
异花柽柳种子萌发对温周期和盐分的响应
2017-09-08 03:40王喜勇蔡丹红
草业科学 2017年8期
王喜勇,蔡丹红,魏 岩
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐830052; 2.中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011; 3.浙江省温岭市松门中学,浙江 温岭 317500)
异花柽柳种子萌发对温周期和盐分的响应
王喜勇1,2,蔡丹红1,3,魏 岩1
(1.新疆农业大学草业与环境科学学院,新疆 乌鲁木齐830052; 2.中国科学院新疆生态与地理研究所,新疆 乌鲁木齐 830011; 3.浙江省温岭市松门中学,浙江 温岭 317500)
以新疆吐鲁番沙漠植物园的异花柽柳(Tamarixgracilis)为研究材料,设置4个温周期、10个NaCl溶液浓度梯度,在实验室条件下研究了温周期和盐分对异花柽柳种子萌发的影响。结果显示,1)异花柽柳种子在5 ℃/15 ℃、10 ℃/20 ℃、15 ℃/25 ℃、20 ℃/30 ℃温周期下均快速萌发,温度越高,萌发速度越快。2)异花柽柳种子在低于0.4 mol·L-1的NaCl溶液处理下萌发率均高,且处理之间无显著差异(P>0.05);而高于0.6 mol·L-1的处理则显著抑制种子的萌发(P<0.05),且NaCl溶液的浓度越高,最终萌发率越低,大于1.6 mol·L-1的溶液处理中未见种子萌发。3)异花柽柳种子在解除盐分胁迫后恢复萌发率低。0.6~1.2 mol·L-1的NaCl溶液处理下未萌发的种子恢复萌发率在2.1%~34.5%,大于1.2 mol·L-1的NaCl溶液处理后种子未见萌发,表明1.2 mol ·L-1NaCl为异花柽柳种子的最大耐盐浓度。综上,异花柽柳种子适宜萌发的温周期范围宽,种子萌发耐盐能力属于中耐盐性,耐盐机制为耐盐型。
新疆;异花柽柳;温周期;盐胁迫;萌发特性;盐渍环境
柽柳属(Tamarix)植物主要分布于亚洲和北非[1],在我国主要分布于西北、内蒙古和华北地区[2]。柽柳属植物具有盐腺,属典型的泌盐植物[3],能适应土壤盐渍化环境,具有较强的生态适应性[4],也是盐渍化沙地和重盐碱地主要造林绿化的优良树种。异花柽柳(Tamarixgracilis)抗寒喜冷,一般生于荒漠和干旱草原地区盐碱化土地、沙地、沙丘边缘,河湖沿岸等处。在我国分布于新疆、柴达木盆地、甘肃以及内蒙古西部地区;在中亚、蒙古也有分布[2-5]。异花柽柳为灌木,高1.5~4.0 m,花大而美丽,是适于较寒地区种植的固沙造林树种[2]。
种子萌发是生活史的重要阶段,也是柽柳种群更新的重要途径之一[2,6]。植物种子萌发信息对于预测野外植物群落动态和植被管理十分重要[7]。柽柳属植物种子有不同的萌发对策,例如刚毛柽柳(T.hispida)和多花柽柳(T.hohenackeri)种子萌发温周期适应幅度宽,萌发速度快,在高浓度盐分溶液处理后刚毛柽柳种子萌发有较高的恢复萌发率[8],而多花柽柳恢复萌发率低[9]。国内外有关异花柽柳的研究主要集中在分类分布[1,2-5]、形态结构[10-11]等方面,对种子萌发特性尚未见报道。那么温变周期对异花柽柳种子的萌发有何影响? 异花柽柳种子对不同盐浓度有何响应?本研究在室内研究种子在不同温周期和盐分下种子的萌发特性,旨为揭示异花柽柳种子的萌发对策以及种苗更新提供基础资料。
