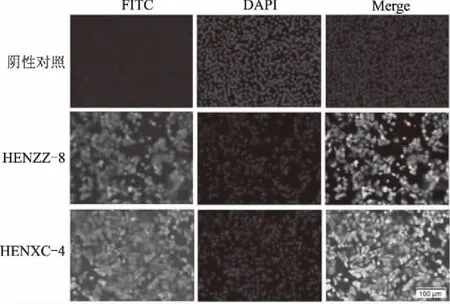
图2 细胞培养物的IFA鉴定结果(100×)
2.2PRRSV分离株在Marc-145细胞上的生长特性
分离株HENXC-4和对照株HENZZ-8接种后,定点收毒,测定TCID50,绘制特异性生长曲线(图3)。在接种60 h内,分离株HENXC-4和对照株HENZZ-8具有相似的生长趋势,HENXC-4较HENZZ-8生长略为缓慢;但对照株HENZZ-8在60 h后有明显的下降趋势,而分离株HENXC-4未出现明显的下降趋势。2株毒株均能很好地适应Marc-145细胞,病毒滴度最高能达到约107TCID50/mL。
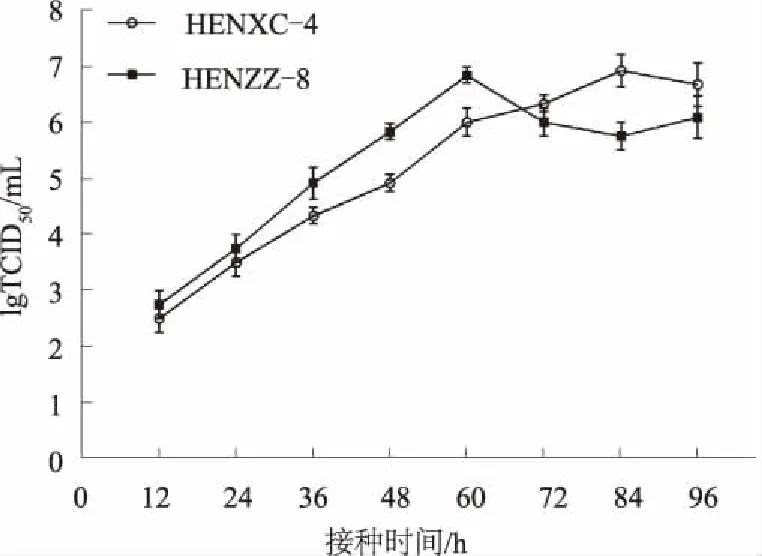
图3 PRRSV分离株生长曲线
2.3全基因组的遗传变异特征
成功扩增得到分离株HENXC-4各基因片段。经DNAMAN软件拼接获得其全基因组序列,全长为15 023 bp,不包含Poly(A)。GenBank登录号为KU950371。
2.3.1 全基因组核苷酸序列及其推导的氨基酸序列相似性比较 由表2可见,分离株HENXC-4的核苷酸序列与欧洲型(Ⅰ型)代表毒株LV的相似性为62.0%,与美洲型(Ⅱ型)代表毒株VR-2332的相似性为85.3%,与国内经典毒株CH-1a的相似性为84.4%,与国内HP-PRRSV JXA1的相似性为83.3%,而与美国NADC30毒株的相似性最高,为95.6%。重组分析结果表明,HENXC-4并未与其他毒株发生重组。
