
气候变化对草原多年生克隆植物无性繁殖的影响
2017-09-08 03:40李卓琳穆春生吴春英
草业科学 2017年8期
李卓琳,穆春生,吴春英
(1.上海市城市化生态过程与生态恢复重点实验室, 华东师范大学生态与环境科学学院,上海 200241; 2.植被生态科学教育部重点实验室, 东北师范大学草地科学研究所,吉林 长春 130024; 3.延边州草原管理站,吉林 延吉 133001)
气候变化对草原多年生克隆植物无性繁殖的影响
李卓琳1,穆春生2,吴春英3
(1.上海市城市化生态过程与生态恢复重点实验室, 华东师范大学生态与环境科学学院,上海 200241; 2.植被生态科学教育部重点实验室, 东北师范大学草地科学研究所,吉林 长春 130024; 3.延边州草原管理站,吉林 延吉 133001)
气候变化是多因子的综合体,常相伴发生,其对地球上各生态系统中的植物、动物、微生物等均有不同程度的影响。克隆植物,是一类兼具有性和无性两种繁殖方式的植物,由于其长期以来对环境的适应,很多克隆植物已放弃或较少地进行有性繁殖,导致其无性繁殖能力在某种程度上可以直接反映地上生产力,所以克隆植物无性繁殖如何响应气候变化成为当前重要的课题。因此,本文从气候变化的几个主要因子出发,综述其对多年生克隆植物无性繁殖的影响,结果发现,绝大部分文献指出CO2升高和氮沉降均促进子株的输出,而降水量的减少抑制或不影响子株的输出,此外,增温时间不同对子株的影响也有很大差异。到目前为止,虽然气候变化中各因子对克隆植物无性繁殖的研究已有大量报道,但关于它们之间交互作用的研究较少。而且可能由于地下取样困难等原因,针对CO2浓度升高、增温、氮沉降等气候变化对地下芽库影响的文献则更少。所以应加强各气候因子之间的交互作用对克隆植物影响的研究工作,因为它不仅可以进一步丰富克隆植物生态学与全球变化生态学的交叉研究内容,而且可以为未来草地管理提供一定的实践依据。
CO2;增温;降水格局;氮沉降;无性繁殖
众所周知,植物种群更新的完成依赖于植物的繁殖过程。而在多年生草地上,克隆植物占据较大比例,且这部分植物由于其有性繁殖能力薄弱,加之幼苗建植困难,绝大部分克隆植物将更多的能量分配给无性繁殖,甚至有一些克隆植物已放弃有性繁殖。所以草地上多年生克隆植物的无性繁殖能力可以很好地预示其生产力的变化。此外,与非克隆植物相比,克隆植物具有生存力、占据新的空间能力、克隆整合能力强等特点,所以它在退化草原恢复中也扮演着重要角色[1]。
目前,CO2作为最主要的人为温室气体,已由工业革命前期的270 μmol·mol-1增加到现在的380 μmol·mol-1,预计到21世纪末将会倍增[2]。全球表面温度增加也已经成为毋庸置疑的事实[3-6],而且在大陆生态系统夜间温度增加的幅度可能大于白天。随着温度的变化,降水格局也将发生改变,主要表现为降水总量及降水分布发生变化,并且以单次降水量增加以及降水间隔时间延长为特征的降水事件频发,即发生极端暴雨的几率增加[7-8]。单次降水量的增加只会导致无效水增加,降水间隔时间的延长则意味着干旱期的延长,这会使部分地区的干燥系数增加,土壤水分减少,进而加重中国半干旱及干旱区域的干旱化[9]。此外,在干旱和半干旱地区,氮是继降水量之后限制植物生产力最主要的因子[10],氮沉降也已成为当今备受关注的全球性环境问题之一[11]。
在全球变化日趋严重的情况下,探究多年生克隆植物无性繁殖的变化规律,对种群生态学、全球变化下的退化生态学以及农牧业生产实践等具有重要意义。至今,国内外已开展了大量的关于全球变化对植物个体、种群、群落乃至整个生态系统的研究[12-13],且研究结果差异较大。综合气候变化对种群水平的研究文献发现,对个体光合作用和植被盖度及生产力的研究较多,而对克隆植物繁殖的关注度相对较少(图1),想要完善克隆植物应对全球气候变化的生活史对策,其繁殖是关键。因此,有必要对草地多年生克隆植物无性繁殖如何响应全球气候变化的研究进展作一综述评价。
1 模拟温度与CO2浓度增加、氮沉降及降水变化的方法
有关全球气候变化对植物影响的研究中,国内外学者采用不同的试验生态学方法,这其中包括野外控制试验、温室模拟试验和模型模拟等(表1)。目前增温手段主要有红外加热,其增温幅度基本在1~3 ℃之间[14-17];利用气候室模拟,增温的幅度在2~10 ℃不等[18-19];开顶式气室,增温幅度也在1 ℃左右[20]。而对CO2模拟多采用气候室和开顶式气室,由于费用较大,一般模拟周期也较短[21-23]。模拟增雨或减雨多采用防雨棚或塑料板装置截留自然降水然后人工施加给增雨区域,或根据自然地理降水分布而采样,亦可根据历年来的降水数据及其植被的变化分析[24-26]。模拟氮沉降的则以施加10 g·(m2·a)-1硝酸铵的居多[17,27]。这些直接模拟的方法可以检验很多科学假设,但这种试验模拟的环境与植物所接触的真正环境是有差异的,所以这种方法仍有一定的局限性,而利用计算机模拟建模,可以提供一些宏观的规律性问题[28-29],但细致的机理性问题的解决仍需要反复试验。
2 克隆植物及芽库的概念
克隆生长是指在自然条件下通过营养生长产生具有潜在独立性的个体的过程[30]。克隆植物的繁殖方式一般分为有性繁殖和无性繁殖两种,对应有种子库和芽库两个概念,在以一年生植物为主的草地或频繁经历扰动的生态系统中,种子库占据主要地位,但在多年生草地生态系统,由于其种子萌发或者幼苗建成困难,且地上部通常只存活一年,而地下的构件可存活多年,所以地上植被的生产力和种群的更新主要依赖于地下芽库而非种子库[31]。芽库的概念最早由Harper[32]于1977年提出,是指与植物多年生器官相联系的分生组织的地下种群,如根茎、球茎、块茎、鳞茎等器官上的芽。而后得到进一步的补充[33-34]。以“bud bank”为检索词,可以查到自2013年以来关于芽库的研究有16 700多篇报道,可见,近几年来研究者对芽库方面的关注已逐渐增加。目前芽库的重要性已经在很多生境中得到证实,例如在北美中部较湿润的草地生态系统,几乎地上所有个体的更新均来自地下芽库而非种子库[35],澳大利亚西南部的很多木本植物也是如此[36]。所以地下芽库的动态与地上生产力密切相关[37],芽库的重要性也逐渐得到体现。

图1 气候变化主要因子对植物光合、生长和无性繁殖的影响Fig. 1 Effect of climate change on photosynthetic rate, growth and propagation of clonal plants
注:红色箭头代表促进作用,蓝色箭头代表抑制作用,黑色箭头代表相关文献研究结果差异较大。
Note: Red, blue and black arrows represent positive, negative and complex influence on plants, respectively.