1 材料与方法
1.1 材料
异花柽柳果实(蒴果)于2014年5月(种子成熟期)在吐鲁番沙漠植物园采集。果实收集后,在通风条件下晾干,待果实全部裂开后挑选饱满的种子用于试验。种子的千粒重为(25.9±5.2) mg,长度为(0.68±0.06) mm,宽度为(0.37±0.04) mm。
1.2 研究方法
1.2.1 萌发试验 挑选的种子置于垫有两层滤纸的培养皿中,在光照培养箱(GTOP-158Y)中模拟自然温度和光照进行种子的萌发试验。每培养皿中放置50粒种子,设置4个重复。在种子萌发过程中,第1天每2 h观测1次,之后每24 h观测1次,持续观察直到连续2 d无新种子萌发时视为萌发结束。
1.2.2 种子在不同温周期下的萌发 试验设置5 ℃/15 ℃、10 ℃/20 ℃、15 ℃/25 ℃、20 ℃/30 ℃(暗/光=12 h/12 h)4个温周期[10-11],分别进行种子萌发试验,并记录不同温周期下种子萌发率达到50%的时间(TG50)。
1.2.3 种子在不同浓度NaCl溶液中的萌发 试验设置10个NaCl溶液的浓度梯度,分别为0(CK)、0.05、0.10、0.20、0.40、0.60、0.80、1.00、1.20、1.60 mol·L-1。依据种子在不同温周期下的萌发结果,种子在不同浓度NaCl溶液中的萌发试验在15 ℃/25 ℃的温度和暗/光12 h/12 h条件下进行;萌发结束后将未萌发的种子用水清洗后转移到蒸馏水中继续恢复萌发。
试验结束后,计算种子在盐溶液中的萌发率(G1),水清洗后种子的恢复萌发率(G2),最终萌发率(G3),计算公式:G1=B/C×100%;G2=(A-B)/(C-B)×100%;G3=A/C×100%。式中:A是试验中全部萌发种子数量,B是盐溶液处理下的萌发种子数量,C是该处理参试种子数量。
1.3 数据分析
使用SPSS 17.0统计软件,在95%水平上对不同温周期和不同浓度NaCl溶液中的萌发结果进行单因子方差分析。采用Duncan’s new multiple range test检验不同浓度NaCl溶液处理间的差异显著性(P<0.05)。用Excel软件制图。
2 结果与分析
2.1 种子在不同温周期下的萌发
异花柽柳种子萌发的适宜温周期范围宽,种子萌发速度快,且最终萌发率高(图1)。4个温周期的萌发时间均小于6d,萌发速率则随着温周期的温度升高而增快,在20 ℃/30 ℃萌发率达到50%时仅需4 h(图2)。

图1 在不同温周期下异花柽柳种子的萌发率Fig. 1 Germination percentage of T. gracilis seeds at different thermoperiods
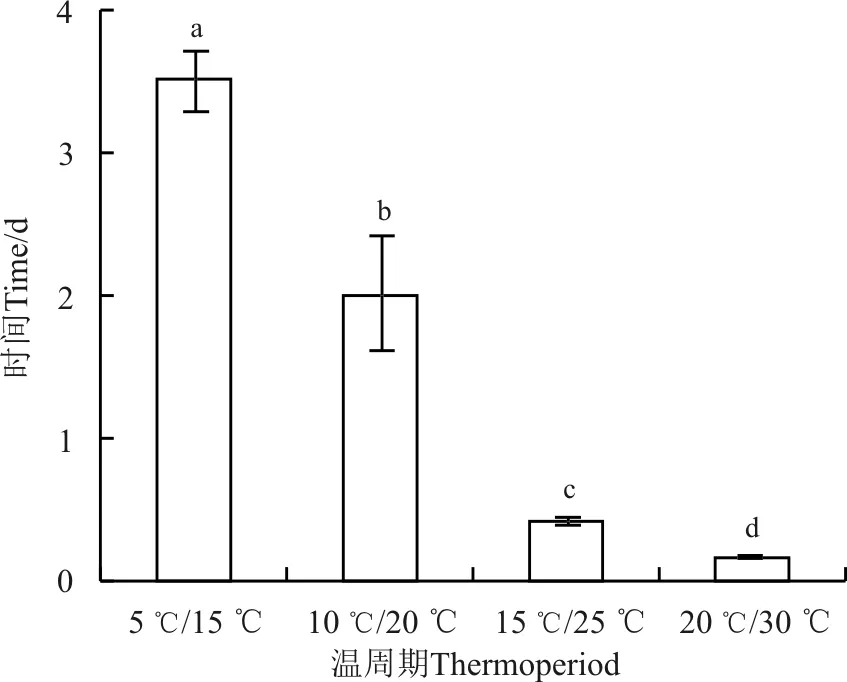
图2 温周期对异花柽柳种子萌发率达到50%所需时间(TG50)的影响Fig. 2 Effect of thermoperiods on germination time of the mean germination percentages reaching 50% ( TG50 ) of T. gracilis seeds
注:不同小写字母表示不同温周期处理间差异显著(P<0.05)。
Note: Different lowercase letters indicate significant difference among different thermoperiods at the 0.05 level.