HENXC-4 与代表毒株NADC30、JXA1、CH-1a、VR-2332、LV的各段序列比对结果见表2和表3。HENXC-4的5′和3′非编码区(UTR)核苷酸序列与其他5个代表毒株的相似性分别为52.7%~96.3%和70.2%~98.7%,且均与NADC30相应片段的相似性最高。HENXC-4各编码区氨基酸序列与NADC30、JXA1、CH-1a、VR-2332和LV的相似性分别为90.6%~100.0%、66.8%~96.4%、67.1%~96.0%、70.8%~100.0%和29.6%~81.2%。在13个非结构蛋白中,NSP2是HENXC-4同其他代表毒株氨基酸相似性较低的蛋白质,仅为29.6%~90.6%。HENXC-4的ORF2a—7区域的核苷酸序列与其他5个代表毒株的相似性为67.2%~96.2%,而在该区域编码的8个结构蛋白中,HENXC-4的氨基酸序列与其他5个代表毒株的相似性为51.0%~97.7%,且ORF3~ORF5a是结构蛋白氨基酸序列比对中相似性较低的蛋白质,仅为51.0%~96.6%,但与M蛋白的氨基酸序列相似性较高,为79.3%~97.7%。值得一提的是,除NSP1α外,HENXC-4各非结构及结构蛋白的氨基酸序列相似性均与NADC30最高。

表2 HENXC-4与PRRSV参考株核苷酸序列相似性 %

表3 HENXC-4与PRRSV参考株氨基酸序列相似性 %
2.3.2 分离株全基因组的遗传进化特征 由HENXC-4及28个参考株的全基因序列构成的系统进化树显示,分离株HENXC-4属于以NADC30为代表的亚群4,且与国内优势流行毒株HP-PRRSV的代表株JXA1的亲缘关系较远(图4)。