表1 模拟气候变化的主要处理手段Table 1 Main treatment methods that mimic climatechange
3 模拟温度与CO2增加、氮沉降及降水量变化对多年生克隆植物无性繁殖的影响
3.1 温度对多年生克隆植物无性繁殖的影响
增温对植物的影响一般取决于环境温度是否达到或超过植物生长的最适温度,这对于植物的光合系统等生理指标如此,对于克隆植物的无性繁殖亦是如此。对称和不对称增温对植物生长发育的影响也不完全一致[38-39]。Hoover等[40]发现夏季增温对美国中部草原的影响并不显著,Wang等[41]得出,虽然夏季增温增加了羊草(Leymuschinensis)地下总芽数、根茎数量和长度,但减少了子株的输出。而夜间增温与全天增温有完全不一样的结果,夏季夜间增温促进了羊草的无性繁殖和生物量[18],这支持了Wan等[42]的野外夜间增温研究。这种结果的差异可能是白天增温超过了其适宜的生长温度,造成了胁迫,植株虽然促进了芽的生长但是为规避高温环境而不输出为子株。这也证明了前人提出的对称和不对称增温产生不同结果的观点。此外,不同季节增温以及针对不同物种都可能产生不同的结果,例如,春季增温减少毛竹(Quercuspubescene)的芽和地上生物量[43],但对无芒雀麦(Bromusinermis)的影响较小[44]。因此增温的时间、增温的幅度、作用的物种或群落不同,得到的结果也完全不同。
3.2 CO2浓度增加对多年生克隆植物无性繁殖的影响
单独CO2浓度升高对植物影响的研究也有报道[45-46]。在草原生态系统,CO2浓度的升高均提高了瑞士草原、新西兰草原、堪萨斯高草草原以及科罗拉多矮草草原的生产力[47]。但以往的大部分研究都是CO2浓度升高促进植物的光合作用进而增加植物的地上生物量[47-49],而对于很多克隆植物而言,更合适的对策是通过芽库产生更多的克隆分株来增加地上生产力。例如,CO2浓度的升高促进毒漆藤(Toxiccodendronradican)以及C3草本植物虉草(Phalarisarundinacea)根茎的延长及生物量的累积[50],增强无性繁殖能力,CO2浓度升高也促进芦苇(Phragmitesaustralis)地下芽和子株的生长[51]。很多试验表明,植物生长在温暖地区对CO2的响应要强于较冷地区[52],且在营养受限的条件下CO2的作用才会得到更好的体现,但是这一论题后来被Poorter等[53]推翻,他们公开发表的数据表明CO2在高营养的条件下才可以更好地发挥作用。李卓琳等[54]的研究显示,在低氮条件下CO2浓度升高虽然促进了羊草地下芽的生长,但是并没有促进子株的输出,而只有在高氮条件才显著增加了子株的数量。Pettersson和McDanold[55]也认为CO2浓度升高对植物光合作用的影响取决于氮的供应水平,同时也源于植株整体的源库关系调节。此外,Li等[56]得出CO2在干旱时对羊草生长的促进作用强于降水充沛处理,但这些研究主要是针对植物的生长或生理对CO2浓度升高及其它因子交互作用的响应。关于克隆生长则很少,但无性繁殖的研究在评估未来气候变化对多年生克隆植物生产力等方面有很重要的意义。
3.3 水分对多年生克隆植物无性繁殖的影响
水分是限制草原生产力的最主要因子[57],因此草原生产力不论在自然状态还是在人为干扰的情景下,都受降水梯度变化的极大影响[57-58]。多数研究表明,年平均降水量(MAP)与草地植物地上净初级生产力(ANPP)呈典型的正相关关系,这在近年对我国北方温带草原的74个样地以及内蒙古针茅(Stipacapillata)草原的研究上也取得了同样的结果[59-61]。但在降水量相近的年份之间草原群落生物量差异显著,说明降水量在一年中的分布变化比年降水总量对草原植被的影响更为重要[62],后来Swemmer等[63]进一步研究证实,决定降水分布的3个因素(生长季内的平均单次降水量、降水次数和平均降水间隔时间)对于水分状况不同的草原的影响程度各不相同,对于半干旱区草原来说,生长季内的平均单次降水量和降水次数对ANPP的影响远大于生长季总降水量对ANPP的影响。研究表明,夏季降水量对多年生根茎型禾草羊草的影响要强于降水频次,但在降水量低于目前降水量的40%时,低降水量频次不利于羊草的生长[54]。