2.2 NaCl溶液对种子萌发的影响
2.2.1 种子萌发 种子在不同NaCl溶液处理下的萌发结果显示(图3),低于0.40 mol·L-1的NaCl溶液对种子的最终萌发率无明显影响,但随着盐浓度的增加,种子的初始萌发时间明显延长。而高于0.60 mol·L-1的溶液则抑制种子的萌发,且随着浓度的增加,种子的最终萌发率呈下降趋势,在NaCl溶液浓度增加至1.20 mol·L-1时,种子的萌发几乎被完全抑制。
2.2.2 复水后的异花柽柳种子的恢复萌发 在不同浓度NaCl溶液中培养14 d后,将未萌发的种子转移到蒸馏水中,种子的恢复萌发率保持在较低的水平,并且随着盐浓度的不断增加,种子的恢复萌发率呈显著下降趋势(P<0.05)。在1.20 mol·L-1盐溶液的恢复萌发率仅为2.10%,最终萌发率仅为4.00%(表1)。异花柽柳的最终萌发率也随盐浓度的增加而显著降低,直至为零。表明高浓度盐溶液对种子有永久性抑制作用。

图3 异花柽柳种子在不同浓度NaCl溶液处理下的萌发率Fig. 3 Germination percentages of T. gracilis seeds at different concentrations of NaCl solution
注:不同小写字母表示不同NaCl处理下异花树柳种子的最终萌发率差异显著(P<0.05)。
Note: Different lowercase letters indicate significant difference of fial germination percentage among different NaCl treatmeats at the 0.05 level.
3 讨论与结论
温度是决定种子萌发的主要条件之一,它不但影响种子的萌发率,也影响种子的萌发速率[7]。本研究结果中,异花柽柳在5 ℃/30 ℃不同温周期条件下种子的最终萌发率都高达100%,表明种子萌发的适宜温周期范围宽;其种子的萌发速率快,温度越高萌发速度越快,在高温周期萌发率达到50%仅需4 h,完成全部发芽过程仅需1 d。同属植物多枝柽柳[12]和刚毛柽柳[8]、密花柽柳(Tamarixarceuthoides)[13]种子对温度的适应幅度也较宽,在10~30 ℃恒温和变温下均具有快速萌发的特性。柽柳属植物种子对温度的适应幅度较宽,并且遇水萌发速度快,具有机会主义萌发对策。

表1 异花柽柳种子在不同浓度的NaCl溶液中的萌发率、恢复萌发率和最终萌发率Table 1 Initial germination percentages, recovery percentages and final germination percentages of T. gracilis seeds at different concentrations of NaCl solution
注:同列不同小写字母表示不同浓度的NaCl溶液处理间差异显著(P<0.05)。
Note: Different lowercase letters within the same column indicate significant differences among different concentrations of NaCl solution at the 0.05 level.