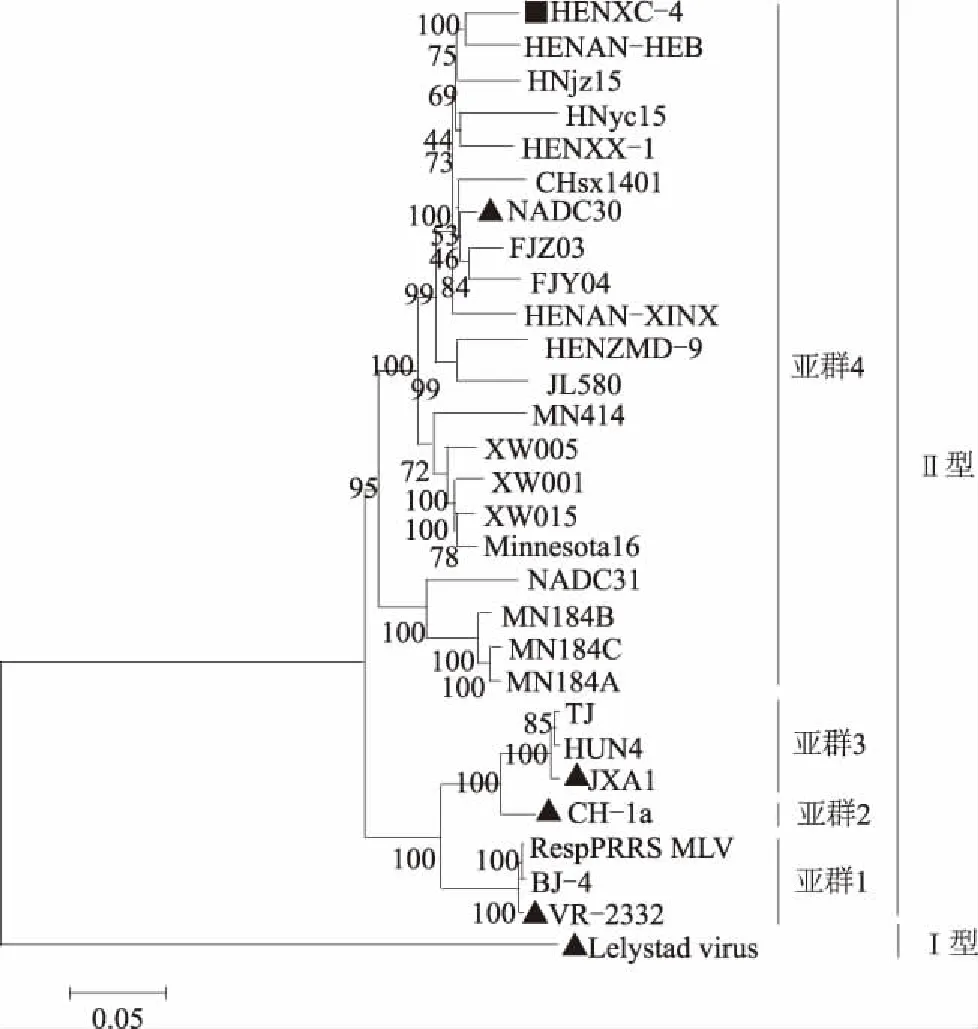
▲为代表性参考毒株;■为分离株HENXC-4图4 基于HENXC-4株全基因组核苷酸序列的遗传进化分析
2.4分离株NSP2和GP5氨基酸序列特征
NSP2的氨基酸序列分析结果显示,HENXC-4与NADC30的相似性最高,为90.6%。序列比对结果表明,HENXC-4与NADC30有着一样的缺失特征,即存在131个氨基酸不连续缺失,这一缺失特征与国内HP-PRRSV的缺失特征不同(图5)。
GP5的氨基酸序列分析结果显示,HENXC-4与NADC30的相似性最高,为93.5%。此外,序列比对结果发现,同其他参考毒株相比,HENXC-4在GP5第58位有1个氨基酸的插入(图6),且这一插入特点在其他类NADC30毒株中未发现。
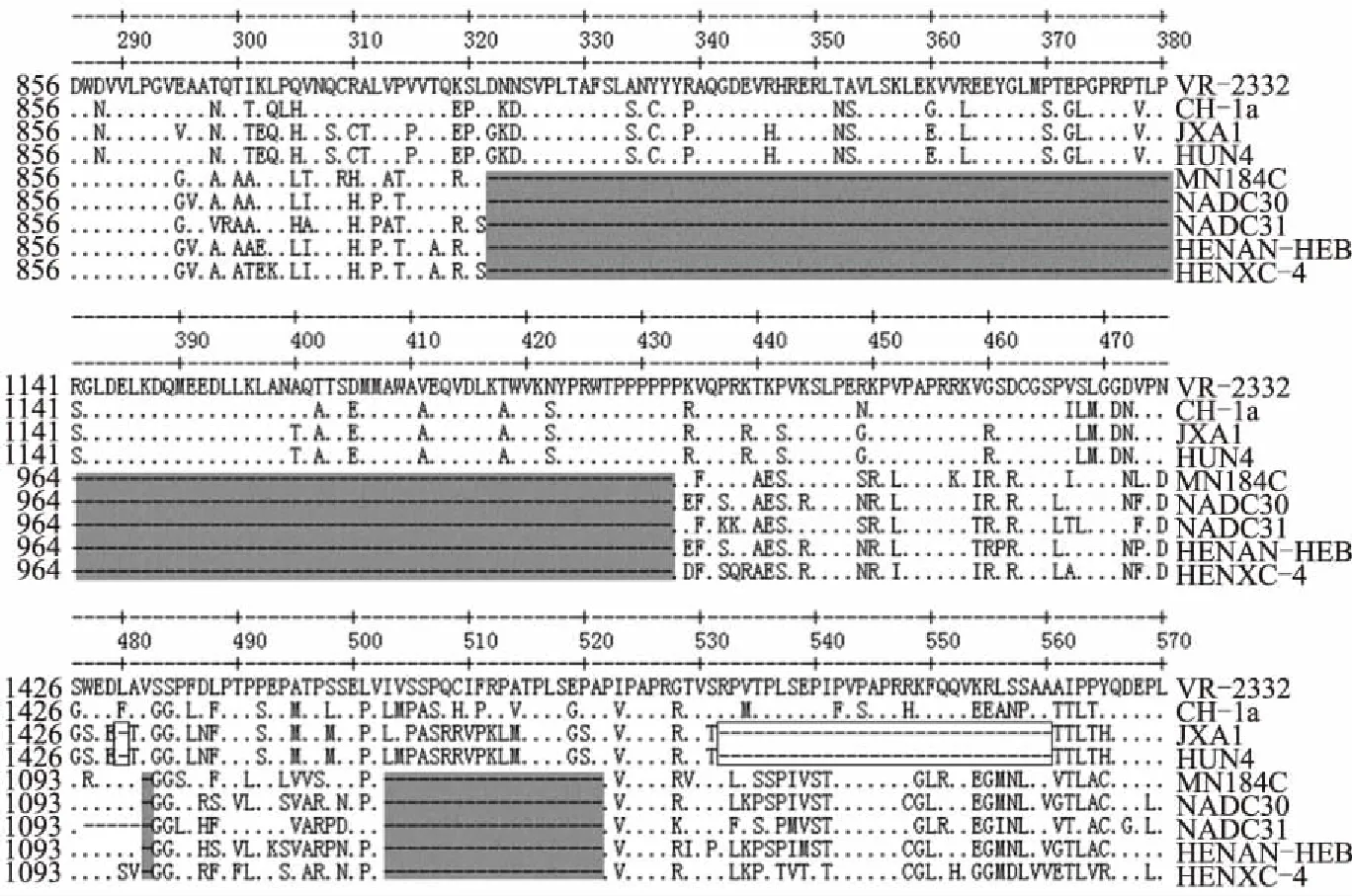
类NADC30株缺失部分用灰色阴影标注;HP-PRRSV株缺失部分用黑色方框标注图5 分离株HENXC-4部分NSP2基因推导的氨基酸序列比对分析结果
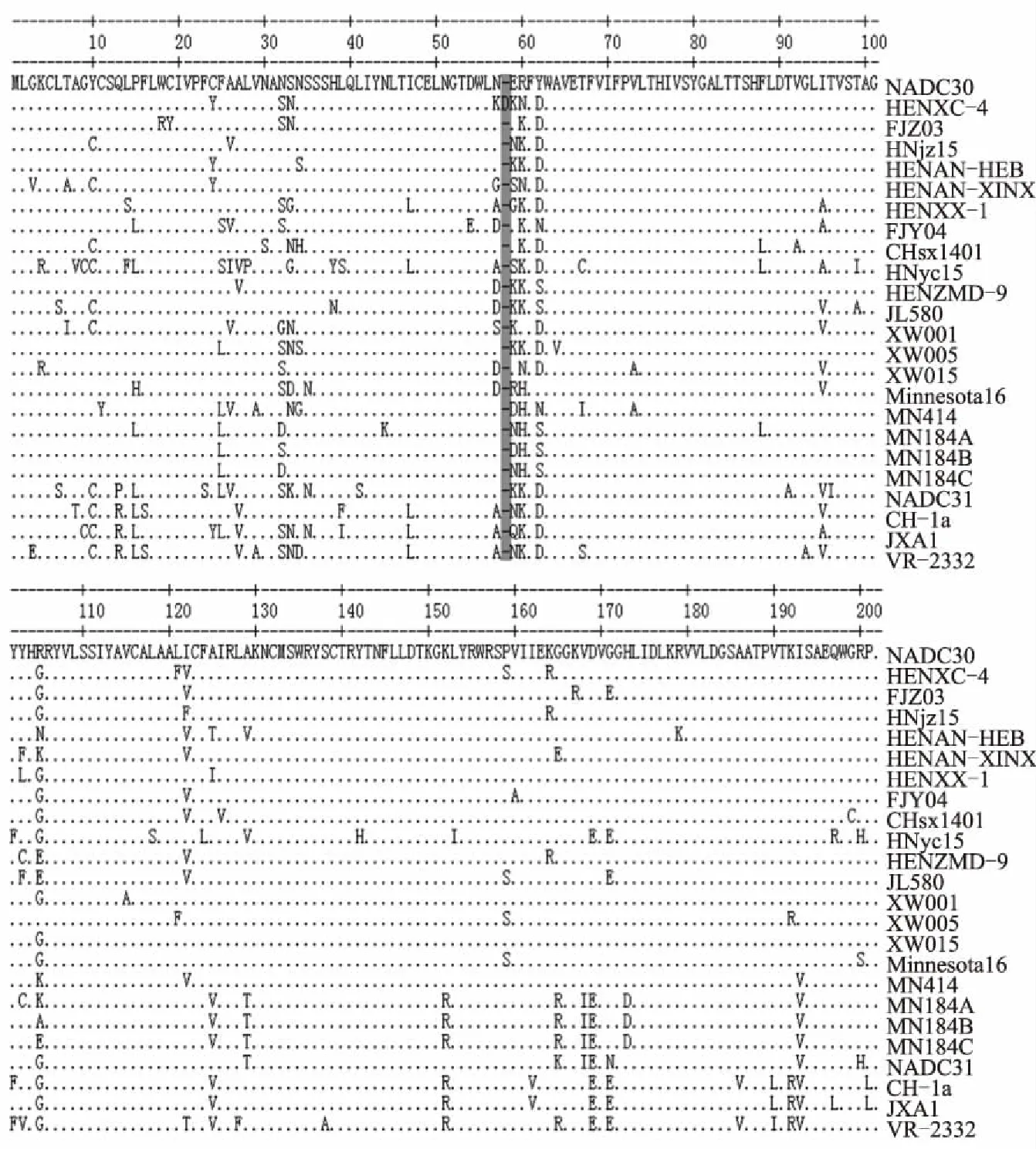
插入部分用灰色阴影标注图6 分离株HENXC-4 ORF5基因推导的氨基酸序列比对分析结果
3 结论与讨论