和其它植物一样,克隆植物应对水分的变化时通常会发生相应的生长和生理变化,地下芽库也会随着降水的增加而增加。在北美大草原上,地下芽库随着降水梯度变化[64]。但草原的生产力与前一年的降水关系密切,因为翌年春季子株的输出主要源于前一年秋季的芽库储存,所以有研究认为当年的干旱对芽库的影响并不显著[65]。在干旱或者半干旱地区,水分是控制地下芽库的关键因子,且地下芽库对水分的敏感性较强,所以Knapp和Smith[57]认为在沙漠和干旱草原上,低的地上净初级生产力的变化主要是由分生组织限制引起的,阻碍了芽的输出。Reichmann和Sala[66]在对奇瓦瓦沙漠北部刺果垂穗草(Boutelouaeriopoda)的研究中发现,匍匐茎对降水的敏感度要强于茎上的活性芽,且降水量对次年的草地结构组分影响很大,这主要源于匍匐茎密度的改变,而在湿润草原,Koerner等[67]指出,在总降水量不变、单次降水量增大而降水频次降低的条件下,与正常降水频次下相比,草原地下芽库和地上子株并没有显著性变化,所以也并未影响该草原的群落结构和生态系统功能,这暗示着该草原的耐旱弹性较高。而当降水量减少80%时,该草原群落的根茎生物量和地下的芽的密度虽然也能保持稳定,但降低了群落的物种丰富度,其中莎草和非禾本科的地下芽库及地上植株的密度有所减少,禾本科则无显著变化[68]。Ott等[44]通过温室的盆栽试验也得出降水频次的降低对无芒雀麦和蓝茎冰草(Pascopyrumsmithii)的地下芽库影响不显著。
当夏季增温与干旱同时发生时,Hoover等[40]的研究显示增温对该草原的影响并不显著,但极端干旱显著影响该草原群落的物种丰富度和群落组分,且非禾本科植物比禾本科植物更易受影响,这与van der Weide和Hartnett[68]的结果基本一致。在对春季增温和干旱对毛竹(Phyllostachysheterocycla)的盆栽试验中也显示增温和干旱并不存在显著的交互作用[43]。
3.4 氮沉降对多年生克隆植物无性繁殖的影响
在干旱和半干旱地区,氮的输入通常是以脉冲的形式进入土壤[69]。植物所能吸收的氮素为有效氮,而土壤中的氮以有机态和无机态形式存在,植物对养分的吸收和利用都依赖于土壤水分,灌溉能显著提高氮素的固定速率和氮的矿化速率,相反干旱可能会降低氮矿化速率,所以土壤的水分状况在很大程度上决定着肥料的合理利用[70]。绝大部分研究赞同降水量增加与施氮之间的正相关关系,例如,申云霞和唐拴虎对冬小麦(Triticumaestivum)的研究中得出在较低的供水量条件下,要获得理想产量必须降低施肥量,只有当供水量增多时,增加施肥量才能得到较高的产量[71];Asner等[72]也认为氮沉降会增加生态系统的生产力,而在水分不足时,生产力不会随施氮的增加而增加;梁银丽和康绍忠[73]研究施氮在不同土壤水分时对植物抗旱的作用,发现施氮对作物在土壤水分良好、轻度干旱和严重干旱下分别表现出正效应、无明显作用和负效应。
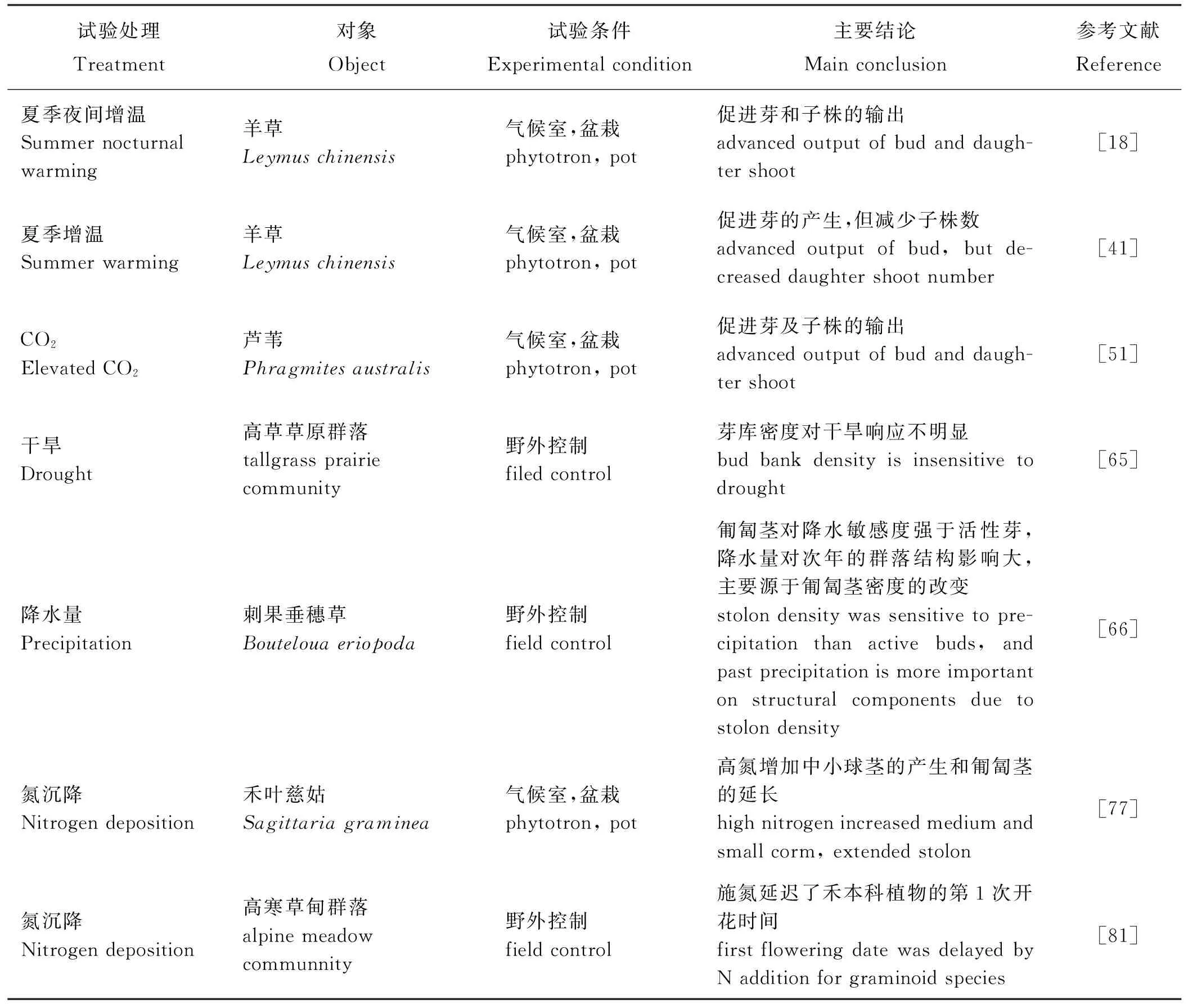
表2 增温、CO2、降水或氮沉降对芽库的影响Table 2 Effects of warming, elevated CO2, precipitation or N deposition on bud bank
此外,施氮对植被生产力的影响取决于植被所处生态系统的氮饱和度。松嫩平原就是一个氮匮乏的地区,羊草的生长通常受土壤中氮的限制,所以施氮能显著增加该平原单一羊草种群的产量和无性繁殖的数量[74]。Tomlinson和Oconnor[75]认为土壤中的N可能是芽休眠或萌发的诱发因素。而氮素营养的添加也可以促进根茎[76]、球茎及匍匐茎的产生[77],扩充芽库;显著增加子株尤其是分蘖子株的数量,因为施氮优先促进植物基部的分蘖节芽形成子株[78]。然而Dalgleish等[79]认为施氮虽然促进了S.heterolepis芽库的扩充以及芽向子株的输出,但对子株个体的大小没有影响。此外,氮素添加对不同类型、不同科的克隆植物也可能产生不同的影响,例如在北美草地,施氮增加了游击型克隆植物的生长而减少了密集型克隆植物的生长[80],这可能导致该地区的密集型克隆植物逐渐被替代;在高寒草甸草原,施氮延迟了禾本科植物的第1次开花时间并减少其繁殖分配,但在非禾本科植物中却产生相反的结果[81]。总体看来,不同物种及不同地区对氮沉降的响应程度不同。在我国东北草甸草原的研究中发现,适度的营养对芽库的建成和萌发是有利的,氮素的供应增加了羊草的无性繁殖能力,提高地上和地下生物量[82]。
表2就增温、CO2升高、氮沉降或降水变化对克隆植物无性繁殖的一些研究结果做了简单总结。但正如前文介绍,氮沉降是伴随降水以脉冲的形式进入土壤,所以氮沉降的作用与降水的关系密不可分,温度虽然可能促进植物的生长但是增温会引发干旱,此外,以CO2为主体的温室效应引发了全球变暖,所以单独研究某一因子的作用并不能很好的预测未来的气候变化产生的影响,现已有一部分气候变化中二因素或三因素交互对植物影响的研究(表3)。
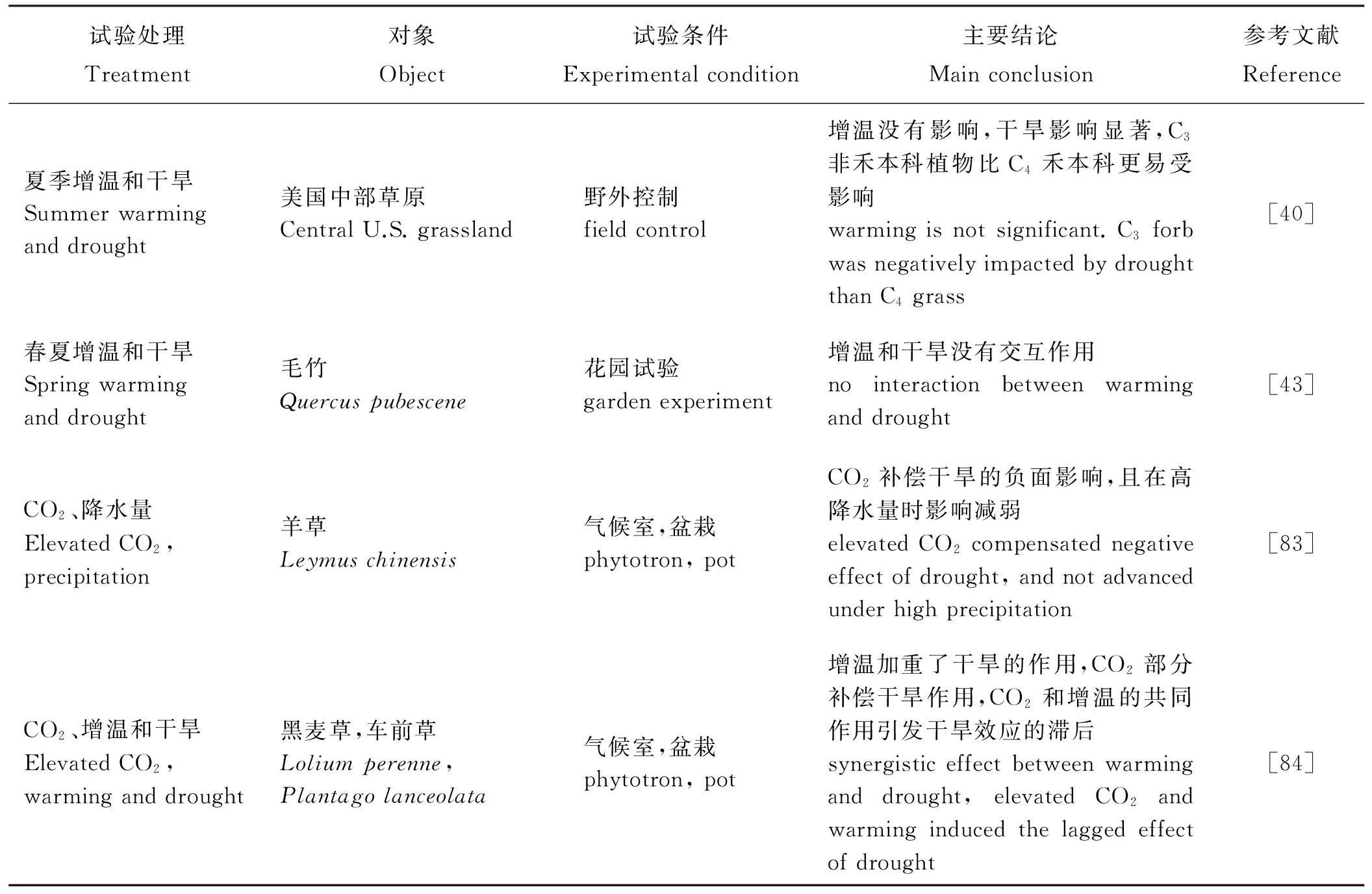
表3 增温、CO2、降水和氮沉降的交互作用对植物种群或群落的影响Table 3 Effects of warming, elevated CO2, rainfall and nitrogen deposition on plant population or community
4 总结