盐渍环境中过多的盐分限制了绝大多数植物的生长[3]。而盐生植物最大的特点就是对盐渍生境有较强的适应能力,能够在盐渍环境中正常萌发、生长并完成生活史[14-16]。长期生活在盐渍环境具有一定的耐盐能力,但植物种类不同,其种子萌发对盐分的响应也不同[14,17-18]。低浓度盐分(≤0.05 mol·L-1NaCl)对异花柽柳种子的萌发无促进作用。但低浓度盐分(≤0.05 mol·L-1NaCl)对同属植物刚毛柽柳[8]种子的萌发有明显的促进作用。本研究表明,小于0.40 mol·L-1的NaCl溶液对异花柽柳种子的最终萌发率无显著影响,之后随着盐分的增加种子的萌发率迅速降低。依据Khan[19]对盐生植物种子在萌发期间的耐盐性分为低耐盐、中耐盐和高耐盐3类的划分标准,异花柽柳种子属于中耐盐类种子。
大部分盐生植物种子在高浓度盐分条件下长时间具有保持活力的能力,且在盐分浓度降低后能恢复萌发[15,20-21],而有的盐生植物种子在高浓度盐分胁迫后丧失萌发能力[12]。Zhang等[22]对12种盐生植物种子萌发和早期幼苗的耐盐性进行研究,将盐生植物的耐盐机制分为耐盐型和避盐型。本研究中,异花柽柳种子在盐分浓度降低后恢复萌发率很低(<35%),在1.2 mol·L-1的NaCl溶液胁迫后恢复萌发率仅2.10%,表明在NaCl处理中大部分未萌发种子丧失了活力,1.2 mol·L-1NaCl为异花柽柳种子的最大耐盐浓度,其耐盐机制属于耐盐型。同属的多枝柽柳[12]、多花柽柳[9]也有相似的现象。这也说明NaCl对种子萌发的抑制作用既表现为渗透胁迫,也存在一定程度的离子毒害[3,15]。
盐渍环境中,种子能否正常萌发对该物种在某一地区的定居至关重要[3,15]。异花柽柳种子小,顶端具毛,可飘浮至远处进行扩散;种子具有遇水快速萌发的机会主义萌发策略,并且属于中耐盐类种子,这些萌发特性可使有限的萌发单元尽可能占据更大的空间,可能是柽柳作为荒漠地带隐域性植被长期对盐渍环境适应的结果。
References:
[1] 张鹏云,张耀甲.中国植物志第50卷第2分册.北京:科学出版社,1990:142-177.
[2] 刘铭庭.柽柳属植物综合研究及大面积推广应用.兰州:兰州大学出版社,1995. Liu M T.TamarixPlant Utilization Research and Large Scale Applications.Lanzhou:Lanzhou University Press,1995.(in Chinese)
[3] 郗金标,张福锁,田长彦.新疆盐生植物.北京:科学出版社,2006:82. Hao J B,Zhang F S,Tian C Y.Halophyte of Xinjiang.Beijing:Science Press,2006:82.(in Chinese)
[4] 尹林克.柽柳属植物的生态适应性与引种.干旱区研究,2002,19(9):12-16. Yin L K.The ex-situ protection and the ecological adaptability ofTamarixL. in China.Arid Zone Research,2002,19(9):12-16.(in Chinese)
[5] 尹林克.中亚荒漠生态系统中的关键种——柽柳.干旱区研究,1995,12(3):43-47. Yin L K.Tamarix,the key species of desert ecosystem.Arid Zone Research,1995,12(3):43-47.(in Chinese)
[6] Gutterman Y.Seed Germination in Desert Plants Adaptations of Desert Organisms.Berlin:Springer-Verlag,1993.