PRRSV自从在我国发现以来,已历经20多年的流行及演变,但继CH-1a之后的BJ-4、HB-1(sh)/2002和HB-2(sh)/2002等经典型毒株并未引发较为严重的疫情[21-22]。HP-PRRS首先于2006年在我国江西省暴发,并迅速蔓延至全国大部分地区,其以高发病率和高死亡率为特征,给养猪业造成了巨大的经济损失[23]。引发该病的病原HP-PRRSV也逐渐成为国内PRRSV优势流行毒株。然而,自2012年以来,与美国NADC30毒株具有相同缺失特征的毒株在我国各地相继被发现,并逐渐成为继HP-PRRSV之后的又一田间优势流行毒株,且易与其他类型PRRSV毒株发生重组[10-15]。NADC30毒株最早在美国发现,近年来随着国内养猪业大规模引进加系和美系种猪,类NADC30毒株相继在国内很多地区被发现,提示类NADC30毒株可能与从美洲国家引进种猪有关[13]。另外值得注意的是,现有的疫苗包括经典疫苗(VR-2332和R98)和高致病性疫苗(JXA1-P80、HuN4-F112、GDr180和TJM-F92),均不能有效地保护猪只免受类NADC30毒株的侵害[23-24]。因此,迫切需要对类NADC30毒株,尤其是未发生重组的类NADC30毒株的分子特征、遗传进化及致病性等方面进行深入研究,为PRRS的防控及新疫苗的研发奠定基础。
本研究成功分离鉴定到1株PRRSV毒株HENXC-4。全基因序列比对分析结果显示,HENXC-4与美国NADC30毒株的全基因核苷酸序列相似性最高,且除NSP1α外,HENXC-4各非结构及结构蛋白的氨基酸序列相似性均与NADC30最高,表明HENXC-4可能为1株类NADC30株。