气候变化并不是完全单独直接作用于植物,往往是由于各种微气候因子发生变化的综合作用,它们对植物生长发育、种群动态、群落组分乃至整个生态系统的影响并不是某一单独因子可以决定的[52-55,70-73],正因为各因子之间的交互作用,所以极大地增加了人们预测气候变化背景下植物动态的难度。综合文献发现,关于CO2浓度升高、增温、干旱和氮沉降之间交互作用的研究较少[83],而且可能由于地下取样困难等原因,关于研究CO2浓度升高、增温、氮沉降等气候变化对地下芽库影响的文献更少,仍需要做大量的研究。但实际上绝大部分植物都是克隆植物,尤其是草本植物,无性繁殖是它们的主要繁殖方式,所以对于主要依赖地下芽库完成植被更新的植物而言,关于无性繁殖应对环境变化的研究需要进一步实行,为未来应对气候变化以及草地管理提供一定的科学依据。
References:
[1] 张文军,周启星,魏巍.克隆植物修复退化生态系统的机制.草地学报,2016,24(3):485-490. Zhang W J,Zhou Q X,Wei W.Mechanisms of restoration of degraded ecosystem by clonal plants.Acta Agrestia Sinica,2016,24(3):485-490.(in Chinese)
[2] Houghton J T,Ding Y,Griggs D J,Noguer M.Climate Change 2001:The Scientific Basis.Cambridge:Cambridge University Press,2001.
[3] Liu B,Xu M,Henderson M,Qi Y,Li Y.Taking China’s temperature:Daily range,warming trends,and regional variations,1955-2000.Journal of Climate,2004,17:4453-4462.
[4] Easterling D R,Horton B,Jones P D,Peterson T C,Karl T R,Parker D E,Salinger M J,Razuvayev V,Plummer N,Jamson P,Folland C K.Maximum and minimum temperature trends for the globe.Science,1997,277:364-367.
[5] Alward R D,Detling J K,Milchunas D G.Grassland vegetation changes and nocturnal global warming.Science,1999,283:229-231.
[6] Solomon S D,Qin D,Manning M,Chen Z,Marquis M.Climate Change 2007:The Physical Science Basis.Working Group Ⅰ Contribution to the Fourth Assessment Report of the IPCC.Computational Geometry,2007,18(2):95-123.
[7] Min S K,Zhang X,Zwiers F W,Hegeri G C.Human contribution to more-intense precipitation extremes.Nature,2011,470:378-381.
[8] Smith M D.An ecological perspective on extreme climatic events:A synthetic definition and framework to guide future research.Journal of Ecology,2011,99:656-663.
[9] 李新周,刘晓东,马柱国.近百年来全球主要干旱区的干旱化特征分析.干旱区研究,2004,21(2):97-103. Li X Z,Liu X D,Ma Z G.Analysis on the drought characteristics in the main arid regions in the world since recent hundred-odd years.Arid Zone Research,2004,21(2):97-103.(in Chinese)
[10] Hooper D U,Johnson L.Nitrogen limitation in dryland ecosystems:Responses to geographical and temporal variation in precipitation.Biogeochemistry,1999,46:247-293.