[7] Grime J P,Mason G,Curtis A V,Rodman J,Band S R.A comparative study of germination characteristics in a local flora.Journal of Ecology,1981,69(3):1017-1059.
[8] 姬慧娟,尹林克,严成.刚毛柽柳(Tamarixhispida)种子萌发特性的研究.生物技术,2008,18(6):35-38. Ji H J,Yin L K,Yan C.Studies on germination characteristics ofTamarixhispidaseeds.Biotechnology,2008,18(6):35-38.(in Chinese)
[9] 王磊,严成,魏岩,尹林克.温度、盐分和储藏时间对多花柽柳种子萌发的影响.干旱区研究,2008,25(6):797-801. Wang L,Yan C,Wei Y,Yin L K.Study on the effects of temperature,salinity and storing time on the germination ofTamarixhohenackeriseed.Arid Zone Research,2008,25(6):797-801.(in Chinese)
[10] 魏岩,谭敦炎,尹林克.中国柽柳科植物叶解剖特征与分类关系的探讨.西北植物学报,1999,19(1):113-118. Wei Y,Tan D Y,Yin L K.The discussinos on the anatomical structure of leaf and its taxonomic relationship of Tamaricaceae in China.Acta Botanica Boreali-Occidentalia Sinica,1999,19(1):113-118.(in Chinese)
[11] 张元明,潘伯荣,尹林克.中国干旱区柽柳科植物种子形态特征及其系统学意义.植物资源与环境,1998,7(2):22-27. Zhang Y M,Pan B R,Yin L K.Seed morphology of Tamaricaceae in China arid areas and its systematic evolution.Journal of Plant Resources and Environment,1998,7(2):22-27.(in Chinese)
[12] Yan C,Wei Y,Yang M L.Comparative germination ofTamarixramosissimaspring and summer seeds.EXCLI Journal,2011,10:198-204.
[13] 严成,魏岩,王磊.密花柽柳春夏两季种子的萌发行为.干旱区研究,2010,27(5):750-754. Yan C,Wei Y,Wang L.Study on germination ofTamarixarceuthoidesseeds in spring and summer.Arid Zone Research,2010,27(5):750-754.(in Chinese)
[14] Ungar I A.Ecophysiology of Vascular Halophytes.Boca Raton:CRC Press,1991.
[15] Ungar I A.Population ecology of halophyte seeds.The Botanical Review,1987,53:301-344.
[16] Khan M A,Ungar I A.Influence of salinity and temperature on the germination ofHaloxylonrecurvumBunge ex Boiss.Annals of Botany,1996,78:547-551.
[17] 程龙,李志军,韩占江,石新建.盐节木种子萌发对温度、光照和盐旱胁迫的响应.草业科学,2015,32(6):961-966. Cheng L,Li Z J,Han Z J,Shi X J.Response ofHalocnermumstrobilaceumto light,temperature,salt and drought stresses during seed germination.Pratacultural Science,2015,32(6):961-966.(in Chinese)
[18] 武祎,田雨,张红香,杨健,吴志红.盐、碱胁迫与温度对黄花苜蓿种子发芽的影响.草业科学,2015,32(11):1847-1853. Wu Y,Tian Y,Zhang H X,Yang J,Wu Z H.Effects of salinity,alkalinity,temperature and their interactions on seed germination ofMedicagofalcata.Pratacultural Science,2015,32(11):1847-1853.(in Chinese)
[19] Khan M A.Comparative influence of salinity and temperature on the germination of subtropical perennial halophytes.In:Hamdy A,Lieth H,Todorovis M,Moschenko M.(eds).Halophyte Uses in Different Climates Ⅰ.Ecological and Physiological Studies.Leiden:Backhuys Publishers,1999:1177-1188.