全基因遗传进化分析显示,分离株HENXC-4属于以NADC30为代表的亚群4,这一结果进一步指示HENXC-4为1株类NADC30毒株。NSP2的氨基酸序列比对结果显示,HENXC-4和NADC30均存在131个氨基酸的不连续缺失,这一结果再次证实HENXC-4为1株类NADC30株。在已报道的类NADC30分离株中,同其他类型毒株发生重组的毒株居多,而未发生重组的类NADC30分离株数量十分有限,这使得全面了解类NADC30毒株的研究受到限制。而本研究所分离到的PRRSV HENXC-4经重组分析后发现,其未与其他PRRSV类型毒株发生重组,是1株与NADC30毒株相似性非常高的类NADC30毒株,为进一步研究该类毒株提供了原材料,并为疫苗研发奠定了基础。此外,HENXC-4的GP5蛋白中第58位有1个氨基酸的插入,且这一插入特点在已报道的所有类NADC30毒株中均未发现。因此,如果后续用该毒株研发类NADC30疫苗,其在GP5蛋白中插入的氨基酸可作为一天然标记,用于区分野毒株和疫苗株。
大部分已报道的类NADC30分离株,尤其是未与其他PRRSV发生重组的NADC30株,是由猪肺泡巨噬细胞(PAM)培养分离所得,并不能很好地适应Marc-145细胞。而本研究中,类NADC30株HENXC-4的分离并不依赖PAM,且可在Marc-145细胞上很好地增殖,不需要分离培养原代细胞PAM,省时省力、节约成本。
[1] Rossow K D.Porcine reproductive and respiratory syndrome[J].Veterinary Pathology,1998,35(1):1-20.