[11] Holland E A,Dentener F J,Braswell B H,Sulzman J M.Contemporary and pre-industrial global reactive nitrogen budgets,in new perspectives on nitrogen cycling in the temperate and tropical americas.Biogeochemistry,1999,46:7-43.
[12] Allen E B,Rao L E,Steers R J,Bytnerowicz A,Fenn M E.Impacts of atmospheric nitrogen deposition on vegetation and soils in joshua tree national park.Las Vegas,Nevada,USA:University of Nevada Press,2009:78-100.
[13] Rao L E,Parker D R,Bytnerowicz A,Allen E B.Nitrogen mineralization across an atmospheric nitrogen deposition gradient in southern california deserts.Journal of Arid Environments,2009,73:920-930.
[14] 武倩,韩国栋,王瑞珍,刘芳,秦洁.模拟增温对草地植物、土壤和生态系统碳交换的影响.中国草地学报,2016,38(4):105-114. Wu Q,Han G D,Wang R Z,Liu F,Qin J.Effects of simulated warming on grassland plants,soil and ecosystem carbon exchange.Chinese Journal of Grassland,2016,38(4):105-114.(in Chinese)
[15] 曹路,李春瑞,田青松,杜建材,王忠武,韩冰.内蒙古荒漠草原植物遗传多样性对模拟增温处理的响应.生态学报,2016,36(21):6909-6918. Cao L,Li C R,Tian Q S,Du J C,Wang Z W,Han B.The response of genetic diversity in desert steppe plants to simulated warming in Inner Mongolia,China.Acta Ecologica Sinica,2016,36(21):6909-6918.(in Chinese)
[16] Cowles J M,Wragg P D,Wright A J,Powers J S,Tilman D.Shifting grassland plant community structure drives positive interactive effects of warming and diversity on aboveground net primary productivity.Global Change Biology,2016,22(2):741-749.
[17] Gao S,Wang J F,Zhang Z J,Dong G,Guo J X.Seed production,mass,germinability,and subsequent seedling growth responses to parental warming environment inLeymuschinensis.Crop & Pasture Science,2012,63:87-94.
[18] Li Z L,Lin J X,Zhang T Y,Zhang N,Mu C S,Wang J F.Effects of summer nocturnal warming on biomass production ofLeymuschinensisin the songnen grassland of China:From bud bank and photosynthetic compensation.Journal of Agronomy and Crop Science,2014,200:66-76.
[19] Kuster T M,WilkinsonA,Hill P W,Jones D L,Bardgett R D.Warming alters competition for organic and inorganic nitrogen between co-existing grassland plant species.Plant and Soil,2016,406(1):117-129.