[20] Wei Y,Dong M,Huang Z Y,Dong M.Factors influencing seed germination ofSalsolaaffinis(Chenopodiaceae),a dominant annual halophyte inhabiting the deserts of Xinjiang,China.Flora,2008,203:134-140.
[21] Huang Z Y,Zhang X S,Zheng G H,Gutterman Y.Influence of light,temperature,salinity and storage on germination ofHaloxylonammodendron.Journal of Arid Environments,2003,55:453-464.
[22] Zhang H X,Zhang G M,Lyu X T,Zhou D W,Han X G.Salt tolerance during seed germination and early seedling stages of 12 halophytes.Plant and Soil,2015,388:229-241.
(责任编辑 张瑾)
Influences of thermoperiod and salinity onTamarixgracilisseed germination
Wang Xi-yong1,2, Cai Dan-hong1,3, Wei Yan1
(1.College of Grassland and Environmental Science, Xinjiang Agricultural University, Urumqi 830052, China;2.Xinjiang Ecology and Geography Research Institute, Chinese Academy of Sciences, Urumqi 830011, China;3.Songmen Middle School of Wenling in Zhejiang, Wenling 317500, China)
In order to reveal the seed germination characteristics ofTamarixgracilisin the Turpan Desert Botanical Garden, Xinjiang, seeds of the species were germinated in incubators with 4 photoperiods and 10 NaCl salinities. Experiments were conducted to determine the effects of thermoperiod and NaCl on seed germination. The results showed that 1) seeds can germinate quickly and reach a high germination percentage in a range of temperatures, from 5 ℃/15 ℃ to 25 ℃/35 ℃ in dark/light = 12 h/12 h. The higher the temperature, the faster the germination. 2) The higher the NaCl salinity, the lower the percentage of germination. Germination was not affected by NaCl at concentrations below 0.4 mol·L-1, while germination at higher concentrations (0.6, 0.8, 1.0, and 1.2 mol·L-1) were significantly lower than that of the control. 3) After transferring to distilled water, un-germinated seeds incubated in NaCl showed a low germination recovery rate. Low germination recovery (2.1%~34.5%) was noted at concentrations of 0.6~1.2 mol·L-1. No germination recovery was noted when NaCl concentration was more than 1.2 mol·L-1. This shows that 1.2 mol·L-1NaCl is the largest concentration of salt tolerance ofT.gracilis.
environment; germination characteristics; saline; salt stress;Tamarixgracilis; thermoperiod; Xinjiang
Wei Yan E-mail:weiyan1966@163.com
10.11829/j.issn.1001-0629.2016-0541
2016-10-24 接受日期:2017-04-26
国家自然科学基金(31360091)
王喜勇(1978-),男,新疆新源人,在读博士生,主要从事草地植物资源研究。E-mail:wxy701@tom.com
魏岩(1966-),河南镇平人,教授,博士,主要从事荒漠植物生活史特征及生态适应研究。E-mail:weiyan1966@163.com
Q945.34;S793.5
A
1001-0629(2017)08-1571-05
王喜勇,蔡丹红,魏岩.异花柽柳种子萌发对温周期和盐分的响应.草业科学,2017,34(8):1571-1575.
Wang X Y,Cai D H,Wei Y.Influences of thermoperiod and salinity onTamarixgracilisseed germination.Pratacultural Science,2017,34(8):1571-1575.
猜你喜欢
今日农业(2021年21期)2021-11-26
河北林业科技(2020年3期)2020-03-23
小学生学习指导(低年级)(2018年9期)2018-09-26
读者·校园版(2018年1期)2017-12-16
花卉(2017年7期)2017-11-15
花卉(2017年7期)2017-07-20
中国民族医药杂志(2016年8期)2016-05-09
浙江大学学报(农业与生命科学版)(2015年4期)2015-12-15
化工进展(2015年6期)2015-11-13
少年科学(2015年2期)2015-08-12