[2] 郭宝清,陈章水,刘文兴,等.从疑似PRRS流产胎儿分离PRRSV的研究[J].中国预防兽医学报,1996(2):1-5.
[3] Song J,Shen D,Cui J,etal.Accelerated evolution of PRRSV during recent outbreaks in China[J].Virus Genes,2010,41(2):241-245.
[4] Tian K,Yu X,Zhao T,etal.Emergence of fatal PRRSV variants:Unparalleled outbreaks of atypical PRRS in China and molecular dissection of the unique hallmark[J].PLoS One,2007,2(6):e526.
[5] Shi M,Holmes E C,Brar M S,etal.Recombination is associated with an outbreak of novel highly pathogenic porcine reproductive and respiratory syndrome viruses in China[J].Journal of Virology,2013,87(19):10904-10907.
[6] 卢晓艳,周永辉,李伟娟,等.猪繁殖与呼吸综合征病毒河南分离株NSP2和GP5基因遗传变异分析[J].华北农学报,2012,27(2):100-104.
[7] 王凤雪,温永俊,刘准,等.高致病性PRRSV TJ株和致弱毒TJM株Nsp2测序及结构分析[J].河南农业大学学报,2011,45(1):86-90.
[8] Jiang Y F,Xia T Q,Zhou Y J,etal.Characterization of three porcine reproductive and respiratory syndrome virus isolates from a single swine farm bearing strong homology to a vaccine strain[J].Veterinary Microbiology,2015,179(3/4):242-249.
[9] 周峰,常洪涛,赵军,等.2012—2013年猪繁殖与呼吸综合征病毒河南流行株的分离鉴定及分子流行病学调查[J].中国兽医学报,2014,34(9):1398-1404,1410.
[10] Zhao K,Ye C,Chang X B,etal.Importation and recombination are responsible for the latest emergence of highly pathogenic porcine reproductive and respiratory syndrome virus in China[J].Journal of Virology,2015,89(20):10712-10716.
[11] Zhou L,Wang Z,Ding Y,etal.NADC30-like strain of porcine reproductive and respiratory syndrome virus,China[J].Emerging Infectious Diseases,2015,21(12):2256-2257.
[12] Ji G,Li Y,Tan F,etal.Complete genome sequence of an NADC30-like strain of porcine reproductive and respiratory syndrome virus in China[J].Genome Announcements,2016,4(6):303-316.
[13] Li C,Zhuang J,Wang J,etal.Outbreak investigation of NADC30-like PRRSV in South-East China[J].Transboundary and Emerging Diseases,2016,63(5):474-479.
[14] Sun Z,Wang J,Bai X,etal.Pathogenicity comparison between highly pathogenic and NADC30-like porcine reproductive and respiratory syndrome virus[J].Archives of Virology,2016,161(8):2257-2261.
[15] 王林建,郭振华,乔松林,等.河南地区NADC30-like PRRSV毒株的增殖特性与遗传进化分析[J].河南农业科学,2017,46(3):122-128.