[20] 付刚,孙维,李少伟,何永涛,沈振西.藏北高原不同海拔高寒草甸群落地上部分碳氮含量对模拟增温的响应.生态环境学报,2015,24(7):1093-1097. Fu G,Sun W,Li S W,He Y T,Shen Z X.Response of community aboveground parts carbon and nitrogen content to experimental warming in an alpine meadow at three elevations in the northern Tibet.Ecology and Environment Sciences,2015,24(7):1093-1097. (in Chinese)
[21] Madhu M,Hatfield J L.Dynamics of plant root growth under increased atmospheric carbon dioxide.Agronomy Journal,2013,105(3):657.
[22] de Souza A P,Arundale R A,Dohleman F G,Long S P,Buckeridge M S.Will the exceptional productivity ofMiscanthusxgiganteusincrease further under rising atmospheric CO2.Agricultural and Forest Meteorology,2013,171:82-92.
[23] Leakedy A D,Bishop K A,Ainsworth E A.A multi-biome gap in understanding of crop and ecosystem responses to elevatd CO2.Current Opinion in Plant Biology,2012,15:228-236.
[24] Zhang B C,Cao J J,Bai Y F,Zhou X,Ning Z,Yang S,Hu L.Effects of rainfall amount and frequency on vegetation growth in a Tibetan alpine meadow.Climate Change,2013,118:197-212.
[25] Padilla F M,Aarts B H J,Roijendijk Y O A,Caluwe H D,Mommer L,Visser E J W.Root plasticity maintains growth of temperate grassland species under pulsed water supply.Plant and Soil,2013,369:1-10.
[26] Cleland E E,Collins S L,Dickson T L,Farre E C,Gross K L,Gherardi L A,Hallett L M,Hobbs R J,Hsu J S,Turnbull L,Suding K N.Sensitivity of grassland plant community composition to spatial vs. temporal variation in precipitation.Ecology,2013,94(8):1687-1696.
[27] Gan J S,Esmaile M M,Sattarian A.Role of soil fertilization and cutting on tillering and yield components ofSorghumhalepense.International Journal of Agronomy and Plant Production,2013,4(5):994-997.
[28] Luo Y Q,Hui D F,Zhang D Q.Elevated CO2stimulates net accumulations of carbon and nitrogen in land ecosystems:A meta-analysis.Ecology,2013,87(1):53-63.
[29] Ainsworth E A,Long S P.What have we learned from 15 years of free-air CO2enrichment (face) a meta-analytic review of the responses of photosynthesis,canopy properties and plant production to rising CO2.New Phytologist,2005,165:351-372.
[30] 董鸣.克隆植物生态学.科学出版社,2011,7.
[31] Benson E J,Hartnett D C,Mann K H.Belowground bud banks and meristem limitation in tallgrass prairieplant populations.American Journal of Botany,2004,91:416-421.
[32] Harper J L.Population Biology of Plants.New York,USA:Academic Press,1977.
[33] Klimeš L.Bud banks and their role in vegetative regeneration——A literature review and proposal for simple classification and assessment.Perspectives in Plant Ecology,Evolution and Systematics,2007,8(3):115-129.
[34] 赵凌平,王占彬,程积民.草地生态系统芽库研究进展.草业学报,2015,24(7):172-179. Zhao L P,Wang Z B,Cheng J M.Review of bud banks in grassland ecosystems.Acta Prataculturae Sinica,2015,24(7):172-179.(in Chinese)
[35] Benson E J,Hartnett D C.The role of seed and vegetative reproduction in plant recruitment and demography in tallgrass prairie.Plant Ecology,2006,187:163-178.
[36] Meney K,Dixon K,Pate J.Reproductive potential of obligate seeder and resprouter herbaceous perennial monocots (Restionaceae,Anarthriaceae,Ecdeiocoleaceae) from South-western Western Australia.Australian Journal of Botany,1997,45:771-782.
[37] Ni J.Estimating net primary productivity of grasslands from field biomass measurements in temperate northern China.Plant Ecology,2004,174(2):217-234.
[38] Bunce J.Acclimation of photosynthesis to temperature inArabidopsisthalianaandBrassicaoleracea.Photosynthetica,2008,46:517-524.
[39] Kim S H,Gitz D C,Sicher R C,Baker J T,Timlin D J,Reddy V R.Temperature dependence of growth,development,and photosynthesis in maize under elevated CO2.Environmental and Experimental Botany,2007,61:224-236.
[40] Hoover D L,Knapp A K,Smith M D.Resistance and resilience of a grassland ecosystem to climate extremes.Ecology,2014,95(5):2646-2656.
[41] Wang J F,Gao S,Lin J,Mu Y G,Mu C S.Summer warming effects on biomass production and clonal growth ofLeymuschinensis.Crop and Pasture Science,2010,61:670-676.
[42] Wan S,Xia J,Liu W,Niu S.Photosynthetic overcompensation under nocturnal warming enhances grassland carbon sequestration.Ecology,2009,90:2700-2710.
[43] Wellstein C,Cianfaglione K.Impact of extreme drought and warming on survival and growth characteristics of different provenences of juvenileQuercuspubesceneWilld.Folia Geobotanica,2014,49:31-47.
[44] Ott J P,Butler J L,Rong Y P,Xu L.Greater bud outgrowth ofBromusinermisthanPascopyrumsmithiiunder multiple environmental conditions.Journal of Plant Ecology,2016:DOI:10.1093/jpe/rtw045.
[45] Long S P,Ainsworth E A,Rogers A,Ort D R.Rising atmospheric carbon dioxide:Plants face the future.Annual Review Plant Biology,2004,55:591-628.
[46] Luo Y,Hui D,Zhang D.Elevated CO2stimulates net accumulations of carbon and nitrogen in land ecosystems:A meta-analysis.Ecology,2006,87:53-63.
[47] Morgan J A,Lecain D R,Mosier A R,Milchunas D G.Elevated CO2enhances water relations and productivity and affects gas exchange in C3and C4grasses of the Colorado shortgrass steppe.Global Change Biology,2001,7:451-466.
[48] Kakani V,Reddy K R.Temperature response of C4species big bluestem is modified by growing carbon dioxide concentration.Environmental and Experimental Botany,2007,61:281-290.
[49] Tobita H,Uemura A,Kitao M,Kitaoka S,Utsugi H.Interactive effects of elevated CO2,phosphorus deficiency,and soil drought on nodulation and nitrogenase activity inAlnushirsutaandAlnusmaximowiczii.Symbiosis,2010,50:59-69.
[50] Kinmonth S H, Kim S H. Carbon gain, allocation and storage in rhizomes in response to elevated atmospheric carbon dioxide and nutrient supply in a perennial C3grass, phalaris arundinacea. Functional Plant Biology,2011,38:797-807.
[51] Zhang N,Lin J X,Yang Y H,Li Z L,Wang Y,Cheng L Y,Shi Y J,Zhang Y T,Wang J F,Mu C S.The tolerance of growth and clonal propagation ofPhragmitesaustralis(common reeds) subjected to lead contamination under elevated CO2conditions.RSC Advances,2015,5:55527-55535.
[52] Mooney H,Drake B G,Luxmoore R,Luxmoore R J,Oechel W C,Pitelka L F.Predicting ecosystem responses to elevated CO2concentrations.Bioscience,1991,41(2):96-104.
[53] Poorter H.Do slow-growing species and nutrient-stressed plants respond relatively strongly to elevated CO2.Global Change Biology,1998,4:693-697.
[54] 李卓琳.羊草生长发育对模拟主要全球气候变化因子的响应.长春:东北师范大学博士学位论文,2014. Li Z L.Study on growth and development response ofLeymuschinensisto main global climate change factors.PhD Thesis.Changchun:Northeast Normal University,2014.(in Chinese)
[55] Pettersson R,McDonald A J S.Effects of nitrogen supply on the acclimation of photosynthesis to elevated CO2.Photosynthesis Research,1994,39:389-400.
[56] Li Z L,Zhang Y T,Yu D F,Zhang N,Lin J X,Zhang J W,Tang J H,Wang J F,Mu C S.The influence of precipitation regimes and elevated CO2on photosynthesis and biomass accumulation and partitioning in seedlings of the rhizomatous perennial grassLeymuschinensis.PLoS One,2014,9(8):e103633.
[57] Knapp A K,Smith M D.Variation among biomes in temporal dynamics of aboveground primary production.Science,2001,291:481-484.
[58] Bai Y,Wu J,Xing Q,Pan Q,Huang J,Yang D,Han X.Primary production and rain use efficiency across a precipitation gradient on the Mongolia Plateau.Ecology,2008,89:2140-2153.
[59] Bai Y,Han X,Wu J,Chen Z,Li L.Ecosystems ability and compensatory effects in the Inner Mongolia grassland.Nature,2004,431:181-184.
[60] 韩国栋.降水量和气温对小针茅(StipaklemenziiRoshev.)草原植物群落初级生产力的影响.内蒙古大学学报:自然科学版,2002,33(1):83-88. Han G D.Influence of precipitation and air temperature on primary productivity ofStipaklemenziiplant community,Nei Mongol.Acta Scientiarum Naturalium Univeristatis Neimongal,2002,33(1):83-88.(in Chinese)
[61] 蒋德明,周全来,阿拉木萨,唐毅,押田敏雄.科尔沁沙地植被生产力对模拟增加降水和氮沉降的响应.生态学杂志,2011,30(6):1070-1074. Jiang D M,Zhou Q L,Alamusa,Tang Y,Toshi O.Responses of vegetation productivity in Horqin sand land to simulated increased precipitation and nitrogen deposition.Chinese Journal of Ecology,2011,30(6):1070-1074.(in Chinese)
[62] Knapp A K,Fay P A,Blair J M,Collins S L,Smith M D,Carlisle J D,Harper C W,Danner B T,Lett M S,Mccarron J K.Rainfall variability,carbon cycling,and plant species diversity in a mesic grassland.Science,2002,298:2202-2205.
[63] Swemmer A M,Knapp A K,Snyman H A.Intra-seasonal precipitation patterns and above-ground productivity in three perennial grasslands.Journal of Ecology,2007,95:780-788.
[64] Niu S,Yang H,Zhang Z,Wu M,Lu Q,Li L,Han X.Non-additive effects of water and nitrogen addition on ecosystem carbon exchange in a temperate steppe.Ecosystems,2009,12:915-926.
[65] vander Weide B L,Hartnett D C,Carter D L.Belowground bud banks of tallgrass prairie are insensitive to multi-year,growing-season drought.Ecosphere,2014,5(8):1-17.
[66] Relchmann L G,Sala O E.Differential sensitivities of grassland structural components to changes in precipitation mediate productivity response in a desert ecosystem.Functional Ecology,2014,28:1292-1298.
[67] Koerner S E,Collins S L,Blair J M,Knapp A K,Smith M D.Rainfall variability has minimal effects on grassland recovery from repeated grazing.Journal of Vegetation Science,2014,25:36-44.
[68] vander Weide B L,Hartnett D C.Belowground bud bank response to grazing under severe,short-term drought.Oecologia,2015,178:795-806.
[69] 周晓兵,张元明.干旱半干旱区氮沉降生态效应研究进展.生态学报,2009,29(7):3835-3845. Zhou X B,Zhang Y M.Review on the ecological effects of N deposition in arid and semi-arid areas.Acta Ecologica Sinica,2009,29(7):3835-3845.(in Chinese)
[70] 李世娟,周殿玺.限水灌溉下不同氮肥用量对小麦产量及氮素分配利用的影响.华北农学报,2001,16:86-91. Li S J,Zhou D X.Effect of different nitrogen application on yield,nitrogen distribution and utilization in winter wheat under soil water stress.Acta Agricultural Boreali-Sinica,2001,16:86-91.(in Chinese)
[71] 申云霞,唐拴虎.冬小麦水肥产量交互效应模拟研究.西北植物学报,1995,15:138-141.
[72] Asner G P,Townsend A R,Riley W J,Maston P A,Neff J C.Physical and biogeochemical controls over terrestrial ecosystem responses to nitrogen deposition.Biogeochemistry,2001,54:1-39.