[16] 王小敏,何孔旺,周忠涛,等.猪繁殖与呼吸综合征病毒变异株的分离鉴定及遗传变异分析[J].华北农学报,2014,29(1):232-238.
[17] 张美玲,简子健,陆桂丽,等.猪繁殖与呼吸综合征病毒新疆株的分离与鉴定[J].天津农业科学,2013,20(11):31-34.
[18] 韩一超,刘文俊,姚敬明,等.猪繁殖与呼吸综合征病毒Shanxi-6株全基因序列分析[J].华北农学报,2014,29(2):62-65.
[19] 郭天准,常洪涛,崔丹丹,等.2014—2015年河南地区PRRSV的分子检测及NSP2、ORF5基因变异分析[J].病毒学报,2016,32(3):298-307.
[20] 周峰.2012—2013年河南地区猪繁殖与呼吸综合征病毒分子流行病学调查及河南流行株的分离、鉴定[D].郑州:河南农业大学,2014.
[21] Li H,Yang H.Infection of porcine reproductive and respiratory syndrome virus suppresses the antibody response to classical swine fever virus vaccination[J].Veterinary Microbiology,2003,95(4):295-301.
[22] Zhou Z,Ni J,Cao Z,etal.The epidemic status and genetic diversity of 14 highly pathogenic porcine reproductive and respiratory syndrome virus(HP-PRRSV) isolates from China in 2009[J].Veterinary Microbiology,2011,150(3/4):257-269.
[23] Zhang Q,Jiang P,Song Z,etal.Pathogenicity and antigenicity of a novel NADC30-like strain of porcine reproductive and respiratory syndrome virus emerged in China[J].Veterinary Microbiology,2016,197:93-101.
[24] Bai X,Wang Y,Xu X,etal.Commercial vaccines provide limited protection to NADC30-like PRRSV infection[J].Vaccine,2016,34(46):5540-5545.
Isolation,Identification and Sequence Analysis of a NADC30-like Strain of PRRSV
CUI Dandan,WANG Aojie,WANG Xingang,ZHOU Feng,PENG Zhifeng,WANG Chuanqing*
(College of Animal Husbandry and Veterinary Science,Henan Agricultural University,Zhengzhou 450002,China)
To investigate the molecular variation characteristics of porcine reproductive and respiratory syndrome virus(PRRSV) in the field,a PRRSV isolate(HENXC-4) was isolated by inoculating the PRRSV positive samples into the Marc-145 cell layers.The whole genome was then sequenced and analyzed.The sequence analysis showed that HENXC-4 was NADC30-like PRRSV without any recombination phenomenon characterized by a discontinuous 131-amino-acid deletion in the NSP2 region.More importantly,it had a 1-amino-acid insertion in GP5,which was absent in other NADC30-like PRRSV strains.
PRRSV; isolation and identification; NADC30-like strain; sequence analysis
2017-04-10
河南省高校科技创新团队与支持计划资助项目(14IRTSHN015)
崔丹丹(1991-),女,河南焦作人,在读硕士研究生,研究方向:猪繁殖与呼吸综合征病毒的流行、变异及致病性。 E-mail:13723171656@163.com
*通讯作者:王川庆(1954-),男,河南濮阳人,教授,主要从事动物传染病发病机制及防制研究。E-mail:wchuanq@163.com
S852.65
: A
: 1004-3268(2017)09-0132-07
猜你喜欢
数学物理学报(2022年5期)2022-10-09
科学大观园(2022年2期)2022-01-23
文萃报·周二版(2021年47期)2021-12-14
今日农业(2021年11期)2021-08-13
中国生殖健康(2020年4期)2020-12-09
河北画报(2020年8期)2020-10-27
中西医结合肝病杂志(2020年2期)2020-10-27
中成药(2018年7期)2018-08-04
中国猪业(2017年11期)2017-12-11
浙江大学学报(工学版)(2016年2期)2016-06-05