[73] 梁银丽,康绍忠.限量灌水和磷营养对冬小麦产量及水分利用的影响.土壤侵蚀与水土保持学报,1997(3):60-67.
[74] 杨允菲,张宝田.松嫩平原羊草种群营养繁殖的季节动态及其生物量与密度关系的分析.植物学报,1992,34(6):443-449. Yang Y F,Zhang B T.An analysis of seasonal variation of vegetative propagation and the relationships between biomass and population density ofAneurolepidiumchinensein Songnen Plain of China.Acta Botanica Sinica,1992,34(6):443-449.(in Chinese)
[75] Tomlinson K,Oconnor T.Control of tiller recruitment in bunchgrasses:Uniting physiology and ecology.Functional Ecology,2004,18:489-496.
[76] Hutchings M,Kroon H D.Foraging in plants:The role of morphological plasticity in resource acquisition.Advances in Ecological Research,1994,25:159-238.
[77] Zhang L H,Zhang Y W,Zhao X N,Huang S J,Zhao J M,Yang Y F.Effects of different nutrient sources on plasticity of reproductive strategies in a monoecious species,Sagittariagraminea(Alismataceae).Journal of Systematics and Evolution,2014,52(1):84-91.
[78] Bai W,Sun X,Wang Z,Li L.Nitrogen addition and rhizome severing modify clonal growth and reproductive modes ofLeymuschinensispopulation.Plant Ecology,2009,205:13-21.
[79] Dalgleish H J,Kula A R,Hartnett D C,Sandercock B K.Responses of two bunchgrasses to nitrogen addition in tallgrass prairie:The role of bud bank demography.American Journal of Botany,2008,95:672-680.
[80] Gough L,Gross K L,Cleland E E,Clark C M,Collins S L.Incorporating clonal growth form clarifies the role of plant height in response to nitrogen addition.Oecologia,2012,169:1053-1062.
[81] Zhang Z L,Niu K C,Liu X D,Jia P,Du G.Linking flowering and reproductive allocation in response to nitrogen addition in an alpine meadow.Journal of Plant Ecology,2014,7(3):231-239.
[82] 王俊锋.氮,水和温度对羊草有性生殖及克隆生长的影响.长春:东北师范大学博士学位论文,2010. Wang J F.Sexual reproduction and clonal growth ofLeymuschinensisin response to nitrogen application,waters,and warming.PhD Thesis.Changchun:Northeast Normal University,2010.(in Chinese)
[83] Li Z L,Zhang Y T,Yu D F,Zhang N,Lin J X,Zhang J W,Tang J H,Wang J F,Mu C S.The influence of precipitation regimes and elebated CO2on photosynthesis and biomass accumulation and partitioning in seedling of the rhizomatous perennial grassLeymuschinensis.PLoS One,2014,9(8):e103633.
[84] vande Velde H,Bonte D,AbdElgawad H,Asard H,Nijs I.Combined elevated CO2and climate warming induces lagged effects of drought inLoliumperenneandPlantagolanceolata.Plant Ecology,2015,216:1047-1059.
(责任编辑 王芳)
Review of the effects of climate change on the propagation of perennial clonal plants
Li Zhuo-lin1, Mu Chun-sheng2, Wu Chun-ying3
(1.Key Laboratory of Urbanization and Ecological Restoration of Shanghai, School of Ecological and Environmental Science, East China Normal University, Shanghai 200241, China; 2.Key Laboratory of Vegetation Ecology, Ministry of Education, Institute of Grassland Science, Northeast Normal University, Changchun 130024, China; 3.Grassland Management Station of Yanbian Autonomous Prefecture, Yanji 133001, China)
Climate change consists of many simultaneous factors which have different influences on plants, animals and microbes in various ecosystems. Clonal plants have two different modes of reproduction, sexual reproduction and vegetative propagation. Many clonal plants have abandoned or nearly abandoned sexual reproduction, owing to environmental adaptation. To some extent, their vegetative propagation ability might represent the total productivity of these plants. Therefore, the response of clonal plant propagation to climate change has becomea crucial issue. In this paper, we review the effects of the main climatic factors on clonal growth of a perennial plant. Both elevated CO2and nitrogen deposition increased the output of daughter shoots in most papers, but decreasing precipitation had adverse or noeffect. Warming time had different effects on daughter shoots. Although there is a lot of research about the effects of eachclimate change factor on clonal plants, studies showing howthe interaction of these factors affectsclonal plant propagation arescarce.To strengthen the body of research, studies about how interaction among the climatic factors affectsclonal plants areessential, because theynot only enrich knowledge of clonal plant and global change ecology, but also provide practical guidance for grassland management in the future.
CO2; warming; precipation pattern; N deposition; clonal growth
Mu Chun-sheng E-mail:mucs821@nenu.edu.cn
10.11829/j.issn.1001-0629.2016-0414
2016-08-08 接受日期:2017-03-24
973课题(2015CB150801);国家自然科学基金(31370432);植被生态科学教育部重点实验室开放课题编号(130028675)
李卓琳(1987-),女,黑龙江漠河人,博士,主要从事全球气候变化和牧草繁殖的研究。E-mail:lizhuolin1987621@126.com
穆春生(1961-),男,吉林长春人,教授,博士,主要从事饲料作物生态学和草业科学方向的研究。E-mail:mucs821@nenu.edu.cn
S162.5+9;Q785
A
1001-0629(2017)08-1694-11
李卓琳,穆春生,吴春英.气候变化对草原多年生克隆植物无性繁殖的影响.草业科学,2017,34(8):1694-1704.
Li Z L,Mu C S,Wu C Y.Review of the effects of climate change on the propagation of perennial clonal plants.Pratacultural Science,2017,34(8):1694-1704.
猜你喜欢
广东气象(2022年5期)2022-10-26
环球时报(2022-09-20)2022-09-20
成都信息工程大学学报(2022年3期)2022-07-21
资源节约与环保(2022年12期)2022-02-02
今日农业(2020年24期)2020-12-15
英语文摘(2020年4期)2020-07-28
英语文摘(2020年4期)2020-07-28
启蒙(3-7岁)(2019年8期)2019-09-10
江西农业(2018年23期)2018-02-11
小资CHIC!ELEGANCE(2015年15期)2015-09-01
